- Катаболизм аминокислот

Содержание
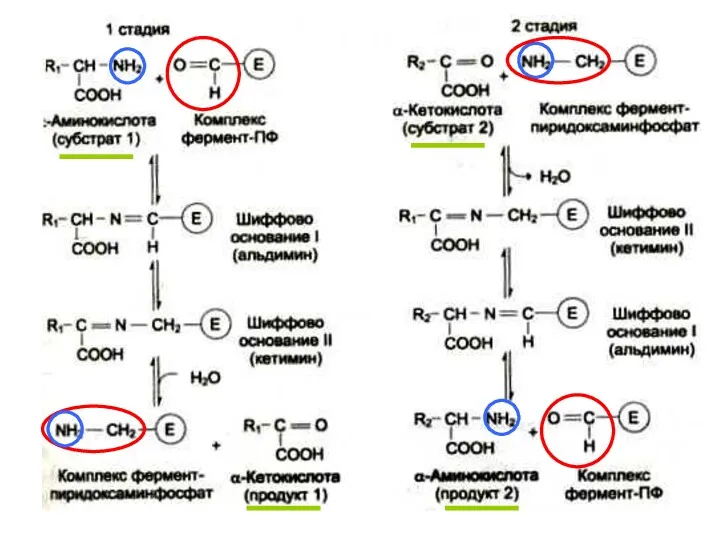
- 2. 1. ТРАНСАМИНИРОВАНИЕ АМИНОКИСЛОТ Реакция переноса α-аминогруппы с аминокилоты на α-кетокислоту. Ре- зультат: образование новой аминокислоты и
- 3. Трансаминирование катализируют аминотрансферазы; кофер-мент – пиридоксальфосфат (предшественник – витамин В6, пири-доксин). Пиридоксальфосфат ковалентно связан с остатком
- 4. Первая стадия реакции трансаминирования
- 6. Наиболее активно трансаминирование происходит с аминокис- лотами, содержание которых в клетке сравнительно высоко. α-аминокислоты: - глутамат
- 7. Физиологическое значение трансаминирования: Обратимость реакций трансаминирование позволяет им участвовать и в катаболизме, и в анаболизме. Трансаминирование
- 8. 2. ДЕЗАМИНИРОВАНИЕ АМИНОКИСЛОТ Реакция дезаминирования: R-NH2 ?? R=O + NH3 амино-кислота кето-кислота Образующаяся кетокислота (безазотистый остаток)
- 9. 2.1. Окислительное дезаминирование. Наиболее интенсивно в клетках человека происходит окисли-тельное дезаминирование глутаминовой кислоты; обратимую реакцию катализирует
- 10. Кофермент глутаматдегидрогеназы – НАД+ (NAD+) – никотин-амидадениндинуклеотид (фосфат) Для синтеза НАД+ и НАДФ+ необходим водорастворимый вита-мин
- 11. Механизм транспорта водорода с участием НАД+/НАДН (показана только часть молекулы) В восстановленной форме НАДН, один атом
- 12. Роль окислительного дезаминирования аминокислот: образова- ние α-кетоглютарата. Поступает в ЦТК, окисляется, участвуя в синтезе АТФ. В
- 13. Оксидазы аминокислот (обнаружены в почках и печени) Оксидаза L-аминокислот. Кофактор реакции – ФМН (флавин-мононуклеотид). Вклад этой
- 14. Для синтеза кофермента ФАД+ (ФМН+) необходим водораствори- мый витамин В2 (рибофлавин), витамин роста.
- 15. Механизм транспорта водорода с участием ФАД+/ФАДН2 (показана только часть молекулы)
- 17. Скачать презентацию

1. ТРАНСАМИНИРОВАНИЕ АМИНОКИСЛОТ
Реакция переноса α-аминогруппы с аминокилоты на α-кетокислоту. Ре-
зультат:
1. ТРАНСАМИНИРОВАНИЕ АМИНОКИСЛОТ
Реакция переноса α-аминогруппы с аминокилоты на α-кетокислоту. Ре-
зультат:

Трансаминирование катализируют аминотрансферазы; кофер-мент – пиридоксальфосфат (предшественник – витамин В6,
Трансаминирование катализируют аминотрансферазы; кофер-мент – пиридоксальфосфат (предшественник – витамин В6,
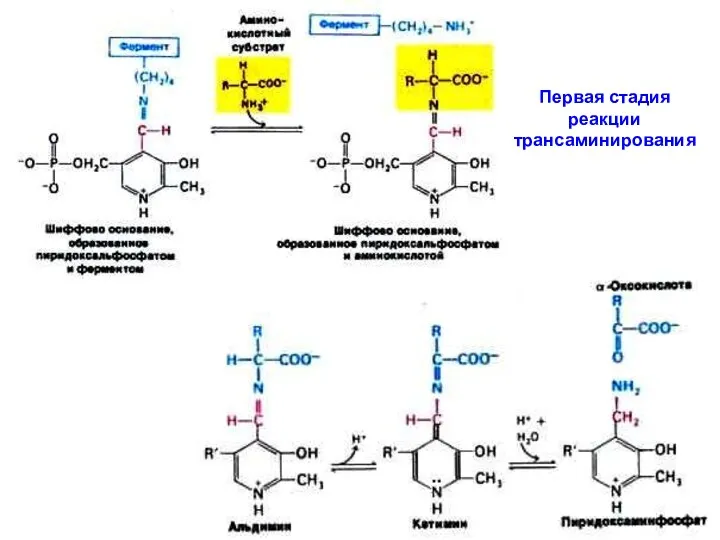
Первая стадия
реакции
трансаминирования
Первая стадия
реакции
трансаминирования

Наиболее активно трансаминирование происходит с аминокис-
лотами, содержание которых в клетке
Наиболее активно трансаминирование происходит с аминокис-
лотами, содержание которых в клетке

Физиологическое значение трансаминирования:
Обратимость реакций трансаминирование позволяет
им участвовать и
Физиологическое значение трансаминирования:
Обратимость реакций трансаминирование позволяет
им участвовать и

2. ДЕЗАМИНИРОВАНИЕ АМИНОКИСЛОТ
Реакция дезаминирования:
R-NH2 ?? R=O + NH3
2. ДЕЗАМИНИРОВАНИЕ АМИНОКИСЛОТ
Реакция дезаминирования:
R-NH2 ?? R=O + NH3
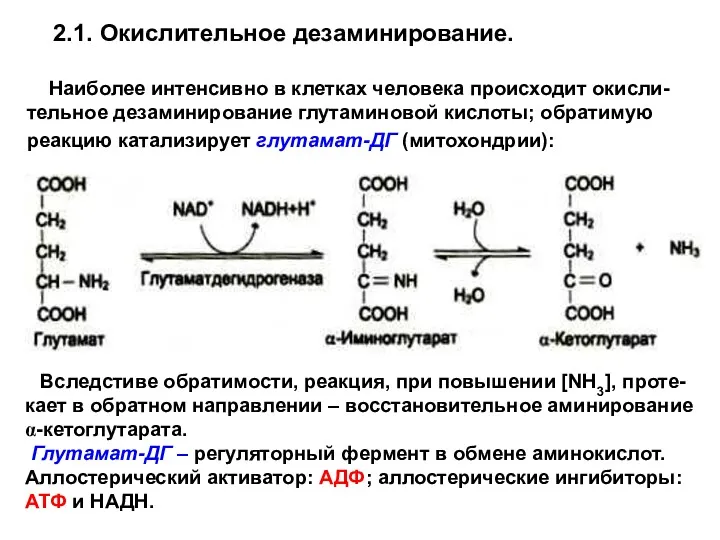
2.1. Окислительное дезаминирование.
Наиболее интенсивно в клетках человека происходит окисли-тельное
2.1. Окислительное дезаминирование.
Наиболее интенсивно в клетках человека происходит окисли-тельное
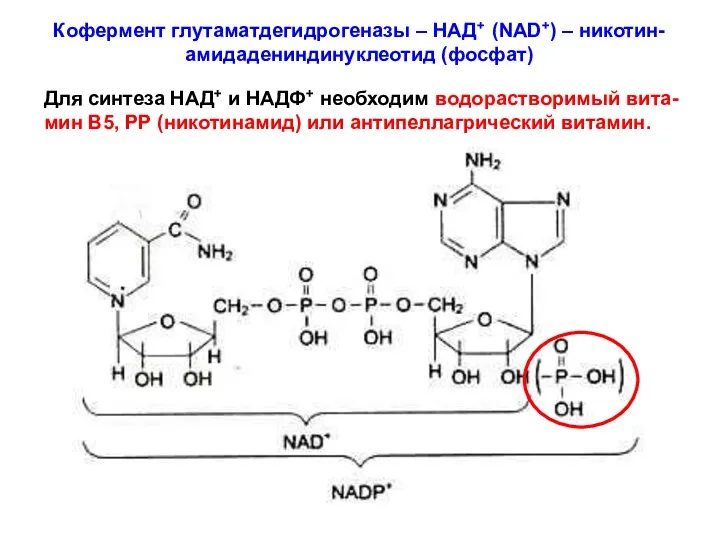
Кофермент глутаматдегидрогеназы – НАД+ (NAD+) – никотин-амидадениндинуклеотид (фосфат)
Для синтеза НАД+ и
Кофермент глутаматдегидрогеназы – НАД+ (NAD+) – никотин-амидадениндинуклеотид (фосфат)
Для синтеза НАД+ и
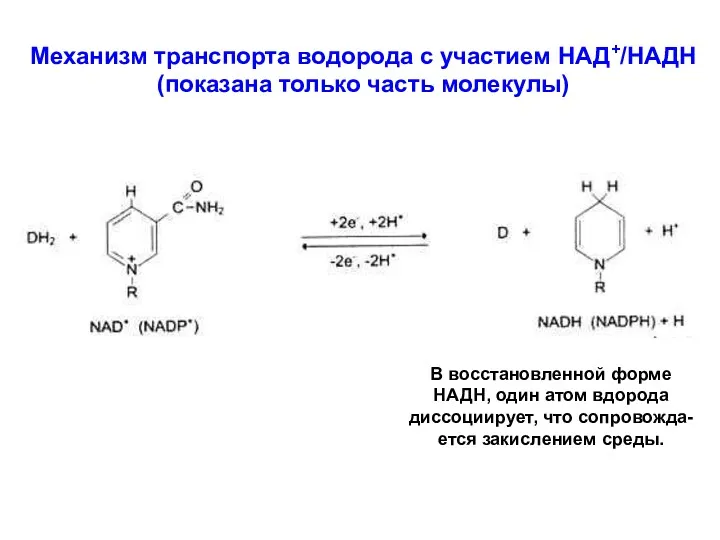
Механизм транспорта водорода с участием НАД+/НАДН
(показана только часть молекулы)
В восстановленной форме
НАДН,
Механизм транспорта водорода с участием НАД+/НАДН
(показана только часть молекулы)
В восстановленной форме
НАДН,

Роль окислительного дезаминирования аминокислот: образова-
ние α-кетоглютарата. Поступает в ЦТК, окисляется,
Роль окислительного дезаминирования аминокислот: образова-
ние α-кетоглютарата. Поступает в ЦТК, окисляется,

Оксидазы аминокислот (обнаружены в почках и печени)
Оксидаза L-аминокислот. Кофактор реакции
Оксидазы аминокислот (обнаружены в почках и печени)
Оксидаза L-аминокислот. Кофактор реакции
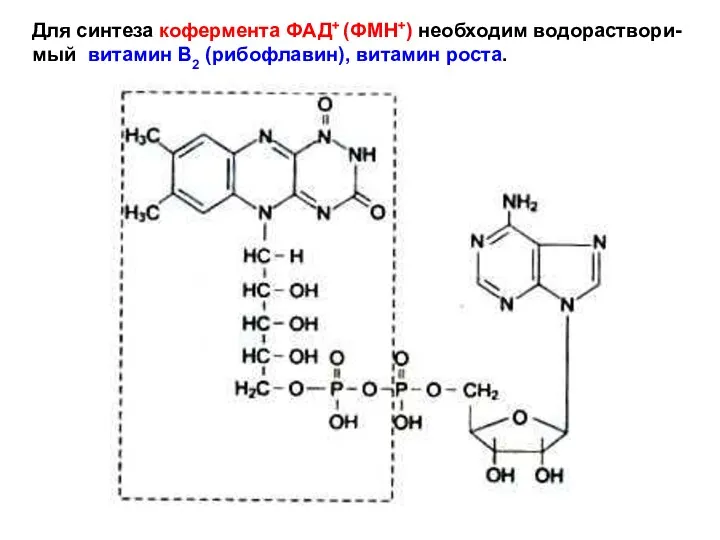
Для синтеза кофермента ФАД+ (ФМН+) необходим водораствори-
мый витамин В2 (рибофлавин), витамин
Для синтеза кофермента ФАД+ (ФМН+) необходим водораствори-
мый витамин В2 (рибофлавин), витамин
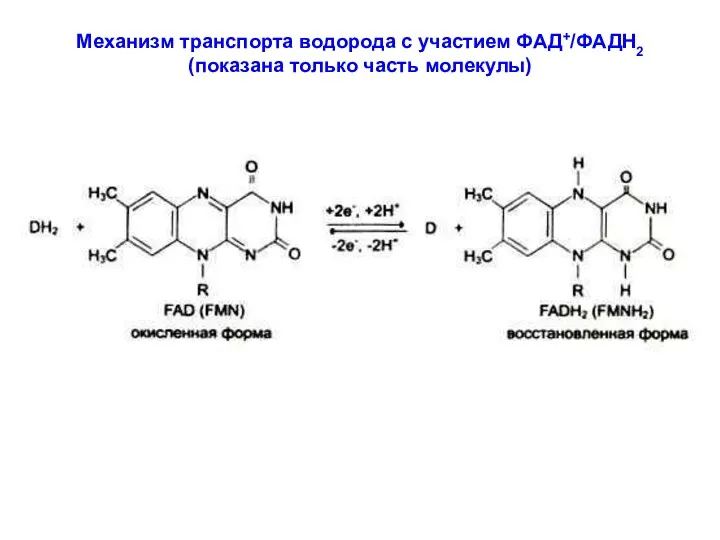
Механизм транспорта водорода с участием ФАД+/ФАДН2
(показана только часть молекулы)
Механизм транспорта водорода с участием ФАД+/ФАДН2
(показана только часть молекулы)
 Значение фотосинтеза и биологического круговорота веществ в развитии жизни
Значение фотосинтеза и биологического круговорота веществ в развитии жизни В гости к весне. (2 класс)
В гости к весне. (2 класс) Цитоскелет. Включения
Цитоскелет. Включения Презентация к уроку биологии 6 кл.
Презентация к уроку биологии 6 кл. Неорганические вещества клетки
Неорганические вещества клетки Рыбы Онежского озера
Рыбы Онежского озера зрительный_анализатор
зрительный_анализатор Видообразование. Результат макроэволюции
Видообразование. Результат макроэволюции Лекция. Витамины
Лекция. Витамины Класс птицы
Класс птицы Терморегуляция
Терморегуляция Развитие и устойчивость экосистем
Развитие и устойчивость экосистем Викторина Животные
Викторина Животные Строение амёбоидной клетки, на примере Amoeba proteus и Arcella sp
Строение амёбоидной клетки, на примере Amoeba proteus и Arcella sp Маленькая система большого значения или вся правда об эндокринной системе
Маленькая система большого значения или вся правда об эндокринной системе Патогенность микроорганизмов. Вирулентность. Факторы адгезии, мимикрия, ферменты, токсины
Патогенность микроорганизмов. Вирулентность. Факторы адгезии, мимикрия, ферменты, токсины Ритмы здоровья. Витаминно-минеральный комплекс
Ритмы здоровья. Витаминно-минеральный комплекс Бионика. Виды бионики
Бионика. Виды бионики Губки. Происхождение многоклеточных
Губки. Происхождение многоклеточных Отряд Парнокопытные
Отряд Парнокопытные Отряд Опоссумы (Didelphimorphia)
Отряд Опоссумы (Didelphimorphia) Водоросли – низшие растения
Водоросли – низшие растения Плоские черви
Плоские черви Немного о лецитине
Немного о лецитине Строение и функции белков – 2
Строение и функции белков – 2 Насекомые. Отличительные признаки насекомых (1 класса)
Насекомые. Отличительные признаки насекомых (1 класса) Пищеварительная система
Пищеварительная система Тайны лесного царства. Игра
Тайны лесного царства. Игра