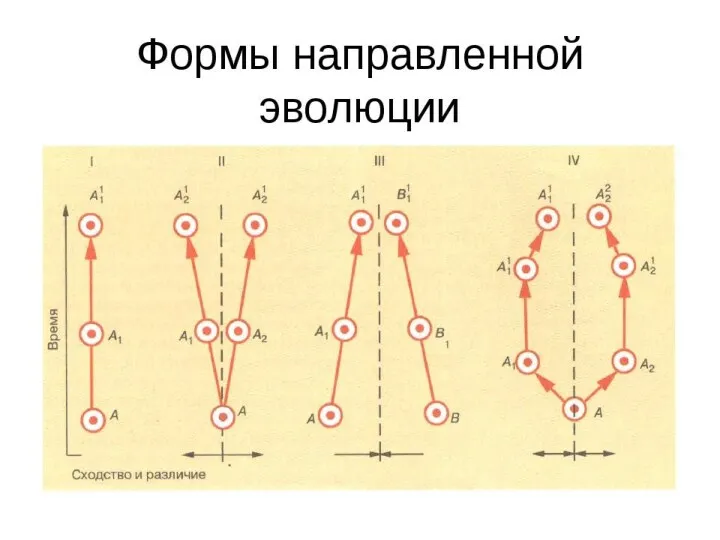
- Направления и формы эволюции групп. Лекция 2

Содержание
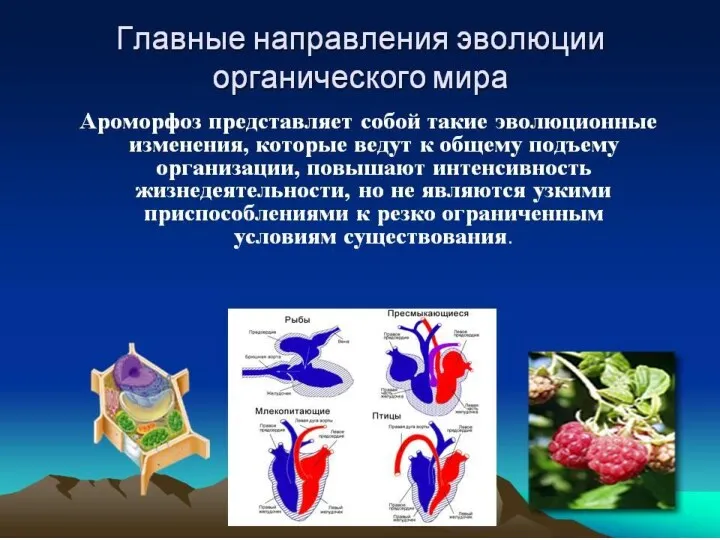
- 4. 1) Морфофизиологический прогресс - за счет ароморфозов - приспособлений, которые существенно повышают уровень организации живых организмов.
- 6. паразит саккулина у крабов, имеющий вид мешка, набитого половыми продуктами, и обладающий пищеварительной системой, пронизывающей тело
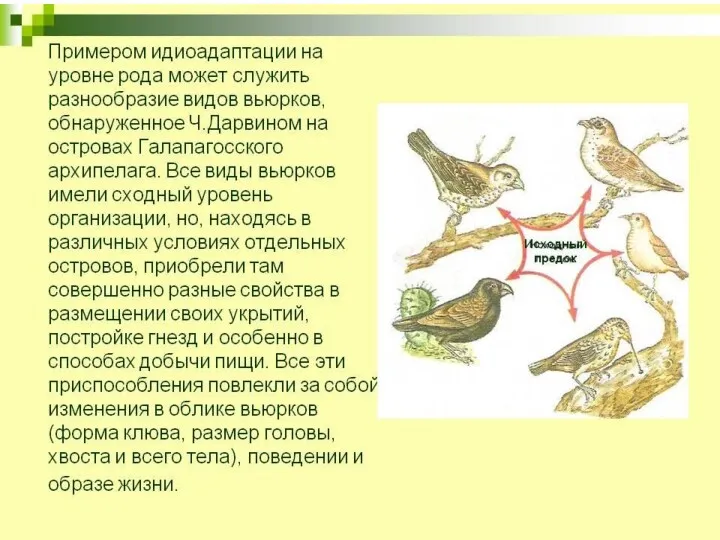
- 8. За счет приспособлений к среде без принципиальной перестройки биологической организации. Пример — разнообразие форм у насекомоядных
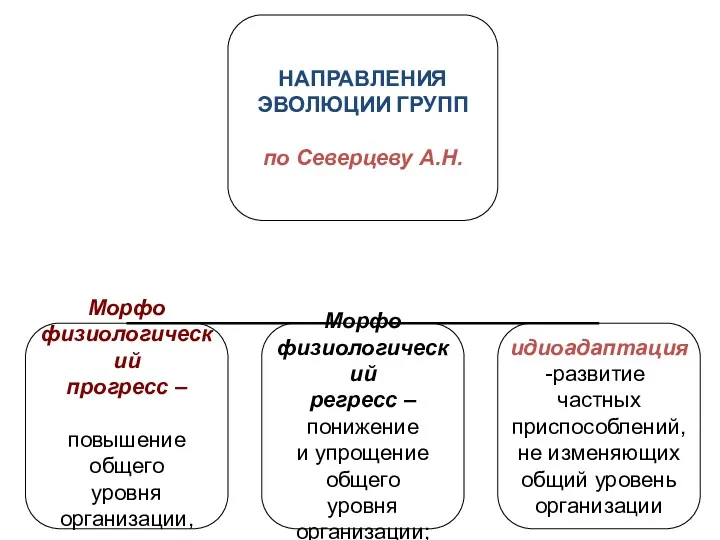
- 11. А.Л. Тахтаджян А.А. Парамонов Б. Ренш НАПРАВЛЕНИЯ ЭВОЛЮЦИИ ГРУПП
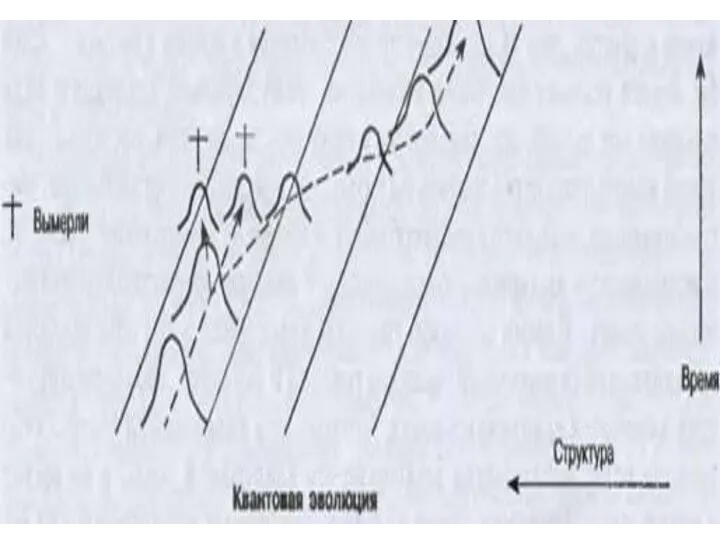
- 14. Современная классификация направлений эволюции групп I. АРОГЕНЕЗ ( греч. -поднимать) - это развитие группы с расширением
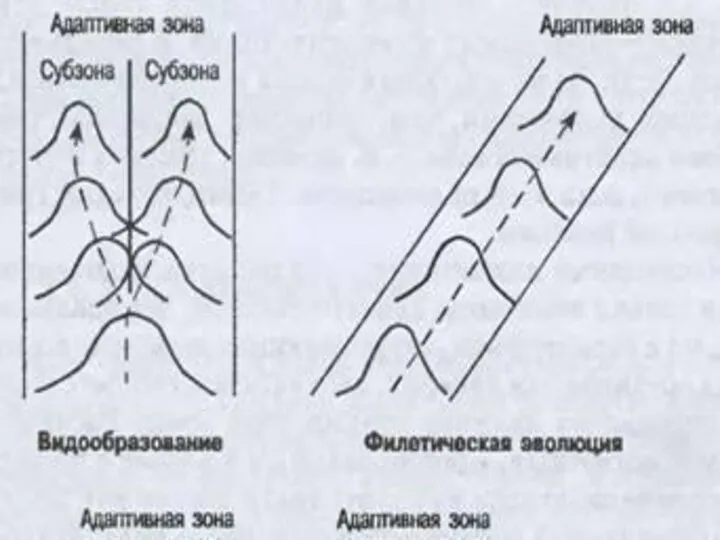
- 16. Адаптивная зона – не только среда обитания, среда эволюции любого надвидового таксона. Так, границы адаптивной зоны
- 17. Понятие адаптивной зоны подразумевает также и ограничения, затрудняющие изменения направления эволюции. Границы адаптивной зоны определяются организацией
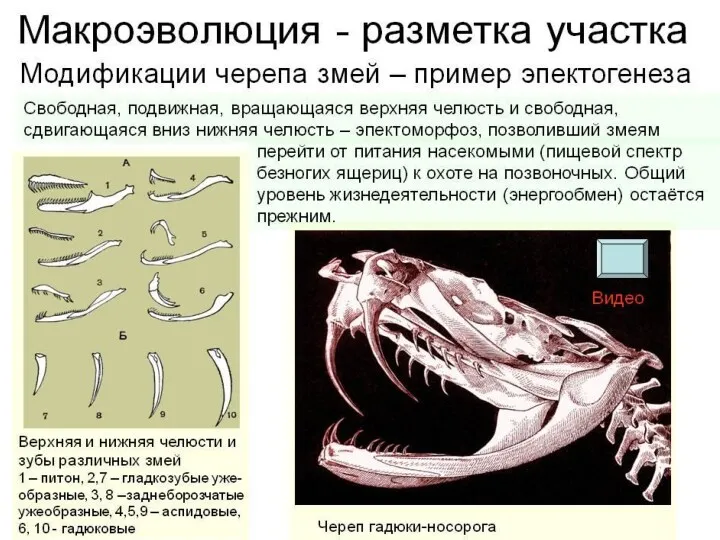
- 20. Эпекто морфозы
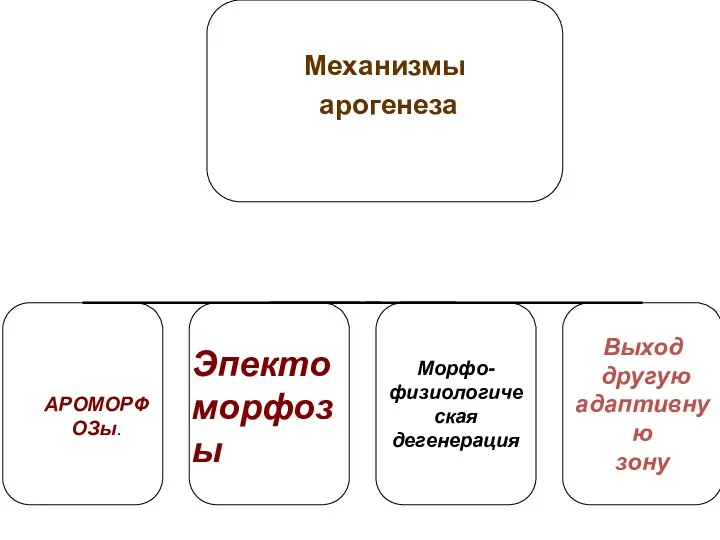
- 21. I. Арогенез Механизмы: АРОМОРФОЗЫ (морфофизиологический прогресс).Усложнение строения, приобретение крупных приспособлений - крыло птиц, 4-х камерное сердца,
- 22. 2. Эпектоморфоз За счет эпектоморфозов - эволюционных преобразований, увеличивающих адаптивные возможности группы, через каскадные изменения. Оказывают
- 23. Эпектоморфоз: развитие кутикулы в покровах нематод.
- 24. Следствия: кутикула (гибкая, прочная и химически устойчивая) ? освоение разнообразных местообитаний (толща органических осадков на дне
- 26. 3. Морфофизиологическая дегенерация Например, паразитизм.
- 27. 4. Выход в другую адаптивную зону через изменение образа жизни и поведения. В дальнейшем группа может:
- 28. II. АЛЛОГЕНЕЗ - развитие группы внутри одной адаптивной зоны с возникновением форм, различающихся адаптациями одного масштаба.
- 29. Ликвидация специализации при аллогенезе: - за счет неотении, - расширения адаптивной зоны - выхода в другую
- 31. Формы эволюции ФИЛОГЕНЕТИЧЕСКИХ ГРУПП
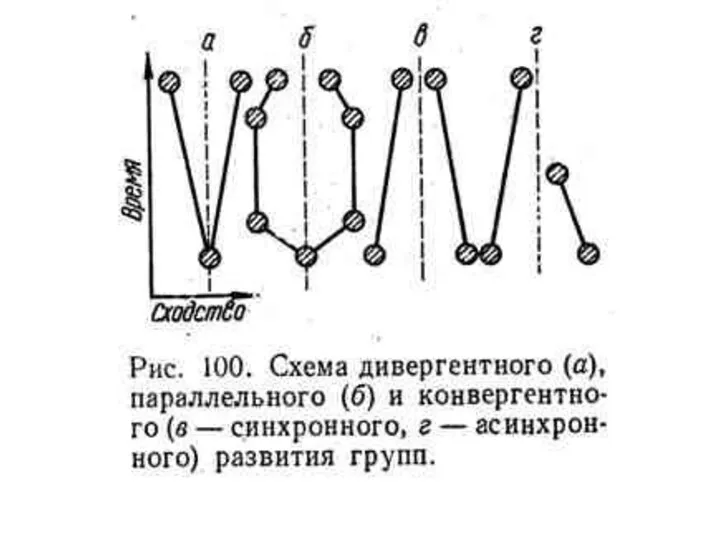
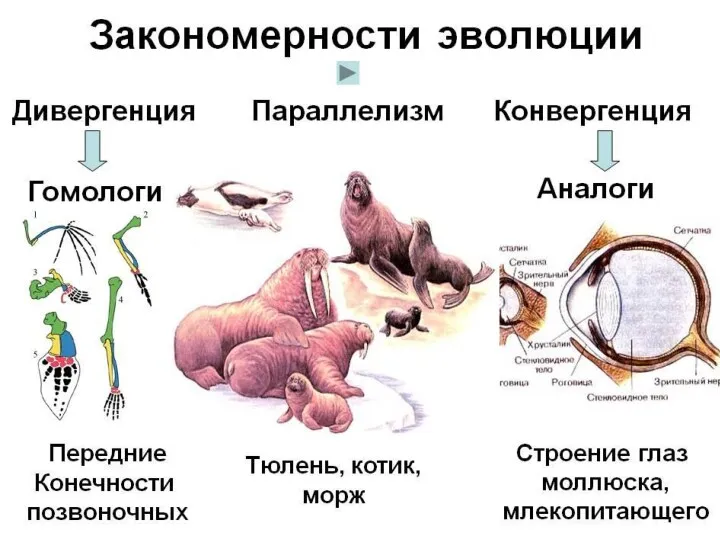
- 32. СОПОСТАВЛЕНИЕ ХОДА ЭВОЛЮЦИОННОГО ПРОЦЕССА РАЗНЫХ СИСТЕМАТИЧЕСКИХ ГРУПП выявило общие закономерности сначала на морфофизиологическом уровне, а сейчас
- 33. 1. Филетическая эволюция это изменение во времени как вида, так и близкородственных групп видов в одном
- 34. Данные молекулярной генетики подтвердили монофилию голо-и покрытосеменных растений от одной общей группы (возможно от семенных папоротников.)
- 35. Затем отделились линии к магнолиидам, однодольным, высшим двудольным. Установление монофилии позволяет уточнить датировку происхождения п/с (не
- 36. Монофилия – происхождение таксона от одного предкового таксона. Симпсон предложил считать монофилетическими таксоны, которые порождены таксоном,
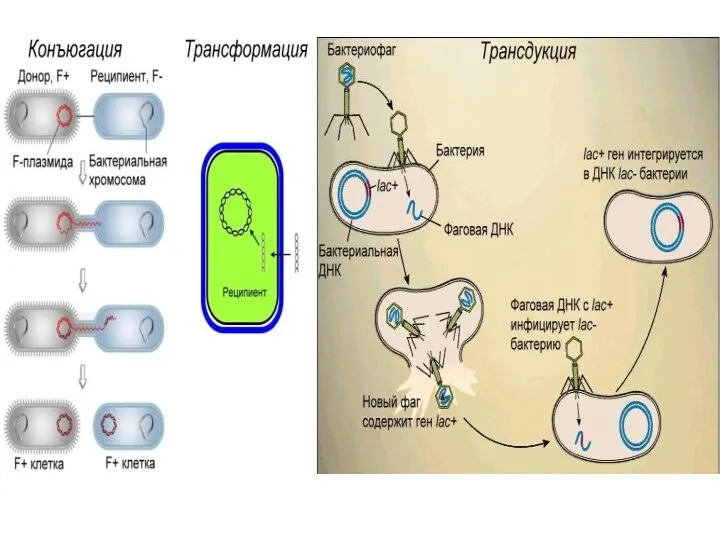
- 37. 2. ПОЛИФИЛИТИЧЕСКОЕ происхождение Полифелия - происхождение нового таксона от разных групп - где генетические барьеры могут
- 38. Гибридизация
- 39. Симбиогенез
- 40. Гетерокариоз — сосуществование генетически различных гаплоидных ядер в цитоплазме одной клетки как результат нерасхождения дочерних ядер
- 42. Монофилия = жёлтый, Полифилия = красный, Парафилия = голубой При парафилетическом происхождении группы включают лишь часть
- 43. При филетической эволюции за счет обмена генами между отдельными группировками возникает (аналогичное внутрипопуляционным связям) - СЕТЧАТОЕ
- 44. Схема дивергенции основных групп (доменов и царств) организмов с учетом симбиотического происхождения митохондрий и пластид Схема
- 45. 2. ДИВЕРГЕНЦИЯ Это расхождение ветвей от единого ствола предков в результате изменения направлений отбора + при
- 47. Дивергенция начинается на микроэволюционном уровне, вследствие полиморфизма вида при разнообразии условий среды под действием расчленяющего отбора.
- 49. Дивергенция любого надвидового уровня ---результат действия группового отбора и изоляции. Процесс необратимый. Возникший вид не может
- 50. Сходные элементы конечностей позвоночных прослеживаются спустя сотни миллионов лет после начала дивергенции. Гомологичные органы, то есть
- 51. Гомологичные органы Конечности млекопитающих: А — стопоходная носухи, В — роющая крота , С—летательная летучей собаки,
- 52. Вторичные (дополнительные) формы развития групп 1. КОНВЕРГЕНЦИЯ - это процесс формирования сходного фенотипического облика групп в
- 54. Крыло стрекозы и летучей мыши имеют общие черты в строении и функционировании, но формируются в ходе
- 55. Примеры конвергенции 1- Череп мадагаскарского лемура, 2- череп грызуна; 3 — лапа хамелеона; 4 — лапа
- 56. Пример конвергенции: акулы, ихтиозавры, киты
- 57. Примеры: прыгуны открытых пространств; зайцеобразные и грызуны; волки, медведи, кроты у сумчатых и плацентарных занимающие сходные
- 58. Конвергентная эволюция животных, которые эволюционируют в сходных условиях среды, занимая сходные экологические ниши. приводит к сходству
- 59. Пример: Ниша муравьедов на разных континентах была заполнена разными неродственными видами млекопитающих. Имеют длинный липкий язык
- 60. А – восточно американский луговой трупиал Б- африканский желтогорлый длинношпорцевый конек В - австралийский вомбат Г-
- 61. Конвергенция Положения головы: у лягушки (1), крокодила (2) бегемота (3).
- 62. 2. Параллелизм -формирование сходного облика, первоначально разошедшихся генетически близких групп. То есть независимое схождение в идентичных
- 64. Синхронный параллелизм. Пример1. В конце силура из костных рыб выделились двоякодышащие и кистеперые. Обитание в мелких
- 65. Примеры возникновения похожих наборов "жизненных форм" на разобщенных континентах. В Южную Америку еще до наступления полной
- 67. Асинхронный параллелизм. Саблезубость у представителей кошачьих возникала 4 раза в двух независимых стволах в разное время.
- 68. Вымершие саблезубые формы хищных млекопитающих Слева – южно-американские саблезубые тигры, Справа – австралийский сумчатый волк.
- 69. Костные панцири появлялись в эволюции дважды у бесчелюстных и примитивных хрящевых рыб
- 70. Примеры параллельной эволюции среди пресноводных цихлид в Америке и Африке , которые потеряли связь около 100
- 71. Схожие по экстерьеру тонкогубые (вверху) и толстогубые (внизу) цихлиды в озере Манагуа (Центральная Америка) в озере
- 77. Традиционные и современные подходы филогенетики .
- 78. 1. Традиционные – филогенетическое древо
- 79. 2. Кладистика Символом стала кладограмма – стилизованное филогенетическое дерево. На нем, в отличие от филограммы, показывающей
- 80. Стилизованные представления филогенетических схем: а – филограмма, б – кладограмма. Сплошными линиями показаны парафилетические группы, штриховыми
- 81. Методология - отказ от учета отношений предок-потомок при обсуждении родства и монофилии. Т.к. конкретный предковый вид
- 83. Основу большинства алгоритмов новой филогенетики составляет принцип синапоморфии, согласно которому монофилию группы можно выявить только через
- 84. Последнее составляет один из важнейших критериев – значимость признаков: их «вес» обратно пропорционален вероятности параллелизмов и
- 85. Категории сходства, связанные с концепцией синапоморфии. Заглавными буквами обозначены апоморфии, строчными – плезиоморфии; цветом выделены апоморфии,
- 86. Основу большинства алгоритмов новой филогенетики составляет принцип синапоморфии, согласно которому монофилию группы можно выявить только через
- 87. Основные методы, связаны с разной трактовкой начальных условий реконструкций экономии (парсимонии) предполагают полный отказ от априорного
- 88. Активная «нумеризация» новой филогенетики все более переносит акцент с теории и методологии реконструкций на их «технологию».
- 89. Особый раздел нумерической филетики составляет компьютерное моделирование филогенетических процессов. Позволяет исследовать некоторые свойства исторического развития организмов
- 90. 4. Концепция молекулярных часов в филогении Была разработана для перехода от сходства по молекулярным структурам -?
- 91. Молекулярные часы . Метод датирования филогенетических событий (расхождений видов или других таксонов), основанный на гипотезе, согласно
- 97. Филогенетические связи между 8 группами позвоночных животных (верхняя схема) и различия по числу аминокислотных замен (Каа)
- 98. 5. Аддитивное дерево, - по различиям в транспортных РНК.
- 99. Филогенетическое древо, построенное на основе анализа последовательностей генов рРНК, показывает общее происхождение организмов всех трёх доменов:
- 100. Бескорневое филогенетическое древо, построенное на сравнительном анализе генов, кодирующих синтез Fe- и Mn- супероксиддисмутазы (SOD). Красным
- 102. Количественная оценка различий по молекулярным структурам (генетическая дистанция) дает собственное время дивергенции, напрямую несопоставимое для разных
- 103. Сравнение времени дивергенции основных групп органического мира по данным ископаемой летописи и молекулярных часов. Масштаб шкалы
- 104. Задача калибровки «молекулярных часов» не решается в рамках самой молекулярной биологии: результат зависит от палеонтологических данных.
- 106. 6. Генофилетика (филогеномика) – раздел новой филогенетики, в котором заключения о кладистической истории выводятся на основании
- 107. В генофилетике при реконструкции родственных отношений особое значение придается изменениям в первичной структуре макромолекул, не связанным
- 108. Реконструкция предковой последовательности +-YERPE | | +BACAN | +----4 | | | +BACCR | +-----3 +--5
- 109. Реконструкция филогении по биологическим последовательностям
- 110. Современные представления о филогении эвкариот, основанные на синтезе данных ультраструктурных исследований и сравнительной геномики
- 111. Филогенетические отношения ранних фораминифер по данным сравнительной геномики
- 113. Скачать презентацию
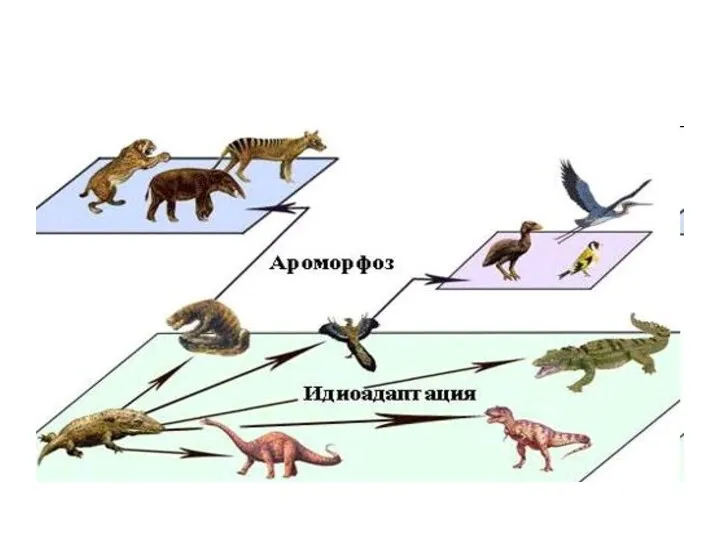

1) Морфофизиологический прогресс - за счет ароморфозов
- приспособлений, которые
1) Морфофизиологический прогресс - за счет ароморфозов
- приспособлений, которые

паразит саккулина у крабов, имеющий вид мешка, набитого половыми продуктами, и
паразит саккулина у крабов, имеющий вид мешка, набитого половыми продуктами, и


За счет приспособлений к среде без принципиальной перестройки биологической организации.
Пример
За счет приспособлений к среде без принципиальной перестройки биологической организации.
Пример

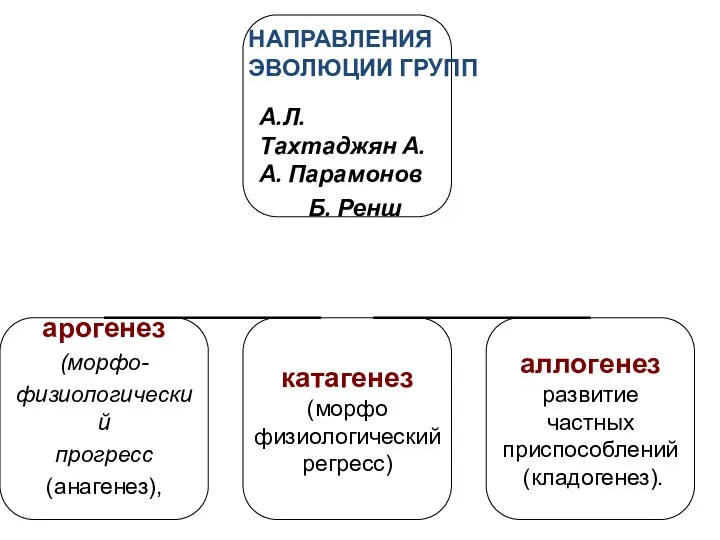
А.Л. Тахтаджян А.А. Парамонов
Б. Ренш
НАПРАВЛЕНИЯ
ЭВОЛЮЦИИ ГРУПП
А.Л. Тахтаджян А.А. Парамонов
Б. Ренш
НАПРАВЛЕНИЯ
ЭВОЛЮЦИИ ГРУПП
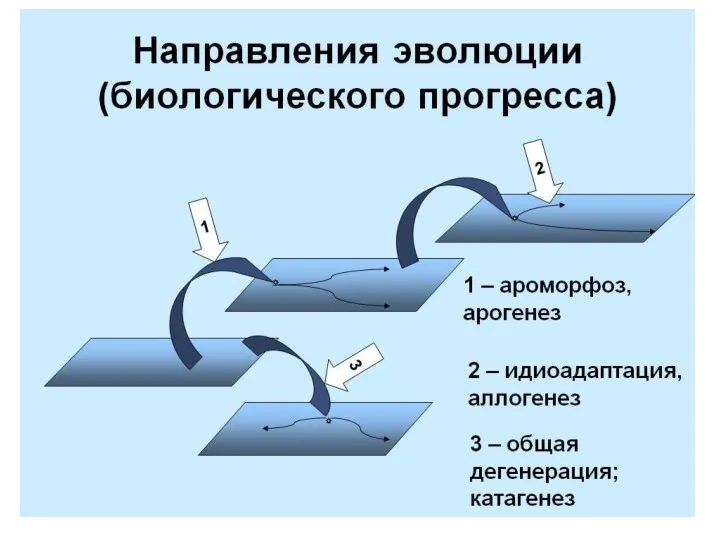
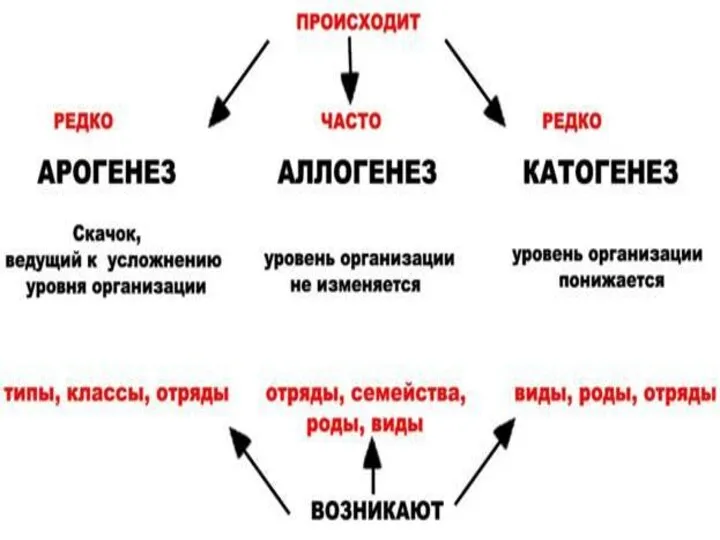

Современная классификация направлений эволюции групп
I. АРОГЕНЕЗ
( греч. -поднимать)
-
Современная классификация направлений эволюции групп
I. АРОГЕНЕЗ
( греч. -поднимать)
-
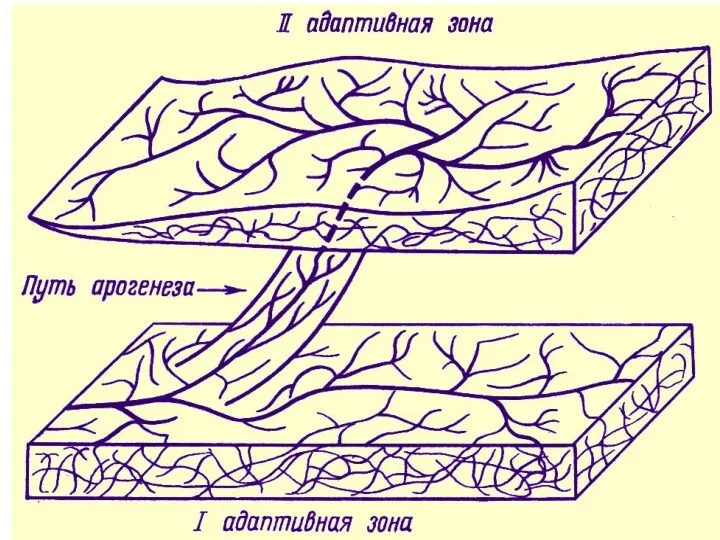

Адаптивная зона – не только среда обитания,
среда эволюции любого надвидового
Адаптивная зона – не только среда обитания,
среда эволюции любого надвидового

Понятие адаптивной зоны подразумевает также и ограничения, затрудняющие изменения направления эволюции.
Понятие адаптивной зоны подразумевает также и ограничения, затрудняющие изменения направления эволюции.
Эпекто
морфозы
Эпекто
морфозы

I. Арогенез
Механизмы:
АРОМОРФОЗЫ (морфофизиологический прогресс).Усложнение строения, приобретение крупных приспособлений -
крыло
I. Арогенез
Механизмы:
АРОМОРФОЗЫ (морфофизиологический прогресс).Усложнение строения, приобретение крупных приспособлений -
крыло

2. Эпектоморфоз
За счет эпектоморфозов - эволюционных преобразований, увеличивающих адаптивные возможности группы,
2. Эпектоморфоз
За счет эпектоморфозов - эволюционных преобразований, увеличивающих адаптивные возможности группы,
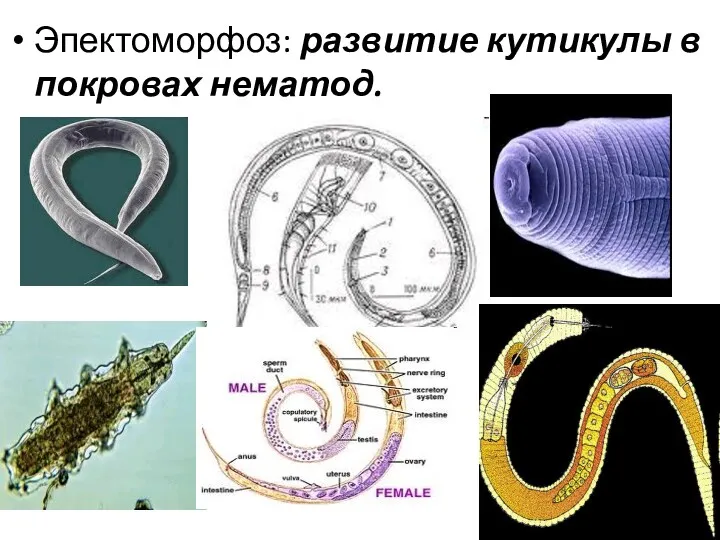
Эпектоморфоз: развитие кутикулы в покровах нематод.
Эпектоморфоз: развитие кутикулы в покровах нематод.

Следствия:
кутикула (гибкая, прочная и химически устойчивая) ? освоение разнообразных местообитаний (толща
Следствия:
кутикула (гибкая, прочная и химически устойчивая) ? освоение разнообразных местообитаний (толща
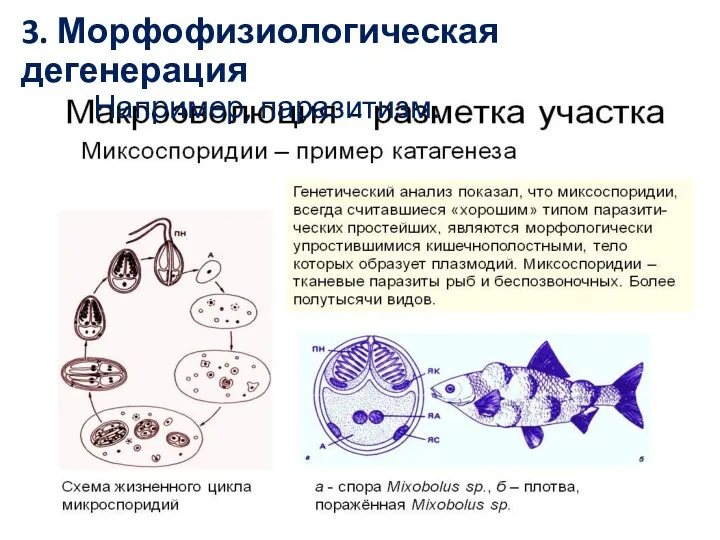
3. Морфофизиологическая дегенерация
Например, паразитизм.
3. Морфофизиологическая дегенерация
Например, паразитизм.

4. Выход в другую адаптивную зону через изменение образа жизни и
4. Выход в другую адаптивную зону через изменение образа жизни и

II. АЛЛОГЕНЕЗ - развитие группы внутри одной адаптивной зоны с
II. АЛЛОГЕНЕЗ - развитие группы внутри одной адаптивной зоны с

Ликвидация специализации при аллогенезе:
- за счет неотении,
- расширения
Ликвидация специализации при аллогенезе:
- за счет неотении,
- расширения

Формы эволюции
ФИЛОГЕНЕТИЧЕСКИХ ГРУПП
Формы эволюции
ФИЛОГЕНЕТИЧЕСКИХ ГРУПП

СОПОСТАВЛЕНИЕ ХОДА ЭВОЛЮЦИОННОГО ПРОЦЕССА РАЗНЫХ СИСТЕМАТИЧЕСКИХ ГРУПП
выявило общие закономерности
СОПОСТАВЛЕНИЕ ХОДА ЭВОЛЮЦИОННОГО ПРОЦЕССА РАЗНЫХ СИСТЕМАТИЧЕСКИХ ГРУПП
выявило общие закономерности

1. Филетическая эволюция
это изменение во времени как вида, так и близкородственных
1. Филетическая эволюция
это изменение во времени как вида, так и близкородственных

Данные молекулярной генетики подтвердили монофилию голо-и покрытосеменных растений от одной общей
Данные молекулярной генетики подтвердили монофилию голо-и покрытосеменных растений от одной общей

Затем отделились линии к магнолиидам, однодольным, высшим двудольным.
Установление монофилии позволяет
Затем отделились линии к магнолиидам, однодольным, высшим двудольным.
Установление монофилии позволяет

Монофилия – происхождение таксона от одного предкового таксона.
Симпсон предложил считать
Монофилия – происхождение таксона от одного предкового таксона.
Симпсон предложил считать

2. ПОЛИФИЛИТИЧЕСКОЕ происхождение
Полифелия - происхождение нового таксона от разных групп -
2. ПОЛИФИЛИТИЧЕСКОЕ происхождение
Полифелия - происхождение нового таксона от разных групп -
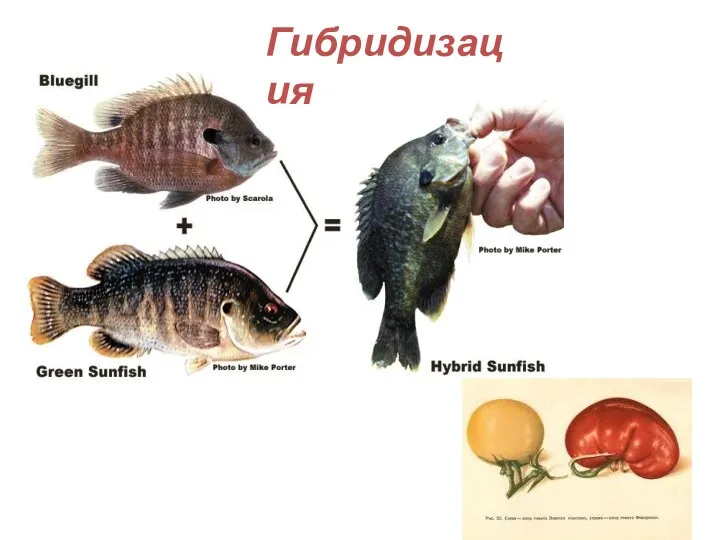
Гибридизация
Гибридизация
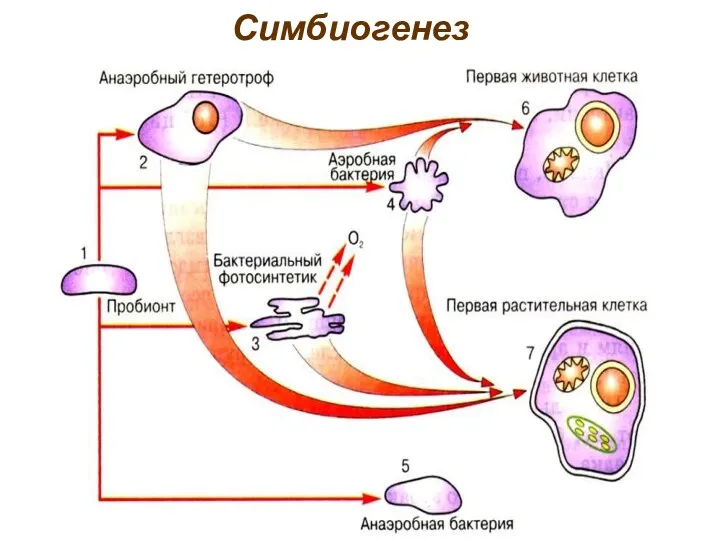
Симбиогенез
Симбиогенез
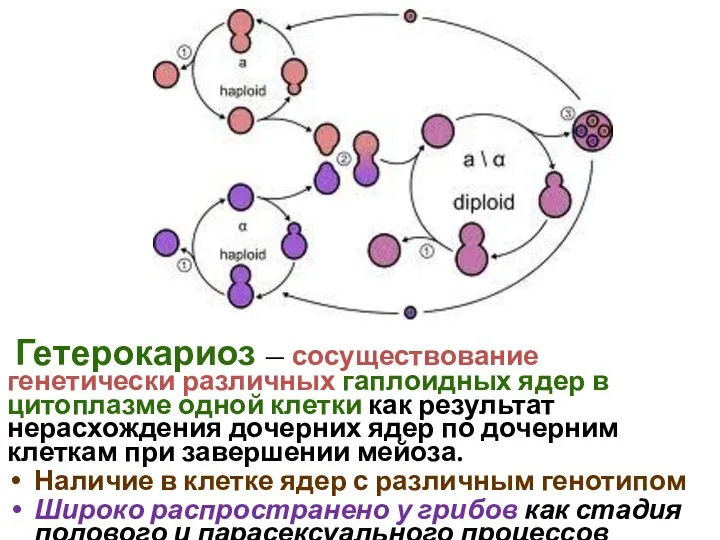
Гетерокариоз — сосуществование генетически различных гаплоидных ядер в цитоплазме одной
Гетерокариоз — сосуществование генетически различных гаплоидных ядер в цитоплазме одной
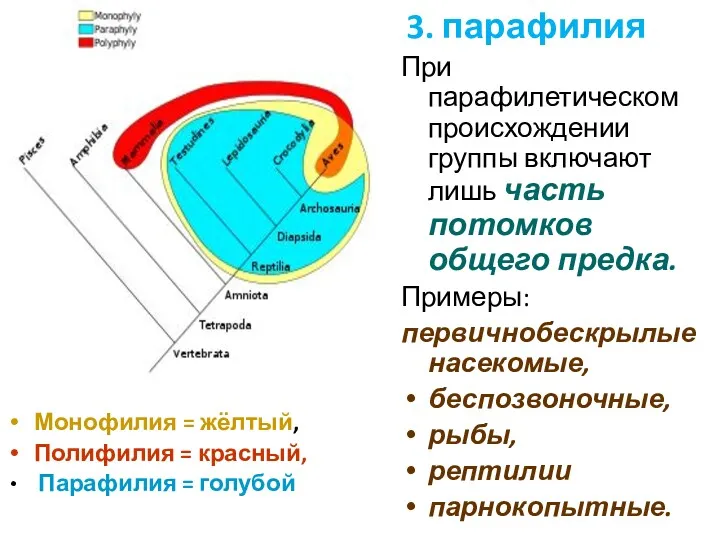
Монофилия = жёлтый,
Полифилия = красный,
Парафилия = голубой
При парафилетическом
Монофилия = жёлтый,
Полифилия = красный,
Парафилия = голубой
При парафилетическом

При филетической эволюции за счет обмена генами между отдельными группировками
возникает (аналогичное
При филетической эволюции за счет обмена генами между отдельными группировками
возникает (аналогичное
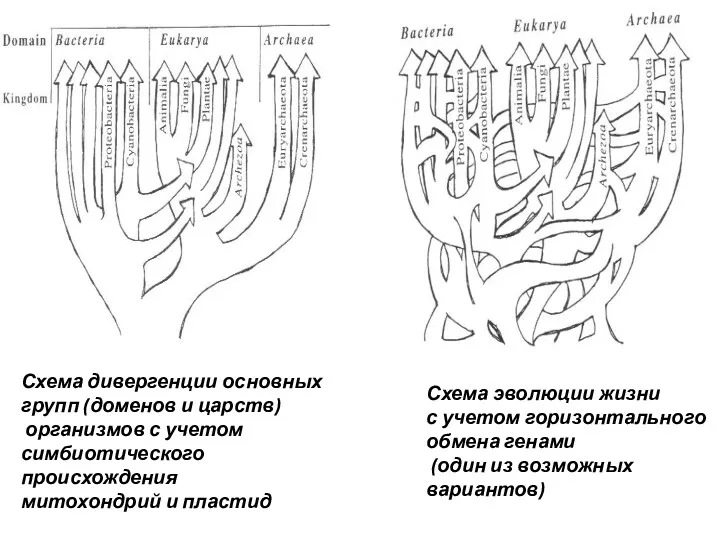
Схема дивергенции основных групп (доменов и царств)
организмов с учетом
Схема дивергенции основных групп (доменов и царств)
организмов с учетом
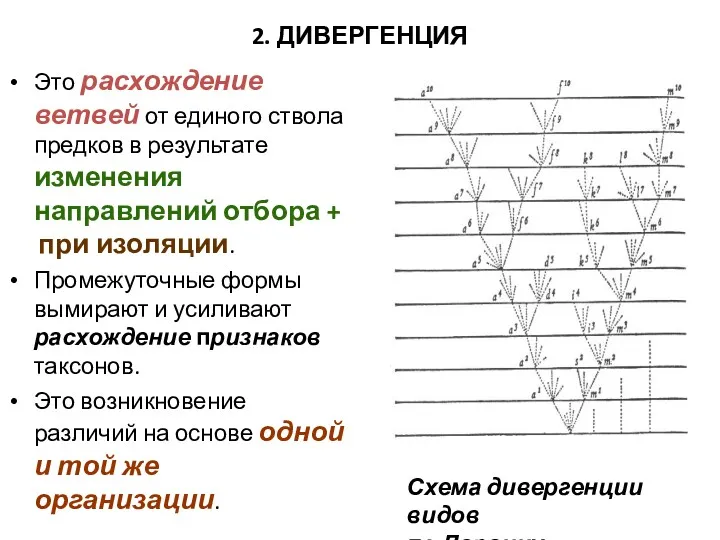
2. ДИВЕРГЕНЦИЯ
Это расхождение ветвей от единого ствола предков в результате изменения
2. ДИВЕРГЕНЦИЯ
Это расхождение ветвей от единого ствола предков в результате изменения
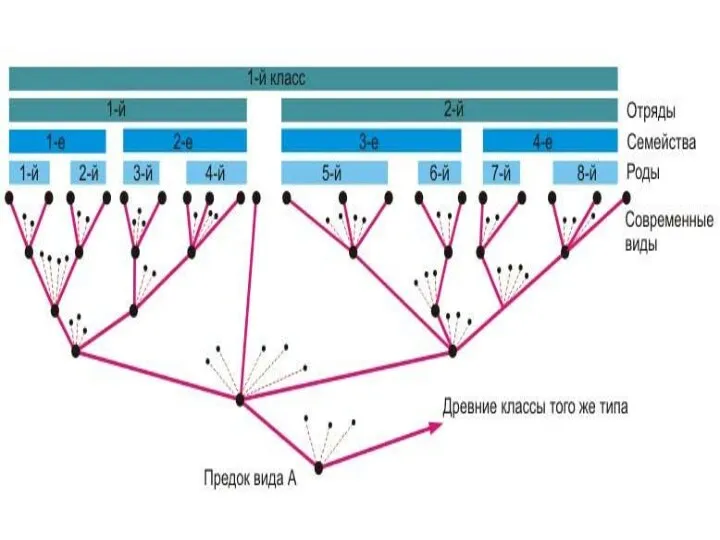

Дивергенция начинается на микроэволюционном уровне, вследствие полиморфизма вида
при разнообразии условий
Дивергенция начинается на микроэволюционном уровне, вследствие полиморфизма вида
при разнообразии условий


Дивергенция любого надвидового уровня ---результат действия группового отбора
и изоляции.
Дивергенция любого надвидового уровня ---результат действия группового отбора
и изоляции.
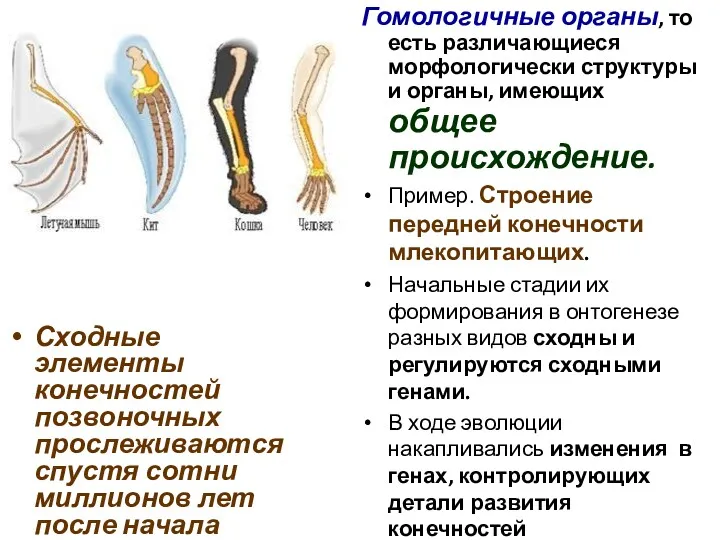
Сходные элементы конечностей позвоночных прослеживаются спустя сотни миллионов лет после начала
Сходные элементы конечностей позвоночных прослеживаются спустя сотни миллионов лет после начала
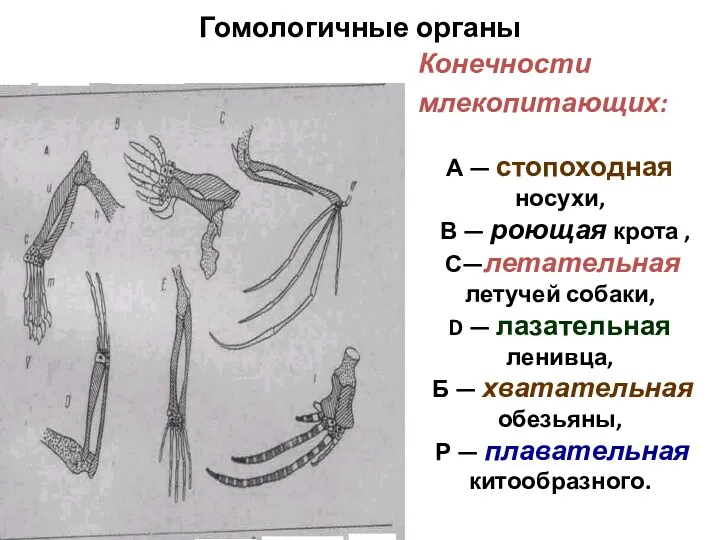
Гомологичные органы
Конечности
млекопитающих:
А — стопоходная носухи,
В — роющая крота ,
С—летательная
Гомологичные органы
Конечности
млекопитающих:
А — стопоходная носухи,
В — роющая крота ,
С—летательная

Вторичные (дополнительные)
формы развития групп
1. КОНВЕРГЕНЦИЯ
- это процесс формирования
Вторичные (дополнительные)
формы развития групп
1. КОНВЕРГЕНЦИЯ
- это процесс формирования


Крыло стрекозы и летучей мыши имеют общие черты в строении и
Крыло стрекозы и летучей мыши имеют общие черты в строении и
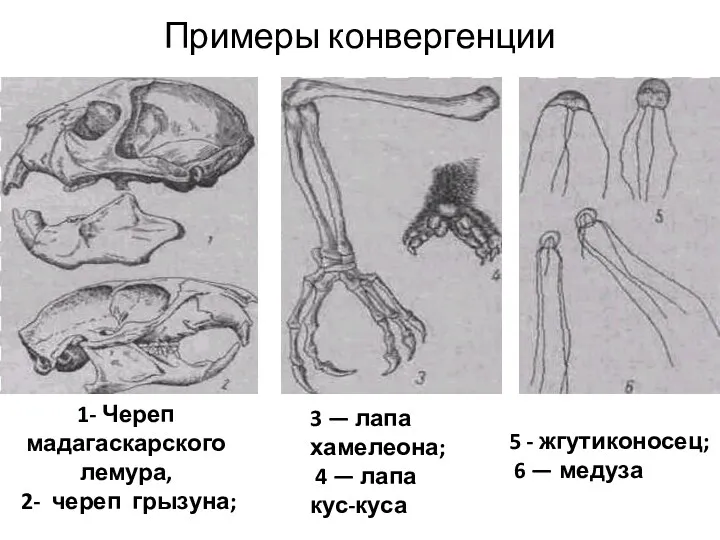
Примеры конвергенции
1- Череп мадагаскарского лемура,
2- череп грызуна;
3 —
Примеры конвергенции
1- Череп мадагаскарского лемура,
2- череп грызуна;
3 —

Пример конвергенции: акулы, ихтиозавры, киты
Пример конвергенции: акулы, ихтиозавры, киты

Примеры:
прыгуны открытых пространств;
зайцеобразные и грызуны;
волки, медведи, кроты у сумчатых
Примеры:
прыгуны открытых пространств;
зайцеобразные и грызуны;
волки, медведи, кроты у сумчатых

Конвергентная эволюция животных, которые эволюционируют в сходных условиях среды, занимая сходные

Пример:
Ниша муравьедов на разных континентах была заполнена разными неродственными видами
Пример:
Ниша муравьедов на разных континентах была заполнена разными неродственными видами
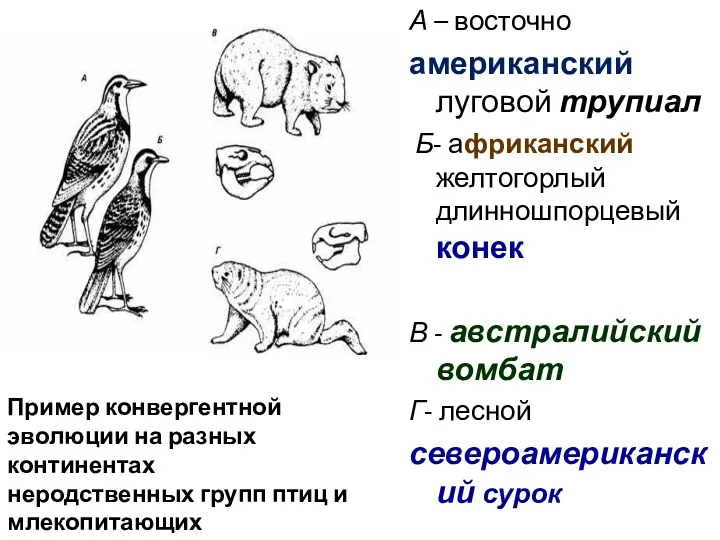
А – восточно
американский луговой трупиал
Б- африканский желтогорлый длинношпорцевый конек
В -
А – восточно
американский луговой трупиал
Б- африканский желтогорлый длинношпорцевый конек
В -
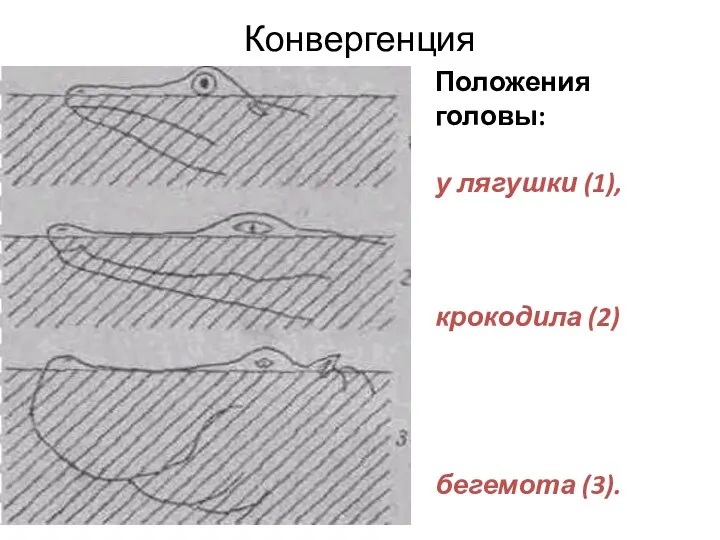
Конвергенция
Положения головы:
у лягушки (1),
крокодила (2)
бегемота (3).
Конвергенция
Положения головы:
у лягушки (1),
крокодила (2)
бегемота (3).

2. Параллелизм
-формирование сходного облика, первоначально разошедшихся генетически близких групп.
То есть
2. Параллелизм
-формирование сходного облика, первоначально разошедшихся генетически близких групп.
То есть
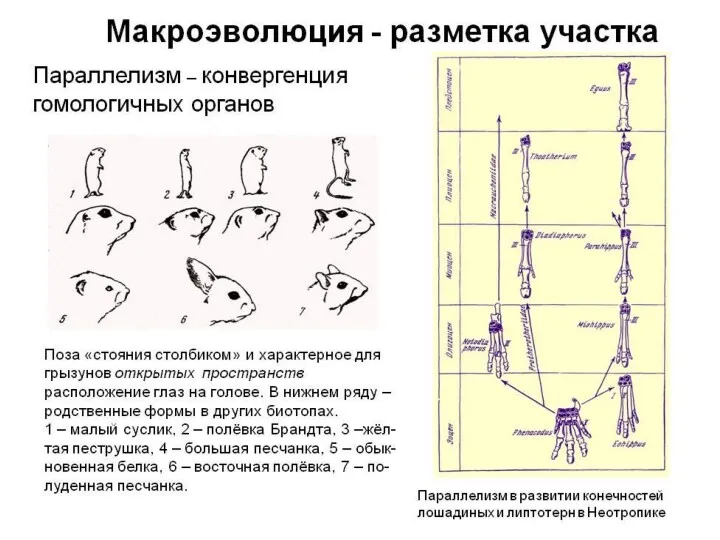

Синхронный параллелизм.
Пример1. В конце силура из костных рыб выделились двоякодышащие
Синхронный параллелизм.
Пример1. В конце силура из костных рыб выделились двоякодышащие

Примеры возникновения похожих наборов "жизненных форм" на разобщенных континентах.
В Южную
Примеры возникновения похожих наборов "жизненных форм" на разобщенных континентах.
В Южную


Асинхронный параллелизм.
Саблезубость у представителей кошачьих
возникала 4 раза в двух независимых стволах
Асинхронный параллелизм.
Саблезубость у представителей кошачьих
возникала 4 раза в двух независимых стволах
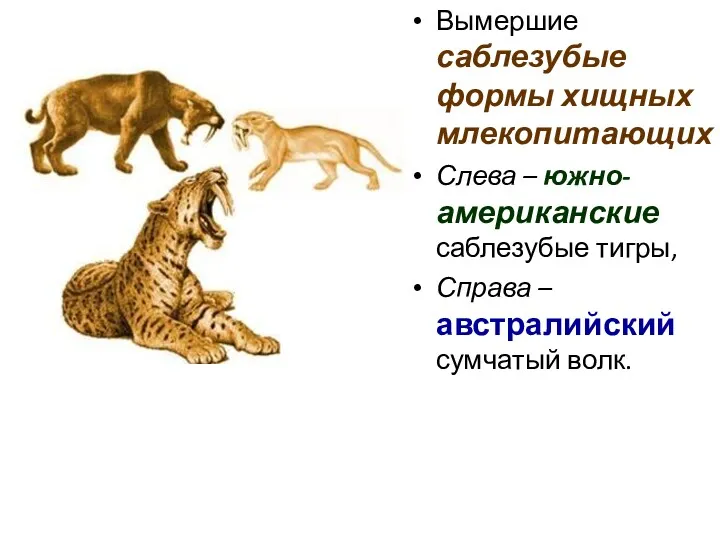
Вымершие саблезубые формы хищных млекопитающих
Слева – южно-американские саблезубые тигры,
Справа
Вымершие саблезубые формы хищных млекопитающих
Слева – южно-американские саблезубые тигры,
Справа

Костные панцири появлялись в эволюции дважды
у бесчелюстных и примитивных хрящевых
Костные панцири появлялись в эволюции дважды
у бесчелюстных и примитивных хрящевых

Примеры параллельной эволюции
среди пресноводных цихлид в Америке и Африке ,
Примеры параллельной эволюции
среди пресноводных цихлид в Америке и Африке ,
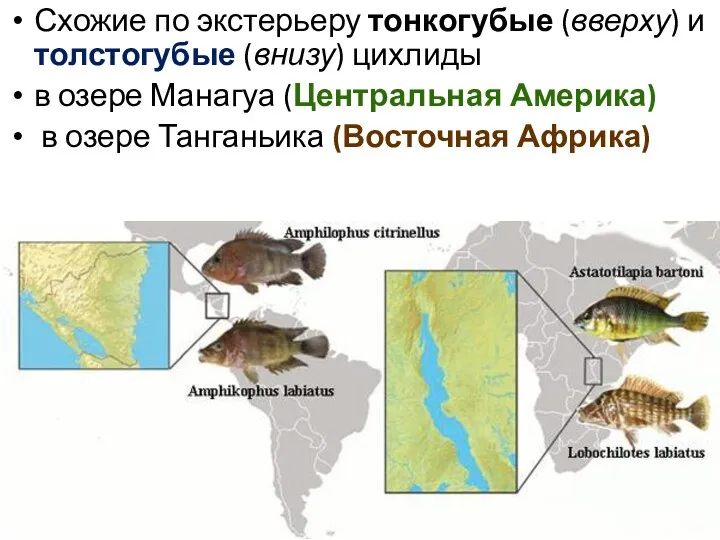
Схожие по экстерьеру тонкогубые (вверху) и толстогубые (внизу) цихлиды
в озере
Схожие по экстерьеру тонкогубые (вверху) и толстогубые (внизу) цихлиды
в озере



Традиционные и современные подходы филогенетики .
Традиционные и современные подходы филогенетики .
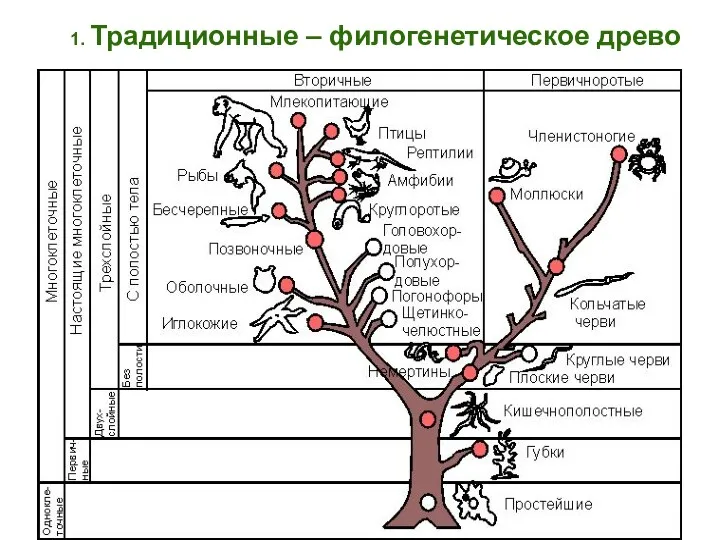
1. Традиционные – филогенетическое древо
1. Традиционные – филогенетическое древо

2. Кладистика
Символом стала кладограмма – стилизованное филогенетическое дерево.
На нем,
2. Кладистика
Символом стала кладограмма – стилизованное филогенетическое дерево.
На нем,
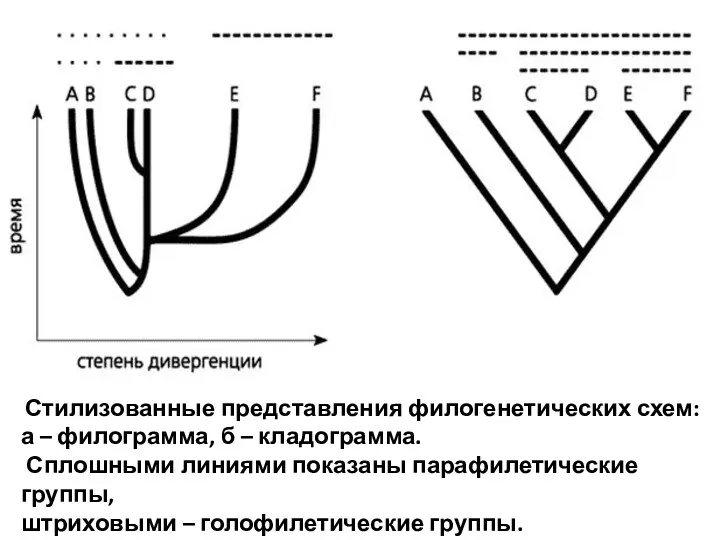
Стилизованные представления филогенетических схем:
а – филограмма, б – кладограмма.
Стилизованные представления филогенетических схем:
а – филограмма, б – кладограмма.

Методология - отказ от учета отношений предок-потомок при обсуждении родства и
Методология - отказ от учета отношений предок-потомок при обсуждении родства и
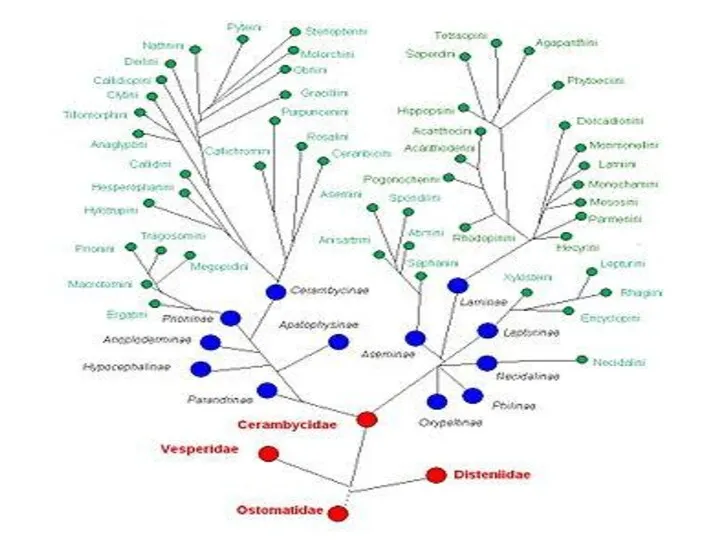

Основу большинства алгоритмов новой филогенетики составляет принцип синапоморфии, согласно которому
Основу большинства алгоритмов новой филогенетики составляет принцип синапоморфии, согласно которому

Последнее составляет один из важнейших критериев – значимость признаков: их «вес»
Последнее составляет один из важнейших критериев – значимость признаков: их «вес»
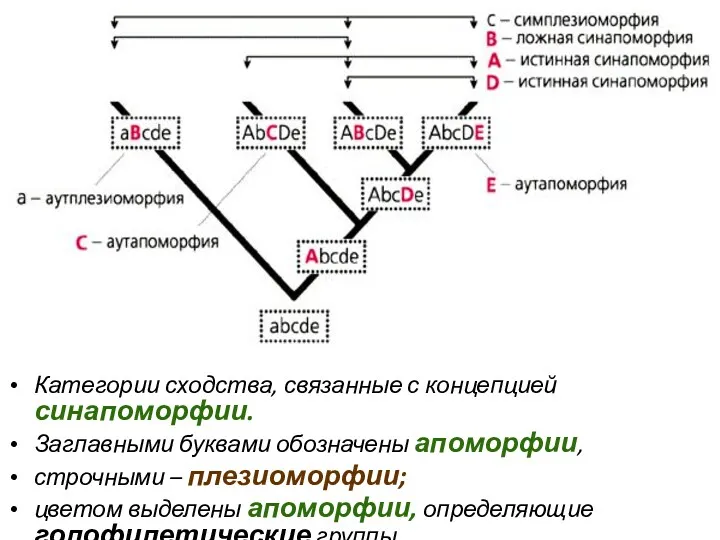
Категории сходства, связанные с концепцией синапоморфии.
Заглавными буквами обозначены апоморфии,
строчными
Категории сходства, связанные с концепцией синапоморфии.
Заглавными буквами обозначены апоморфии,
строчными

Основу большинства алгоритмов новой филогенетики составляет принцип синапоморфии, согласно которому монофилию

Основные методы, связаны с разной трактовкой начальных условий реконструкций
экономии (парсимонии) предполагают
Основные методы, связаны с разной трактовкой начальных условий реконструкций
экономии (парсимонии) предполагают

Активная «нумеризация» новой филогенетики все более переносит акцент с теории и
Активная «нумеризация» новой филогенетики все более переносит акцент с теории и

Особый раздел нумерической филетики составляет
компьютерное моделирование филогенетических процессов.
Позволяет исследовать
Особый раздел нумерической филетики составляет
компьютерное моделирование филогенетических процессов.
Позволяет исследовать

4. Концепция молекулярных часов в филогении
Была разработана для перехода
от
4. Концепция молекулярных часов в филогении
Была разработана для перехода
от

Молекулярные часы .
Метод датирования филогенетических событий (расхождений видов или других
Молекулярные часы .
Метод датирования филогенетических событий (расхождений видов или других

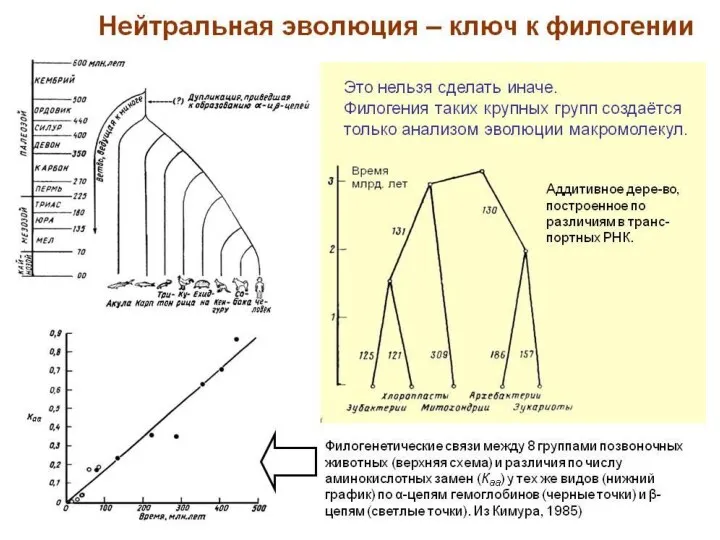


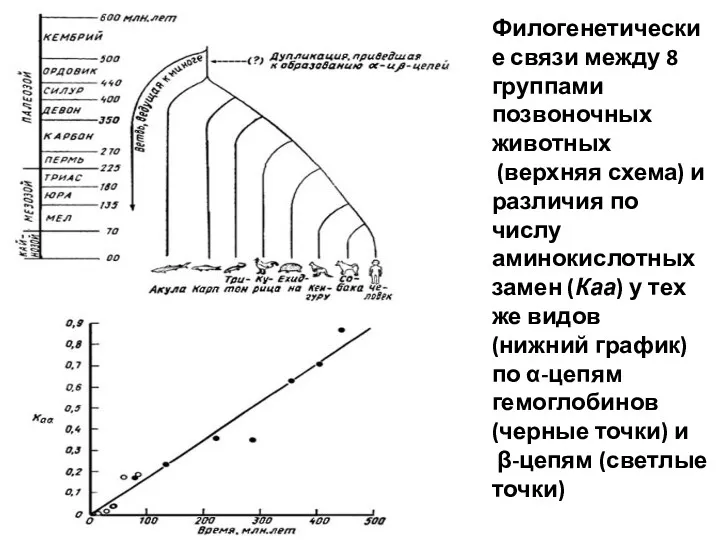
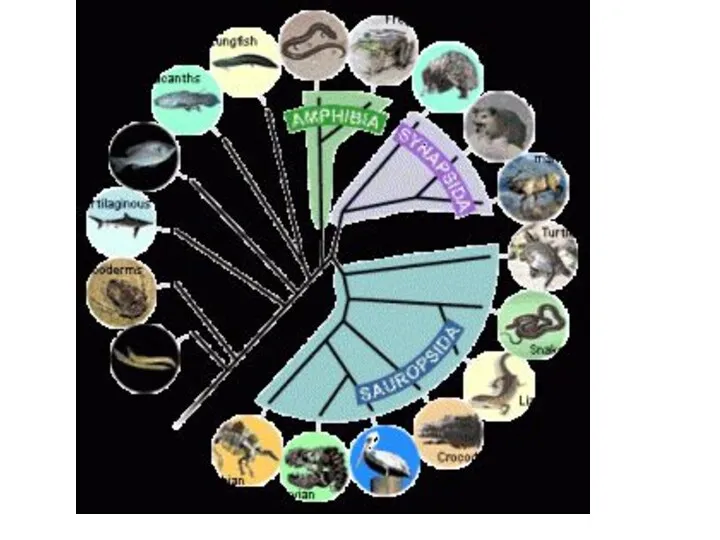
Филогенетические связи между 8 группами позвоночных животных
(верхняя схема) и различия
Филогенетические связи между 8 группами позвоночных животных
(верхняя схема) и различия
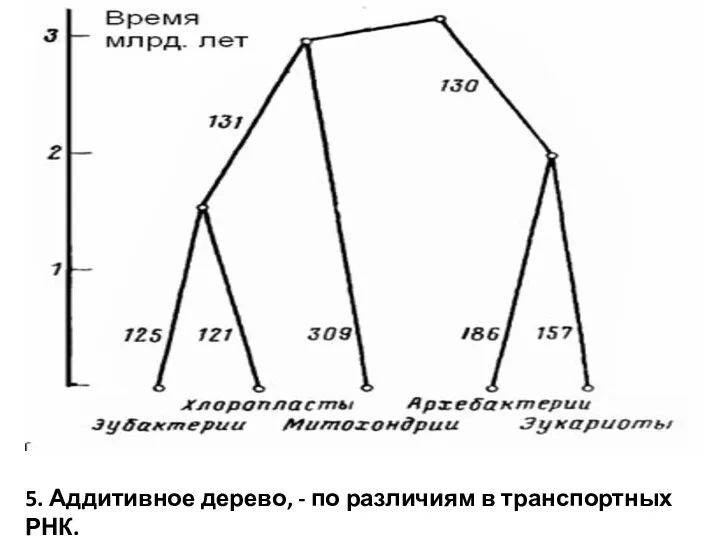
5. Аддитивное дерево, - по различиям в транспортных РНК.
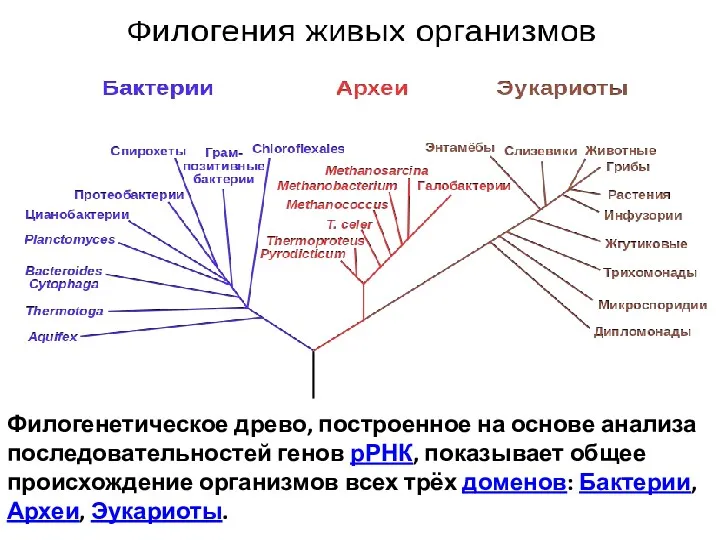
Филогенетическое древо, построенное на основе анализа последовательностей генов рРНК, показывает общее
Филогенетическое древо, построенное на основе анализа последовательностей генов рРНК, показывает общее
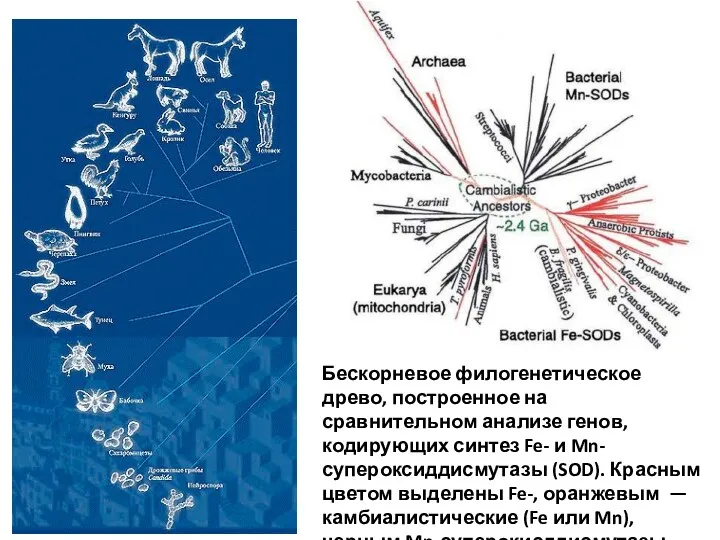
Бескорневое филогенетическое древо, построенное на сравнительном анализе генов, кодирующих синтез Fe-
Бескорневое филогенетическое древо, построенное на сравнительном анализе генов, кодирующих синтез Fe-

Количественная оценка различий по молекулярным структурам (генетическая дистанция) дает собственное время
Количественная оценка различий по молекулярным структурам (генетическая дистанция) дает собственное время

Сравнение времени дивергенции основных групп органического мира по данным ископаемой летописи
Сравнение времени дивергенции основных групп органического мира по данным ископаемой летописи

Задача калибровки «молекулярных часов» не решается в рамках самой молекулярной биологии:
Задача калибровки «молекулярных часов» не решается в рамках самой молекулярной биологии:

6. Генофилетика (филогеномика)
– раздел новой филогенетики, в котором заключения о
6. Генофилетика (филогеномика)
– раздел новой филогенетики, в котором заключения о

В генофилетике при реконструкции родственных отношений особое значение придается изменениям в
В генофилетике при реконструкции родственных отношений особое значение придается изменениям в
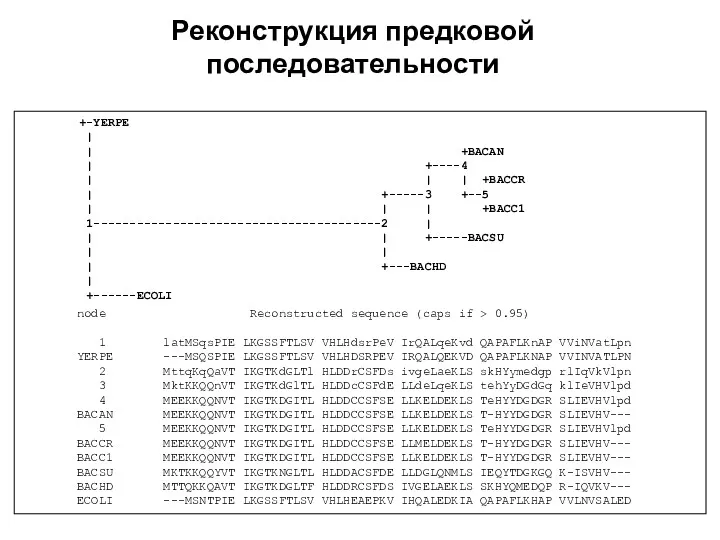
Реконструкция предковой последовательности
+-YERPE
|
| +BACAN
|
Реконструкция предковой последовательности
+-YERPE
|
| +BACAN
|

Реконструкция филогении
по биологическим последовательностям
Реконструкция филогении
по биологическим последовательностям
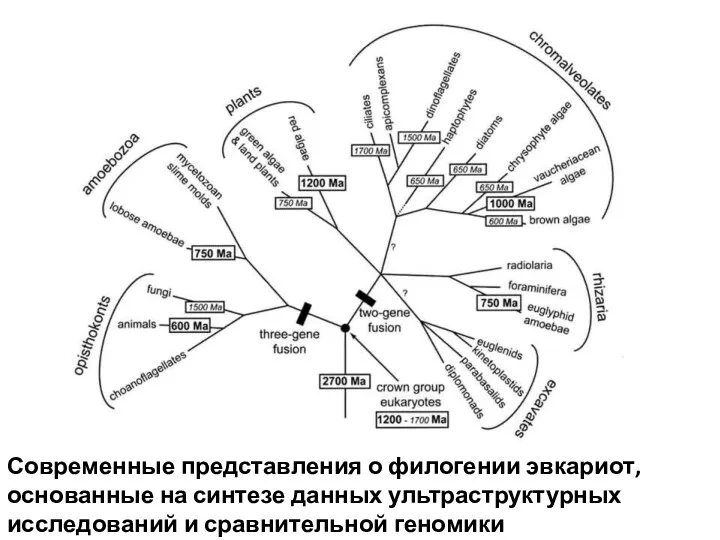
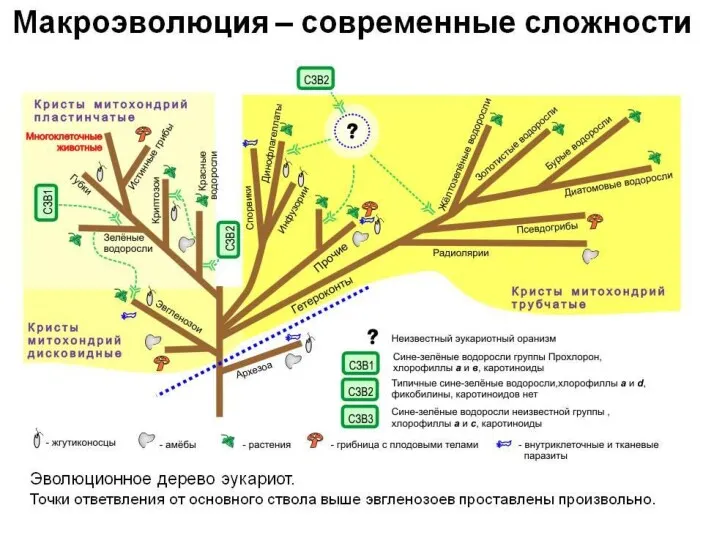
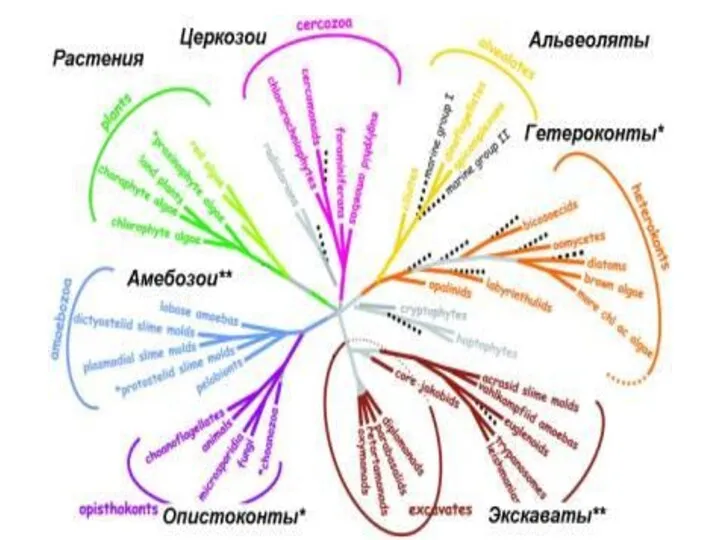
Современные представления о филогении эвкариот, основанные на синтезе данных ультраструктурных исследований
Современные представления о филогении эвкариот, основанные на синтезе данных ультраструктурных исследований
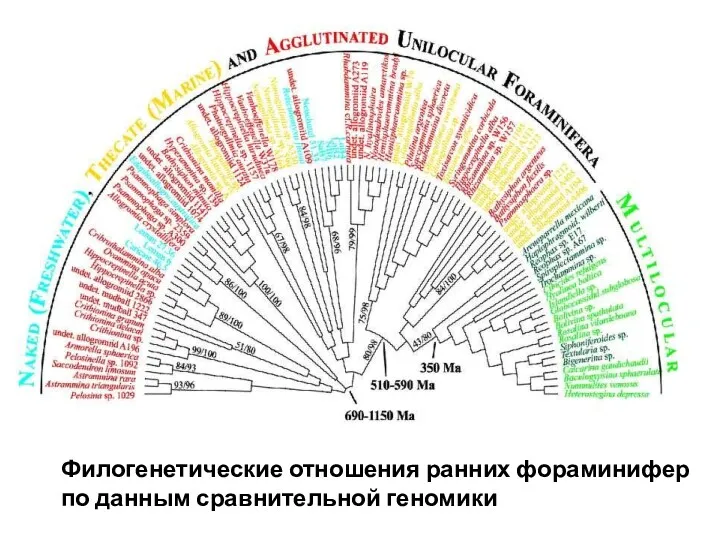
Филогенетические отношения ранних фораминифер
по данным сравнительной геномики
Филогенетические отношения ранних фораминифер
по данным сравнительной геномики
 Класс Млекопитающие (Звери)
Класс Млекопитающие (Звери) Нервная система
Нервная система Проект Геном человека - занятие по биохимии
Проект Геном человека - занятие по биохимии Отдел Мхи
Отдел Мхи Закономерности наследования признаков, установленные Г. Менделем. Моногибридное скрещивание. Первый и второй законы
Закономерности наследования признаков, установленные Г. Менделем. Моногибридное скрещивание. Первый и второй законы Поток энергии и цепи питания.
Поток энергии и цепи питания. Двигательные системы
Двигательные системы Экскурсия в науку (по кабинету биологии)
Экскурсия в науку (по кабинету биологии) Сущность жизни и свойства живого
Сущность жизни и свойства живого Қазақстандағы дәрілік өсімдіктер
Қазақстандағы дәрілік өсімдіктер Алгоритм решения задания №27 на антипараллельность
Алгоритм решения задания №27 на антипараллельность железы внутренней секреции
железы внутренней секреции Популяциядағы жүретін процестерді зерттеу
Популяциядағы жүретін процестерді зерттеу Різновиди хижих рослин
Різновиди хижих рослин Мейоз
Мейоз Как размножаются живые организмы
Как размножаются живые организмы Устройство пищевода
Устройство пищевода Физиология и экология диатомовых водорослей
Физиология и экология диатомовых водорослей Геоботаникаю лекция 2
Геоботаникаю лекция 2 Выступление на всероссийском семинаре Личностно- ориентированнный подход при создании проектных работ.
Выступление на всероссийском семинаре Личностно- ориентированнный подход при создании проектных работ. Домашние животные и птицы
Домашние животные и птицы Кальциевые каналы
Кальциевые каналы Сексована сперма
Сексована сперма Физиология сенсорных систем
Физиология сенсорных систем Наши домашние питомцы (1 класс)
Наши домашние питомцы (1 класс) Вода. Свойства воды. Тест
Вода. Свойства воды. Тест Сущность жизни и свойства живого
Сущность жизни и свойства живого Мезозойская эра
Мезозойская эра