- Нервная система

Содержание
- 2. ТРИ ОСНОВНЫХ ФУНКЦИОНАЛЬНЫХ ОТДЕЛА ЦНС СОМАТИЧЕСКАЯ (двигательная) СИСТЕМА ВЕГЕТАТИВНАЯ СИСТЕМА: СИМПАТИЧЕСКИЙ ОТДЕЛ ПАРАСИМПАТИЧЕСКИЙ ОТДЕЛ СЕНСОРНАЯ СИСТЕМА
- 3. ОСНОВНЫЕ ФУНКЦИИ ЦНС Регуляция двигательных функций Регуляция функций внутренних органов Восприятие, переработка и хранение информации Осуществление
- 4. ОСНОВНОЙ ПРИНЦИП ДЕЯТЕЛЬНОСТИ ЦНС РЕФЛЕКТОРНЫЙ РЕФЛЕКС – ответная реакция организма на раздражители внешней и внутренней среды
- 5. РЕФЛЕКТОРНАЯ ДУГА - совокупность структур, необходимых для осуществления рефлекса РЕФЛЕКТОРНАЯ ДУГА СОСТОИТ ИЗ ПЯТИ ЧАСТЕЙ: Сенсорный
- 6. Перифериче- ский ганглий Перифериче- ский ганглий
- 7. ФИЗИОЛОГИЯ НЕЙРОНА
- 8. Афферентные (чувствительные) нейроны Эфферентные (двигательные) нейроны Вставочные нейроны Рецепторы (сенсоры) 50 тысяч разных типов нейронов в
- 9. КЛАССИФИКАЦИЯ НЕЙРОНОВ Учитывая размеры и форму: большие пирамидные, малые пирамидные и др. По количеству отростков: униполярные,
- 10. NEURON ТЕЛО: метаболизм, синтез, восприятие сигналов ДЕНДРИТЫ: восприятие сигналов АКСОННЫЙ ХОЛМИК: генерация ПД АКСОН: передача импульсов
- 11. СВЯЗЬ МЕЖДУ НЕЙРОНАМИ с помощью химических синапсов (от 200 до 200 000 синапсов). 95% синапсов расположено
- 12. КЛАССИФИКАЦИЯ СИНАПСОВ По локализации: аксо-сомальные, аксо-дендритные, аксо-аксональные и др. По типу влияния: возбуждающие и тормозные По
- 13. ЭЛЕКТРИЧЕСКИЙ СИНАПС МЕДИАТОР ИОНЫ Ионные каналы ПД Ионные токи ХИМИЧЕСКИЙ СИНАПС
- 14. СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА ЭЛЕКТРИЧЕСКИХ И ХИМИЧЕСКИХ СИНАПСОВ ЭЛЕКТРИЧЕСКИЙ СИНАПС ХИМИЧЕСКИЙ СИНАПС Без задержки Синаптическая задержка Без утомления
- 15. 1 синапс на мыш.волокне Большая площадь синаптического контакта 200 везикул медиатора на 1 ПД ПКП всегда
- 16. МЕХАНИЗМ ВОЗБУЖДЕНИЯ НЕЙРОНА
- 17. ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ (ВПСП) Свойства ВПСП: Зависит от количества медиатора Способен к суммации Распространяется с затуханием
- 18. РЕАКЦИЯ НЕЙРОНА НА ОДИНОЧНЫЙ НЕРВНЫЙ ИМПУЛЬС Слишком слабая деполяризация постсинаптической мембраны (0,1-1 мВ). ПД не возникает.
- 19. РЕАКЦИЯ НЕЙРОНА НА СЕРИЮ НЕРВНЫХ ИМПУЛЬСОВ ВПСП распространяется с затуханием, вызывает допороговую деполяризацию аксонного холмика. ПД
- 20. РЕАКЦИЯ НЕЙРОНА НА ВПСП, ВОЗНИКШИЕ ОДНОВРЕМЕННО В РАЗНЫХ СИНАПСАХ Множество ВПСП, возникших одновременно в разных участках
- 21. Чем выше деполяризация аксонного холмика, тем больше частота импульсов Критический уровень деполяризации
- 22. СВОЙСТВА ТПСП: Зависит от количества медиатора 2. Способен к суммации 3. Распространяется с затуханием 4. Уменьшает
- 23. ПОСТСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ Сущность постсинаптического торможения – гиперполяризация нервной клетки. Гиперполяризация возникает за счёт входа в клетку
- 24. ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ Торможение развивается в пресинаптическом нервном окончании возбуждающего нейрона (аксо-аксональный синапс). ПД не может пройти
- 25. ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ Пресинаптическое торможение развивается за счёт длительной стойкой деполяризации постсинаптической мембраны в аксо-аксональном синапсе. Длительная
- 26. ВЫВОД ТОРМОЖЕНИЕ – это активный нервный процесс, который направлен на прекращение генерации импульсов и (или) выделения
- 27. НЕЙРОМЕДИАТОРЫ
- 28. Высокомолекулрные, долговременного действия: НЕЙРОПЕПТИДЫ Гипоталамические Гипофизарные Гастро-интестинальные и другие Низкомолекулярные, кратковременного действия: АЦЕТИЛХОЛИН АМИНЫ АМИНОКИСЛОТЫ ОКСИД
- 29. МОЛЕКУЛЯРНЫЕ РЕЦЕПТОРЫ ПОСТСИНАПТИЧЕСКИХ МЕМБРАН ИОНОТРОПНЫЕ (связанные с хемочувствительными ионными каналами) Р Р ИОННЫЙ КАНАЛ ЗАКРЫТ ИОННЫЙ
- 30. МОЛЕКУЛЯРНЫЕ РЕЦЕПТОРЫ ПОСТСИНАПТИЧЕСКИХ МЕМБРАН МЕТАБОТРОПНЫЕ (связанные с системой вторых посредников) С метаботропными рецепторами взаимодействуют: катехоламины, серотонин,
- 32. НЕРВНЫЙ ЦЕНТР – скопление нейронов, которые участвуют в регуляции одной и той же функции организма. Анатомически
- 33. СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ ЗАВИСЯТ: От свойств нервных клеток. От свойств химических синапсов. От сочетания возбуждающих и
- 34. ОСОБЕННОСТИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ПО РЕФЛЕКТОРНОЙ ДУГЕ (по сравнению с нервным волокном) Одностороннее проведение: за счёт 1-стороннего
- 35. СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ Суммация возбуждения (временная и пространственная) Инерционность Последействие Пластичность Фоновая активность, тонус Высокая утомляемость
- 36. ЭЛЕМЕНТАРНЫЕ НЕЙРОННЫЕ КОНТУРЫ ДИВЕРГЕНЦИЯ Дивергенция приводит к распространению (иррадиации) возбуждения по всем отделам ЦНС
- 37. КОНВЕРГЕНЦИЯ Конвергенция лежит в основе простран- ственной суммации возбуждения, ведёт к трансформации ритма возбуждения
- 38. КОЛЬЦЕВЫЕ НЕЙРОННЫЕ ЦЕПИ Благодаря циркуляции возбуждения по замкнутым нейронным цепям, происходит усиление импульсации. Возбуждение нейронов продолжается
- 39. ВОЗВРАТНОЕ ПОСТСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ В замкнутой цепи нейронов с возвратным постсинаптическим торможением не только снижается возбудимость нейрона
- 40. Суммация ВПСП и ТПСП при возвратном постсинаптическом торможении нейрона ВПСП ТПСП Результат суммации (аксонный холмик) Импульсация
- 41. ВОЗВРАТНОЕ ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ С помощью возвратного пресинаптического торможения происходит ограничение потока импульсов, поступающих к отдельному нейрону
- 42. РЕЦИПРОКНОЕ ТОРМОЖЕНИЕ К сгибательным мышцам К разгибательным мышцам Нет импульсов Благодаря реципрокному (сопряжённому) торможению, при возбуждении
- 45. КООРДИНАЦИЯ ДЕЯТЕЛЬНОСТИ ЦНС ПРИНЦИП СУБОРДИНАЦИИ (или иерархии) СПИННОЙ МОЗГ СТВОЛ ГОЛОВНОГО МОЗГА КОРА БОЛЬШИХ ПОЛУШАРИЙ
- 46. II. ПРИНЦИП ОБРАТНОЙ СВЯЗИ КОРА БОЛЬШИХ ПОЛУШАРИЙ СТВОЛ ГОЛОВНОГО МОЗГА СПИННОЙ МОЗГ ВТОРИЧНАЯ АФФЕРЕНТАЦИЯ КОПИЯ ЭФФЕРЕНТАЦИИ
- 47. III. ПРИНЦИП ОБЛЕГЧЕНИЯ И ОККЛЮЗИИ (В СИНЕРГИЧЕСКИХ НЕРВНЫХ ЦЕНТРАХ) ОБЛЕГЧЕНИЕ Афферентный вход А допороговый Не вызывает
- 48. ОБЛЕГЧЕНИЕ Афферентный вход А пороговый Возбуждает 1 группу нейронов Афферентный вход Б пороговый Возбуждает 1 группу
- 49. Суммарный ответ меньше, чем простая сумма двух отдельных ответных реакций (за счёт конвергенции возбуждения к одним
- 50. IV. ПРИНЦИП РЕЦИПРОКНОЙ ИННЕРВАЦИИ СПИННОЙ МОЗГ вход возбуждающий К сгибательным мышцам Отсутствие импульсации к разгибательным мышцам
- 51. V. ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ (ОКП), ФАКТОР СИЛЫ В БОРЬБЕ ЗА ОКП Конвергенция импульсов от разных
- 52. Доминантный центр «притягивает» к себе возбуждение, возникшее в других центрах. Из всех возможных ответных реакций в
- 53. МЕХАНИЗМ ФОРМИРОВАНИЯ ДОМИНАНТЫ (СХЕМА ФУНКЦИОНАЛЬНОЙ СИСТЕМЫ) П.К. АНОХИН ОБСТАНОВОЧ- НАЯ АФФЕРЕНТАЦИЯ ВНУТРЕННЯЯ МОТИВАЦИЯ ПУСКОВАЯ АФФЕРЕНТАЦИЯ ПАМЯТЬ
- 55. Скачать презентацию

ТРИ ОСНОВНЫХ ФУНКЦИОНАЛЬНЫХ ОТДЕЛА ЦНС
СОМАТИЧЕСКАЯ (двигательная) СИСТЕМА
ВЕГЕТАТИВНАЯ СИСТЕМА:
СИМПАТИЧЕСКИЙ ОТДЕЛ
ПАРАСИМПАТИЧЕСКИЙ ОТДЕЛ
СЕНСОРНАЯ СИСТЕМА
ТРИ ОСНОВНЫХ ФУНКЦИОНАЛЬНЫХ ОТДЕЛА ЦНС
СОМАТИЧЕСКАЯ (двигательная) СИСТЕМА
ВЕГЕТАТИВНАЯ СИСТЕМА:
СИМПАТИЧЕСКИЙ ОТДЕЛ
ПАРАСИМПАТИЧЕСКИЙ ОТДЕЛ
СЕНСОРНАЯ СИСТЕМА

ОСНОВНЫЕ ФУНКЦИИ ЦНС
Регуляция двигательных функций
Регуляция функций внутренних органов
Восприятие, переработка и хранение
ОСНОВНЫЕ ФУНКЦИИ ЦНС
Регуляция двигательных функций
Регуляция функций внутренних органов
Восприятие, переработка и хранение

ОСНОВНОЙ ПРИНЦИП ДЕЯТЕЛЬНОСТИ ЦНС
РЕФЛЕКТОРНЫЙ
РЕФЛЕКС – ответная реакция организма на раздражители
ОСНОВНОЙ ПРИНЦИП ДЕЯТЕЛЬНОСТИ ЦНС
РЕФЛЕКТОРНЫЙ
РЕФЛЕКС – ответная реакция организма на раздражители

РЕФЛЕКТОРНАЯ ДУГА -
совокупность структур, необходимых для осуществления рефлекса
РЕФЛЕКТОРНАЯ ДУГА
СОСТОИТ
РЕФЛЕКТОРНАЯ ДУГА -
совокупность структур, необходимых для осуществления рефлекса
РЕФЛЕКТОРНАЯ ДУГА
СОСТОИТ
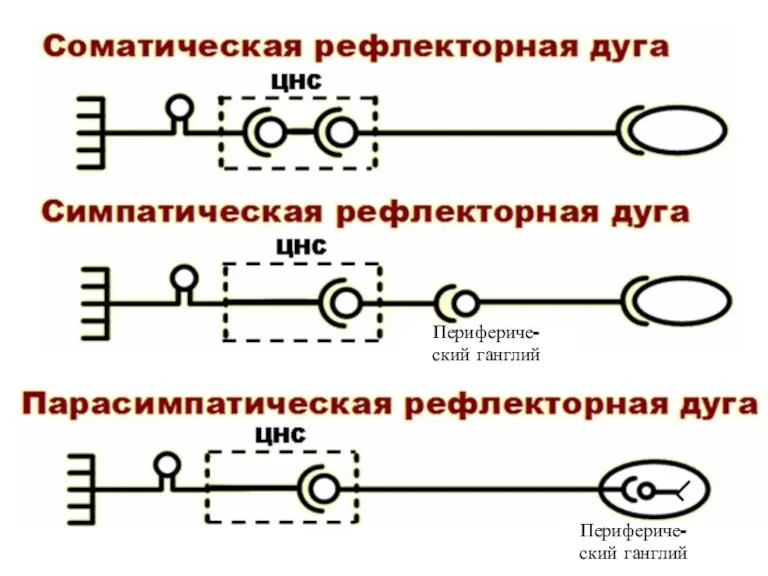
Перифериче-
ский ганглий
Перифериче-
ский ганглий
Перифериче-
ский ганглий
Перифериче-
ский ганглий

ФИЗИОЛОГИЯ
НЕЙРОНА
ФИЗИОЛОГИЯ
НЕЙРОНА
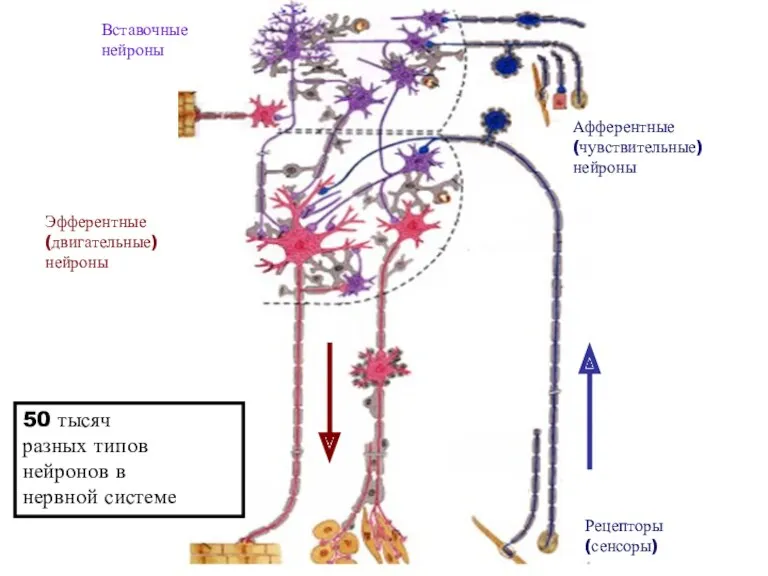
Афферентные
(чувствительные)
нейроны
Эфферентные
(двигательные)
нейроны
Вставочные
нейроны
Рецепторы
(сенсоры)
50 тысяч
разных типов
нейронов в
нервной системе
Афферентные
(чувствительные)
нейроны
Эфферентные
(двигательные)
нейроны
Вставочные
нейроны
Рецепторы
(сенсоры)
50 тысяч
разных типов
нейронов в
нервной системе

КЛАССИФИКАЦИЯ НЕЙРОНОВ
Учитывая размеры и форму:
большие пирамидные, малые пирамидные и др.
По количеству
КЛАССИФИКАЦИЯ НЕЙРОНОВ
Учитывая размеры и форму:
большие пирамидные, малые пирамидные и др.
По количеству
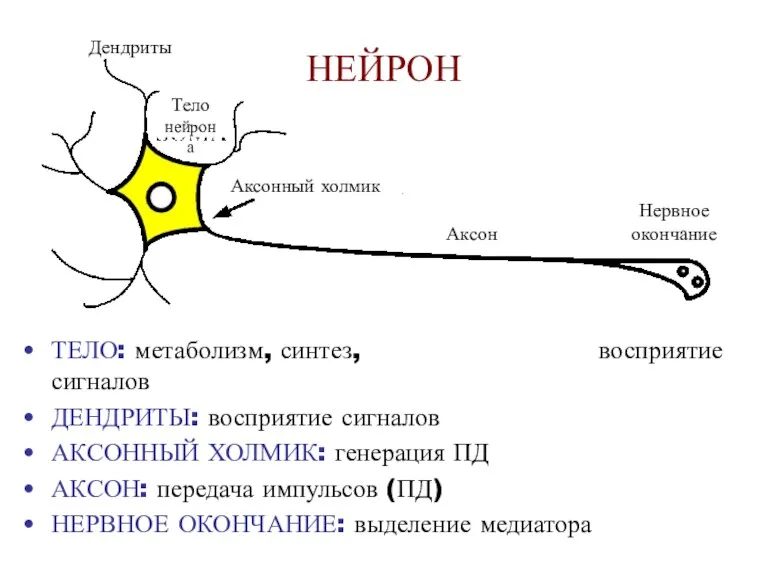
NEURON
ТЕЛО: метаболизм, синтез, восприятие сигналов
ДЕНДРИТЫ: восприятие сигналов
АКСОННЫЙ ХОЛМИК: генерация ПД
АКСОН:
NEURON
ТЕЛО: метаболизм, синтез, восприятие сигналов
ДЕНДРИТЫ: восприятие сигналов
АКСОННЫЙ ХОЛМИК: генерация ПД
АКСОН:
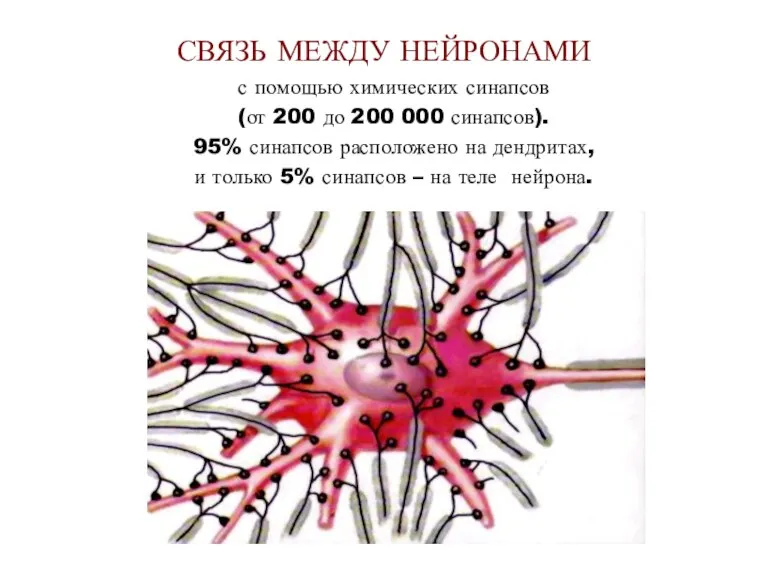
СВЯЗЬ МЕЖДУ НЕЙРОНАМИ
с помощью химических синапсов
(от 200 до 200
СВЯЗЬ МЕЖДУ НЕЙРОНАМИ
с помощью химических синапсов
(от 200 до 200

КЛАССИФИКАЦИЯ СИНАПСОВ
По локализации:
аксо-сомальные, аксо-дендритные, аксо-аксональные и др.
По типу влияния:
возбуждающие и тормозные
По
КЛАССИФИКАЦИЯ СИНАПСОВ
По локализации:
аксо-сомальные, аксо-дендритные, аксо-аксональные и др.
По типу влияния:
возбуждающие и тормозные
По
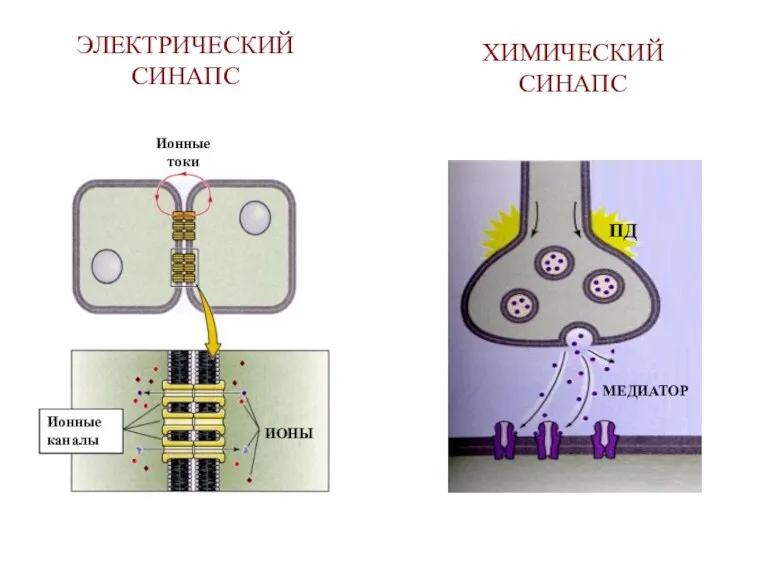
ЭЛЕКТРИЧЕСКИЙ СИНАПС
МЕДИАТОР
ИОНЫ
Ионные
каналы
ПД
Ионные
токи
ХИМИЧЕСКИЙ
СИНАПС
ЭЛЕКТРИЧЕСКИЙ СИНАПС
МЕДИАТОР
ИОНЫ
Ионные
каналы
ПД
Ионные
токи
ХИМИЧЕСКИЙ
СИНАПС

СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА
ЭЛЕКТРИЧЕСКИХ И ХИМИЧЕСКИХ СИНАПСОВ
ЭЛЕКТРИЧЕСКИЙ СИНАПС ХИМИЧЕСКИЙ СИНАПС
Без задержки Синаптическая задержка
Без утомления Быстро утомляются
Слабо чувствительны
СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА
ЭЛЕКТРИЧЕСКИХ И ХИМИЧЕСКИХ СИНАПСОВ
ЭЛЕКТРИЧЕСКИЙ СИНАПС ХИМИЧЕСКИЙ СИНАПС
Без задержки Синаптическая задержка
Без утомления Быстро утомляются
Слабо чувствительны

1 синапс на мыш.волокне
Большая площадь синаптического контакта
200 везикул медиатора на 1
1 синапс на мыш.волокне
Большая площадь синаптического контакта
200 везикул медиатора на 1

МЕХАНИЗМ
ВОЗБУЖДЕНИЯ НЕЙРОНА
МЕХАНИЗМ
ВОЗБУЖДЕНИЯ НЕЙРОНА
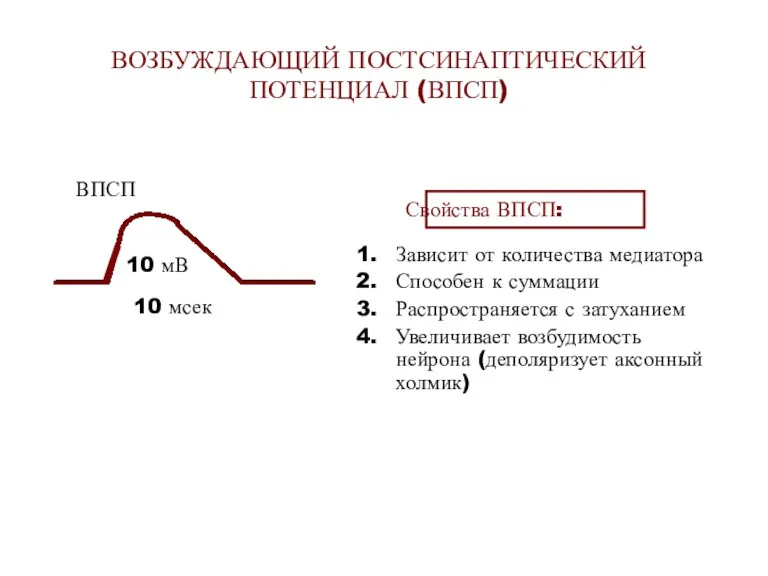
ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ (ВПСП)
Свойства ВПСП:
Зависит от количества медиатора
Способен к суммации
Распространяется с
ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ (ВПСП)
Свойства ВПСП:
Зависит от количества медиатора
Способен к суммации
Распространяется с
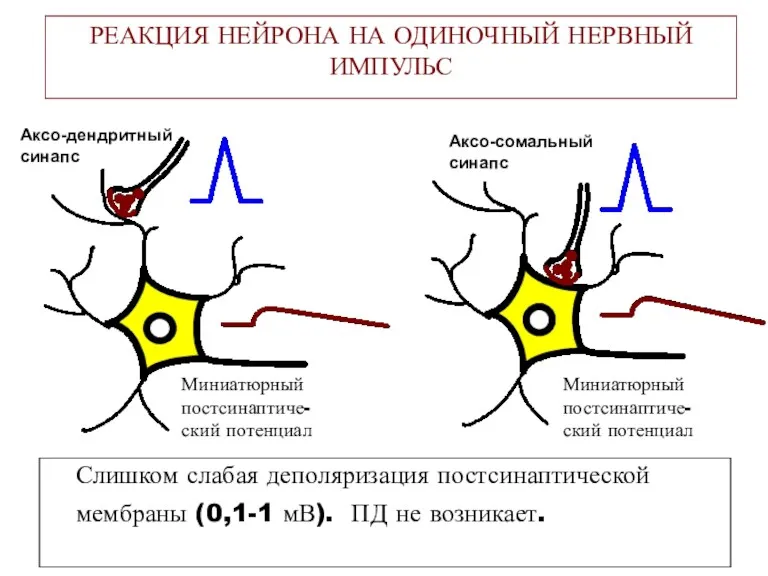
РЕАКЦИЯ НЕЙРОНА НА ОДИНОЧНЫЙ НЕРВНЫЙ ИМПУЛЬС
Слишком слабая деполяризация постсинаптической мембраны (0,1-1
РЕАКЦИЯ НЕЙРОНА НА ОДИНОЧНЫЙ НЕРВНЫЙ ИМПУЛЬС
Слишком слабая деполяризация постсинаптической мембраны (0,1-1
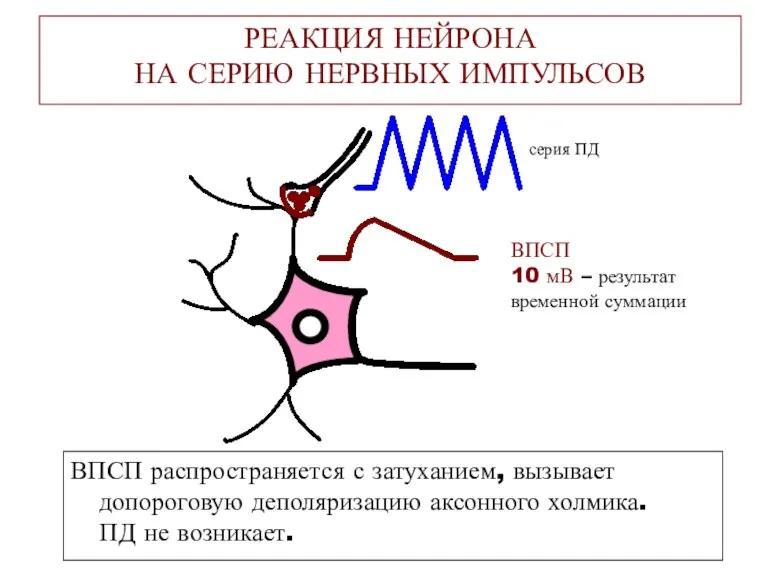
РЕАКЦИЯ НЕЙРОНА
НА СЕРИЮ НЕРВНЫХ ИМПУЛЬСОВ
ВПСП распространяется с затуханием, вызывает допороговую
РЕАКЦИЯ НЕЙРОНА
НА СЕРИЮ НЕРВНЫХ ИМПУЛЬСОВ
ВПСП распространяется с затуханием, вызывает допороговую
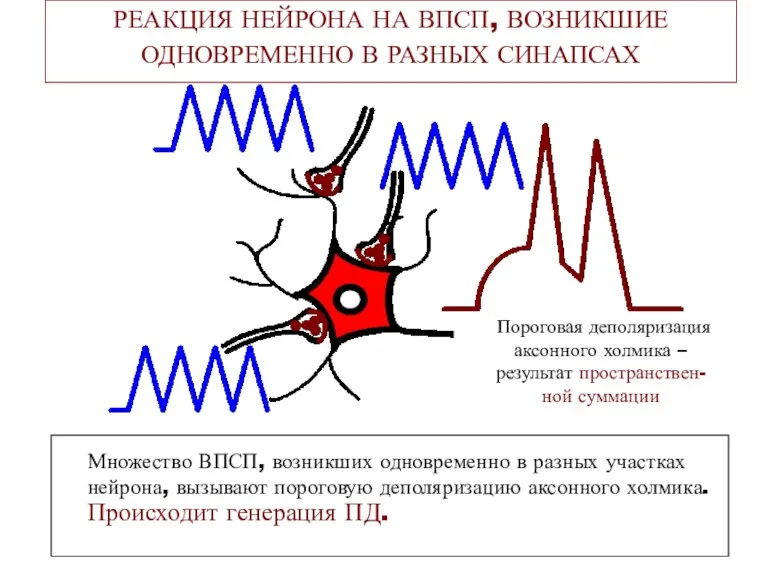
РЕАКЦИЯ НЕЙРОНА НА ВПСП, ВОЗНИКШИЕ ОДНОВРЕМЕННО В РАЗНЫХ СИНАПСАХ
Множество ВПСП,
РЕАКЦИЯ НЕЙРОНА НА ВПСП, ВОЗНИКШИЕ ОДНОВРЕМЕННО В РАЗНЫХ СИНАПСАХ
Множество ВПСП,
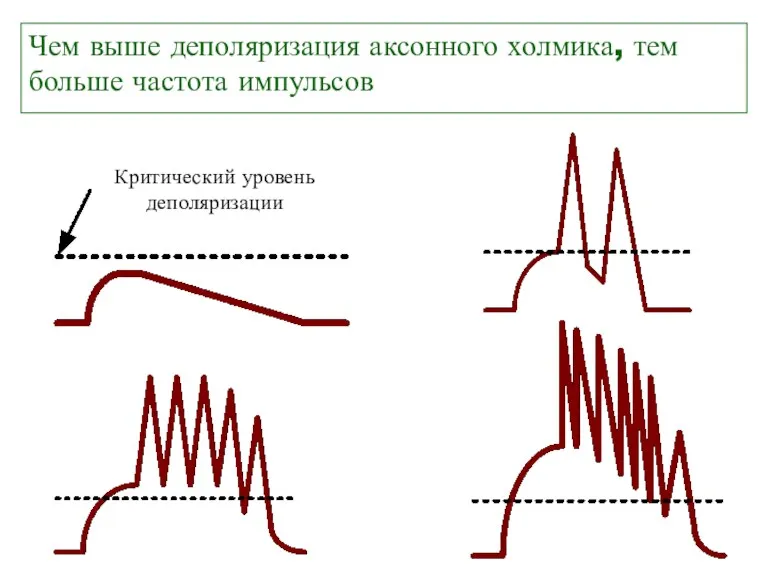
Чем выше деполяризация аксонного холмика, тем больше частота импульсов
Критический уровень
деполяризации
Чем выше деполяризация аксонного холмика, тем больше частота импульсов
Критический уровень
деполяризации
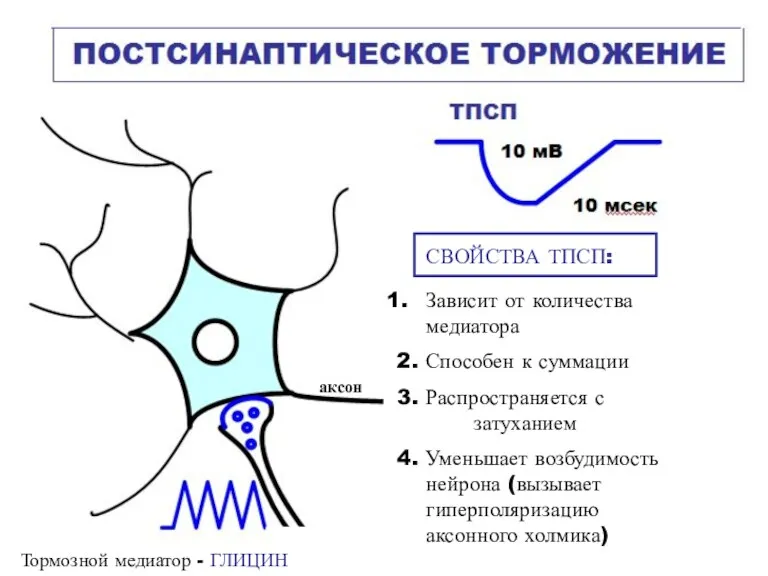
СВОЙСТВА ТПСП:
Зависит от количества
медиатора
2. Способен к суммации
3. Распространяется с
затуханием
4. Уменьшает
СВОЙСТВА ТПСП:
Зависит от количества
медиатора
2. Способен к суммации
3. Распространяется с
затуханием
4. Уменьшает

ПОСТСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
Сущность постсинаптического торможения – гиперполяризация нервной клетки.
Гиперполяризация возникает за счёт
ПОСТСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
Сущность постсинаптического торможения – гиперполяризация нервной клетки.
Гиперполяризация возникает за счёт
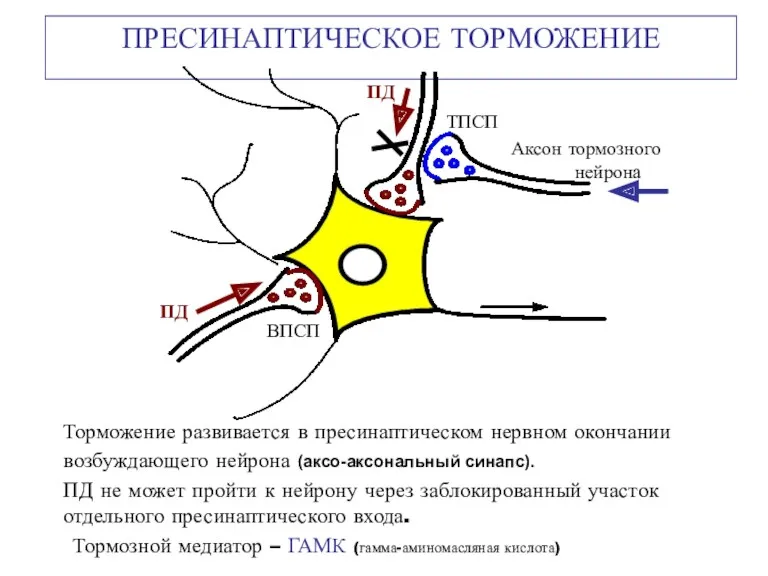
ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
Торможение развивается в пресинаптическом нервном окончании возбуждающего нейрона (аксо-аксональный синапс).
ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
Торможение развивается в пресинаптическом нервном окончании возбуждающего нейрона (аксо-аксональный синапс).

ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
Пресинаптическое торможение развивается за счёт длительной стойкой деполяризации постсинаптической мембраны
ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
Пресинаптическое торможение развивается за счёт длительной стойкой деполяризации постсинаптической мембраны

ВЫВОД
ТОРМОЖЕНИЕ – это активный нервный процесс, который направлен на прекращение
ВЫВОД
ТОРМОЖЕНИЕ – это активный нервный процесс, который направлен на прекращение

НЕЙРОМЕДИАТОРЫ
НЕЙРОМЕДИАТОРЫ
Высокомолекулрные, долговременного действия:
НЕЙРОПЕПТИДЫ
Гипоталамические
Гипофизарные
Гастро-интестинальные
и другие
Низкомолекулярные, кратковременного действия:
АЦЕТИЛХОЛИН
АМИНЫ
АМИНОКИСЛОТЫ
ОКСИД АЗОТА (NO)
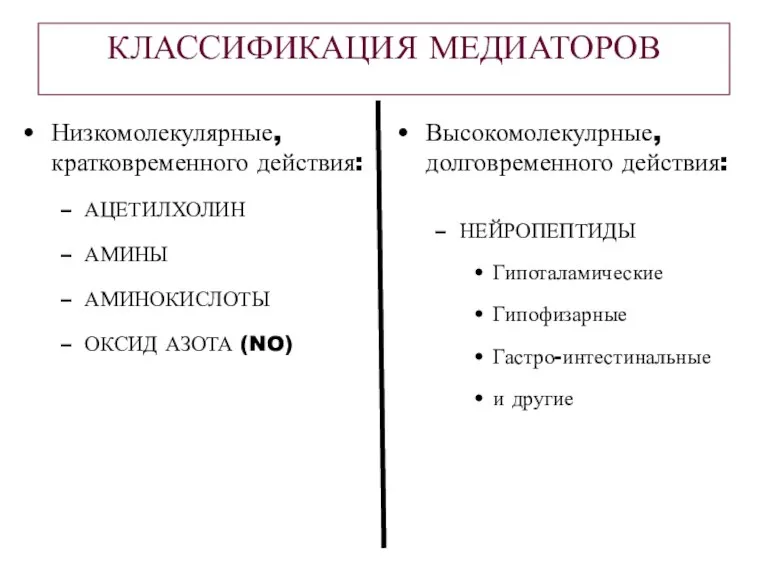
КЛАССИФИКАЦИЯ МЕДИАТОРОВ
Высокомолекулрные, долговременного действия:
НЕЙРОПЕПТИДЫ
Гипоталамические
Гипофизарные
Гастро-интестинальные
и другие
Низкомолекулярные, кратковременного действия:
АЦЕТИЛХОЛИН
АМИНЫ
АМИНОКИСЛОТЫ
ОКСИД АЗОТА (NO)
КЛАССИФИКАЦИЯ МЕДИАТОРОВ
МОЛЕКУЛЯРНЫЕ РЕЦЕПТОРЫ
ПОСТСИНАПТИЧЕСКИХ МЕМБРАН
ИОНОТРОПНЫЕ
(связанные с хемочувствительными ионными каналами)
Р
Р
ИОННЫЙ
КАНАЛ
ЗАКРЫТ
ИОННЫЙ
КАНАЛ
ОТКРЫТ
С ионотропными рецепторами
заимодействуют:
ацетилхолин,
глутамат,
аспартат,
глицин,
ГАМК
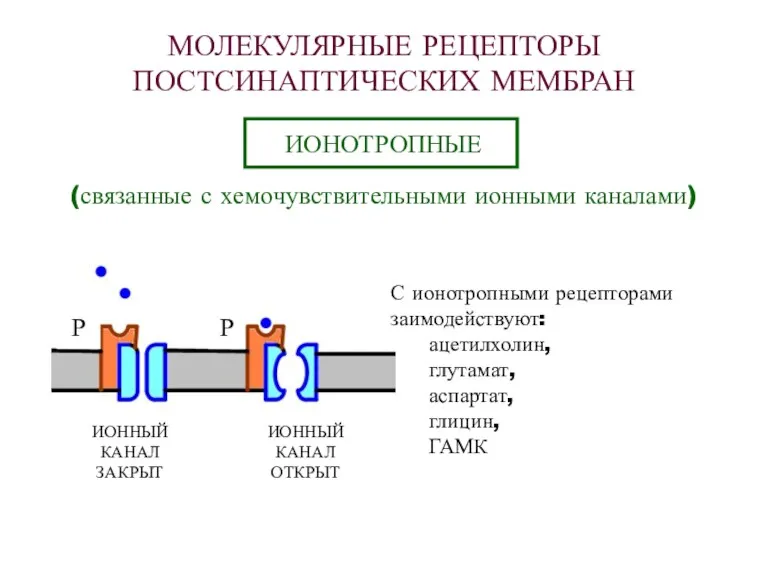
МОЛЕКУЛЯРНЫЕ РЕЦЕПТОРЫ
ПОСТСИНАПТИЧЕСКИХ МЕМБРАН
ИОНОТРОПНЫЕ
(связанные с хемочувствительными ионными каналами)
Р
Р
ИОННЫЙ
КАНАЛ
ЗАКРЫТ
ИОННЫЙ
КАНАЛ
ОТКРЫТ
С ионотропными рецепторами
заимодействуют:
ацетилхолин,
глутамат,
аспартат,
глицин,
ГАМК

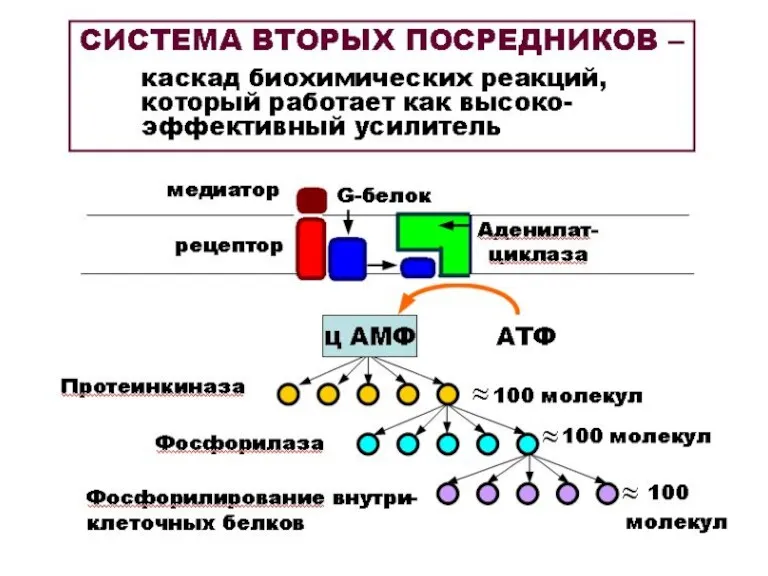
МОЛЕКУЛЯРНЫЕ РЕЦЕПТОРЫ
ПОСТСИНАПТИЧЕСКИХ МЕМБРАН
МЕТАБОТРОПНЫЕ
(связанные с системой
вторых посредников)
С метаботропными рецепторами
взаимодействуют:
катехоламины,
серотонин,
ГАМК,
опиоиды,
нейропептиды
МОЛЕКУЛЯРНЫЕ РЕЦЕПТОРЫ
ПОСТСИНАПТИЧЕСКИХ МЕМБРАН
МЕТАБОТРОПНЫЕ
(связанные с системой
вторых посредников)
С метаботропными рецепторами
взаимодействуют:
катехоламины,
серотонин,
ГАМК,
опиоиды,
нейропептиды

НЕРВНЫЙ ЦЕНТР –
скопление нейронов, которые участвуют в регуляции одной и
НЕРВНЫЙ ЦЕНТР –
скопление нейронов, которые участвуют в регуляции одной и

СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
ЗАВИСЯТ:
От свойств нервных клеток.
От свойств химических синапсов.
От сочетания возбуждающих
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
ЗАВИСЯТ:
От свойств нервных клеток.
От свойств химических синапсов.
От сочетания возбуждающих

ОСОБЕННОСТИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ПО РЕФЛЕКТОРНОЙ ДУГЕ
(по сравнению с нервным волокном)
Одностороннее проведение:
за
ОСОБЕННОСТИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ПО РЕФЛЕКТОРНОЙ ДУГЕ
(по сравнению с нервным волокном)
Одностороннее проведение:
за

СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
Суммация возбуждения (временная и пространственная)
Инерционность
Последействие
Пластичность
Фоновая активность, тонус
Высокая утомляемость
Высокая
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
Суммация возбуждения (временная и пространственная)
Инерционность
Последействие
Пластичность
Фоновая активность, тонус
Высокая утомляемость
Высокая
ЭЛЕМЕНТАРНЫЕ НЕЙРОННЫЕ КОНТУРЫ
ДИВЕРГЕНЦИЯ
Дивергенция приводит к распространению
(иррадиации) возбуждения по всем отделам ЦНС
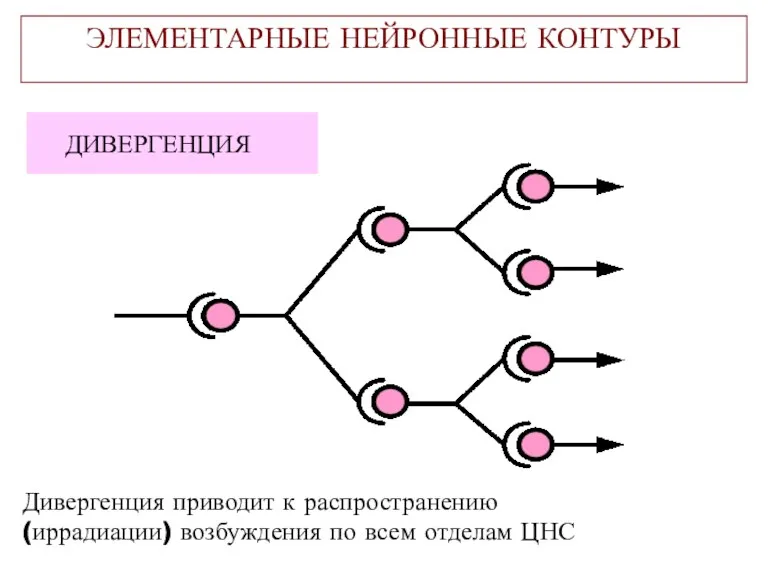
ЭЛЕМЕНТАРНЫЕ НЕЙРОННЫЕ КОНТУРЫ
ДИВЕРГЕНЦИЯ
Дивергенция приводит к распространению
(иррадиации) возбуждения по всем отделам ЦНС
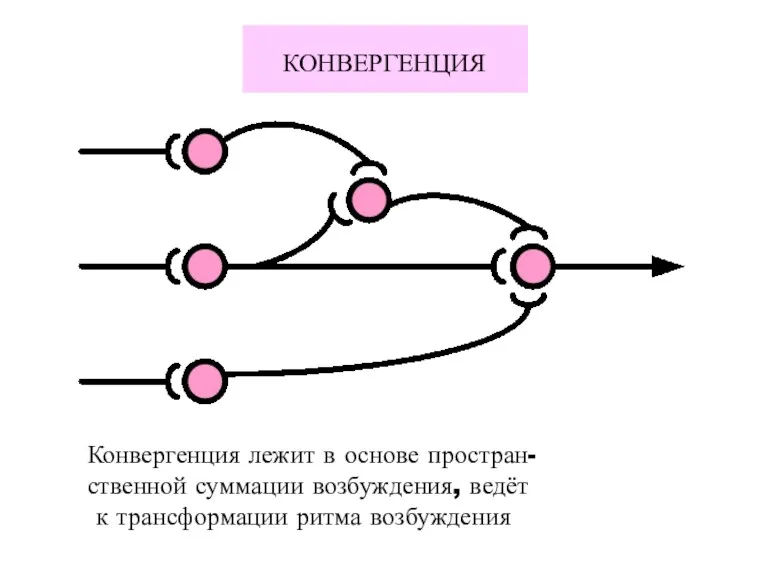
КОНВЕРГЕНЦИЯ
Конвергенция лежит в основе простран-
ственной суммации возбуждения, ведёт
к трансформации ритма
КОНВЕРГЕНЦИЯ
Конвергенция лежит в основе простран-
ственной суммации возбуждения, ведёт
к трансформации ритма
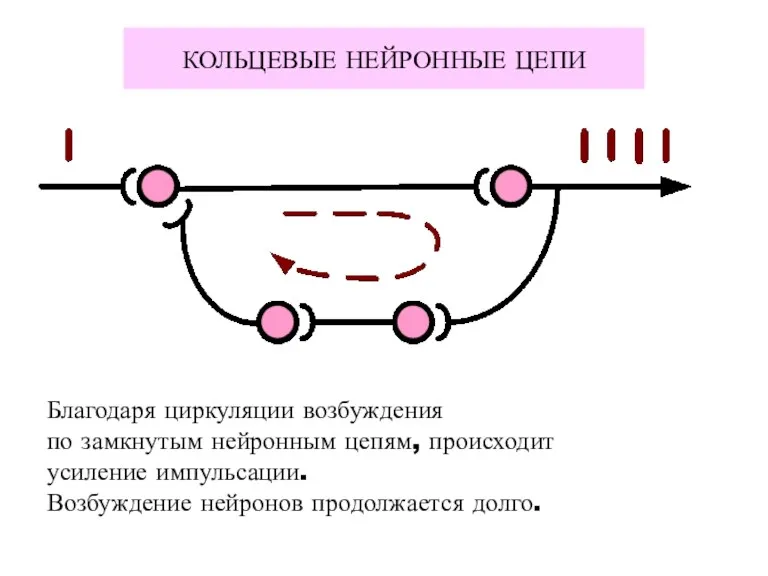
КОЛЬЦЕВЫЕ НЕЙРОННЫЕ ЦЕПИ
Благодаря циркуляции возбуждения
по замкнутым нейронным цепям, происходит
усиление импульсации.
Возбуждение
КОЛЬЦЕВЫЕ НЕЙРОННЫЕ ЦЕПИ
Благодаря циркуляции возбуждения
по замкнутым нейронным цепям, происходит
усиление импульсации.
Возбуждение
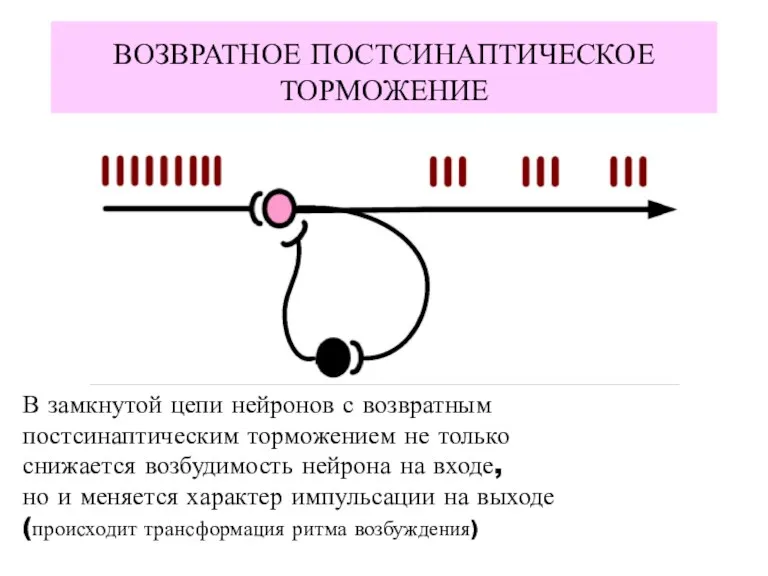
ВОЗВРАТНОЕ ПОСТСИНАПТИЧЕСКОЕ
ТОРМОЖЕНИЕ
В замкнутой цепи нейронов с возвратным
постсинаптическим торможением не только
снижается
ВОЗВРАТНОЕ ПОСТСИНАПТИЧЕСКОЕ
ТОРМОЖЕНИЕ
В замкнутой цепи нейронов с возвратным
постсинаптическим торможением не только
снижается
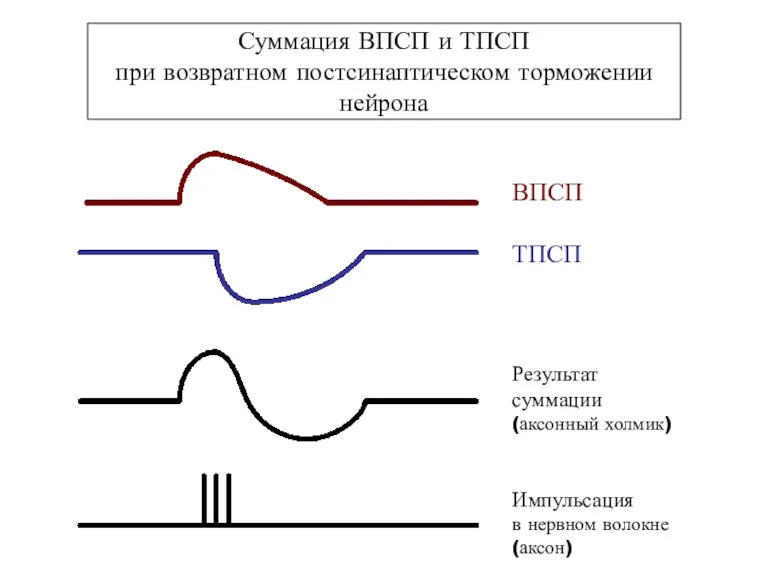
Суммация ВПСП и ТПСП
при возвратном постсинаптическом торможении нейрона
ВПСП
ТПСП
Результат
суммации
(аксонный холмик)
Импульсация
в нервном волокне
(аксон)
Суммация ВПСП и ТПСП
при возвратном постсинаптическом торможении нейрона
ВПСП
ТПСП
Результат
суммации
(аксонный холмик)
Импульсация
в нервном волокне
(аксон)
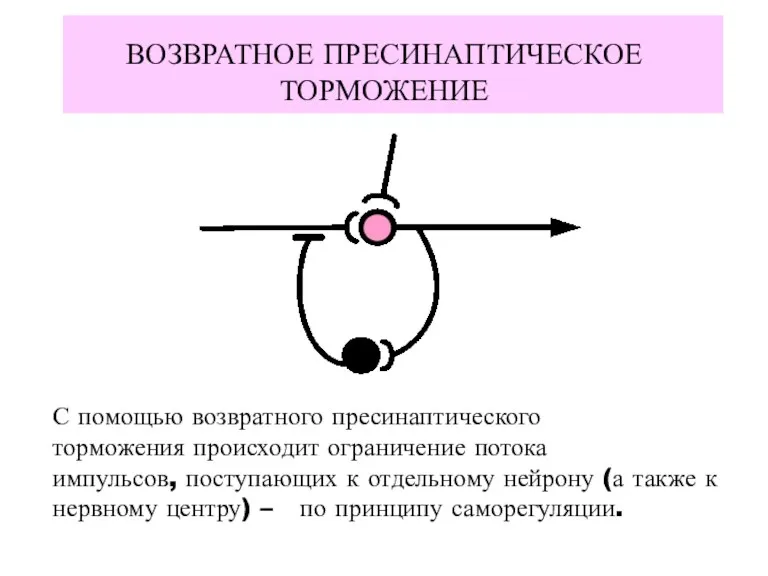
ВОЗВРАТНОЕ ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
С помощью возвратного пресинаптического
торможения происходит ограничение потока
импульсов, поступающих к
ВОЗВРАТНОЕ ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
С помощью возвратного пресинаптического
торможения происходит ограничение потока
импульсов, поступающих к
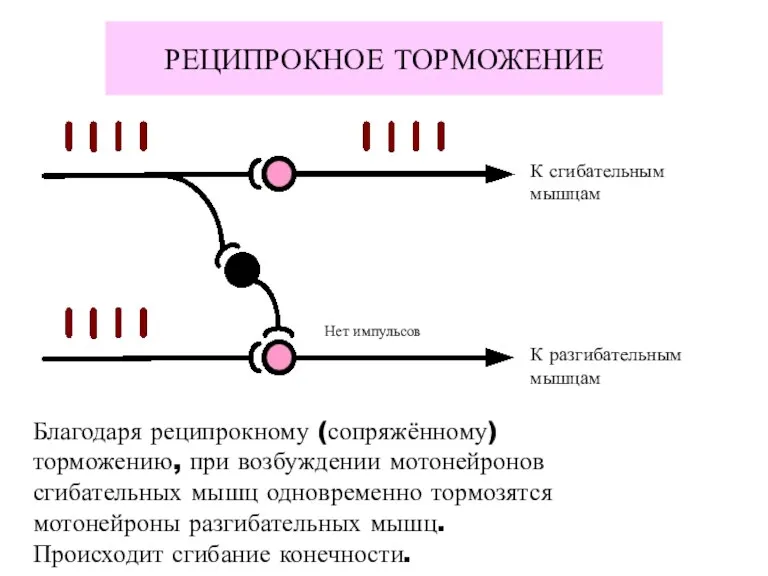
РЕЦИПРОКНОЕ ТОРМОЖЕНИЕ
К сгибательным
мышцам
К разгибательным
мышцам
Нет импульсов
Благодаря реципрокному (сопряжённому)
торможению, при возбуждении мотонейронов
сгибательных
РЕЦИПРОКНОЕ ТОРМОЖЕНИЕ
К сгибательным
мышцам
К разгибательным
мышцам
Нет импульсов
Благодаря реципрокному (сопряжённому)
торможению, при возбуждении мотонейронов
сгибательных
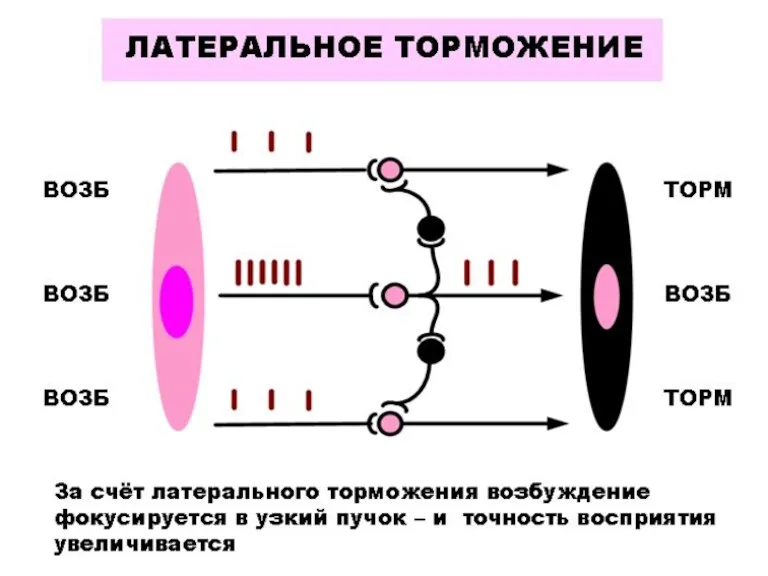
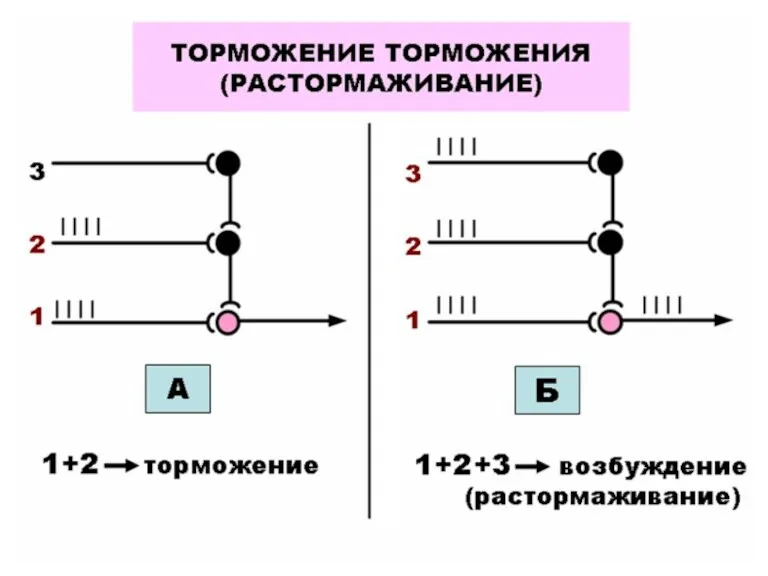
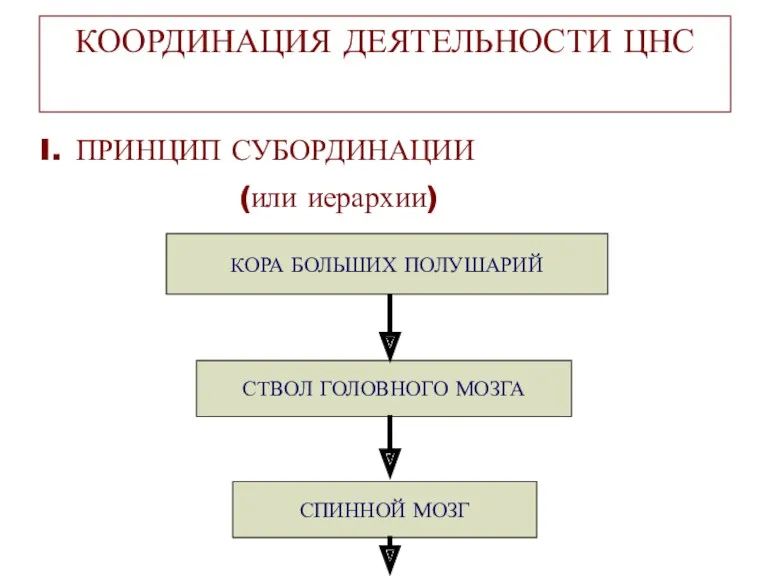
КООРДИНАЦИЯ ДЕЯТЕЛЬНОСТИ ЦНС
ПРИНЦИП СУБОРДИНАЦИИ
(или иерархии)
СПИННОЙ МОЗГ
СТВОЛ ГОЛОВНОГО МОЗГА
КОРА БОЛЬШИХ ПОЛУШАРИЙ
КООРДИНАЦИЯ ДЕЯТЕЛЬНОСТИ ЦНС
ПРИНЦИП СУБОРДИНАЦИИ
(или иерархии)
СПИННОЙ МОЗГ
СТВОЛ ГОЛОВНОГО МОЗГА
КОРА БОЛЬШИХ ПОЛУШАРИЙ
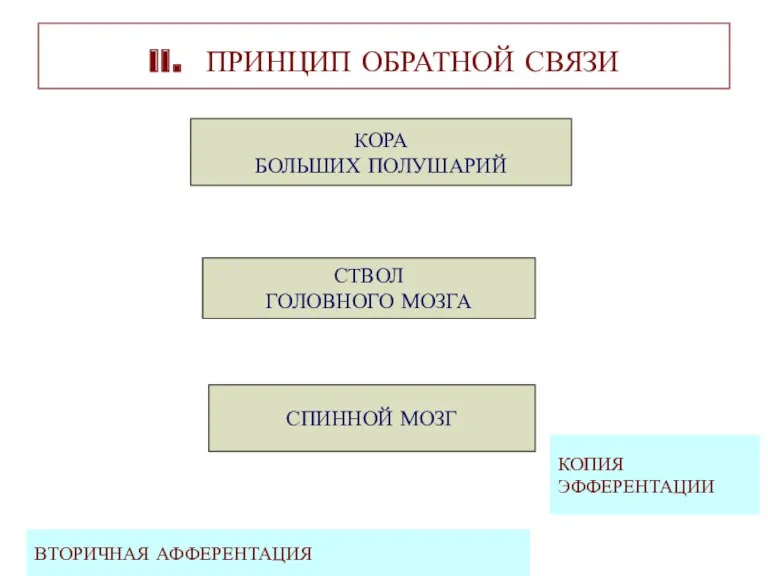
II. ПРИНЦИП ОБРАТНОЙ СВЯЗИ
КОРА
БОЛЬШИХ ПОЛУШАРИЙ
СТВОЛ
ГОЛОВНОГО МОЗГА
СПИННОЙ МОЗГ
ВТОРИЧНАЯ АФФЕРЕНТАЦИЯ
КОПИЯ
ЭФФЕРЕНТАЦИИ
II. ПРИНЦИП ОБРАТНОЙ СВЯЗИ
КОРА
БОЛЬШИХ ПОЛУШАРИЙ
СТВОЛ
ГОЛОВНОГО МОЗГА
СПИННОЙ МОЗГ
ВТОРИЧНАЯ АФФЕРЕНТАЦИЯ
КОПИЯ
ЭФФЕРЕНТАЦИИ
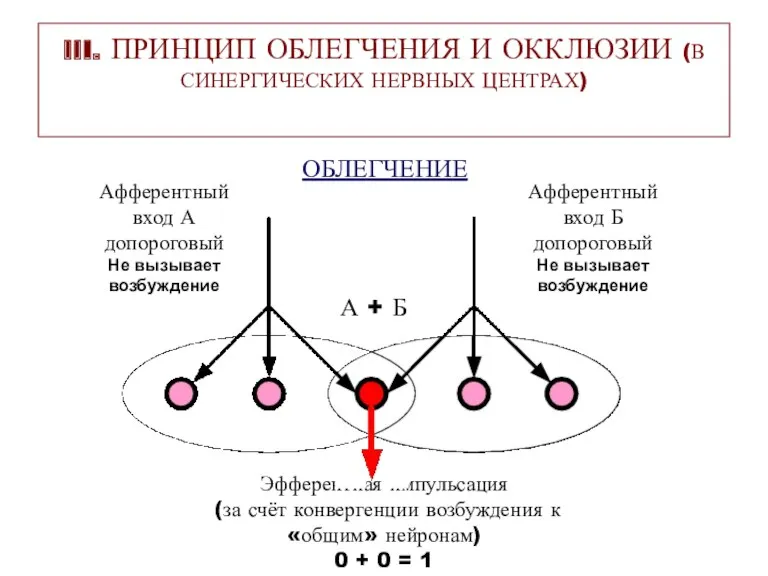
III. ПРИНЦИП ОБЛЕГЧЕНИЯ И ОККЛЮЗИИ (В СИНЕРГИЧЕСКИХ НЕРВНЫХ ЦЕНТРАХ)
ОБЛЕГЧЕНИЕ
Афферентный
вход А
допороговый
Не вызывает
возбуждение
Афферентный
вход
III. ПРИНЦИП ОБЛЕГЧЕНИЯ И ОККЛЮЗИИ (В СИНЕРГИЧЕСКИХ НЕРВНЫХ ЦЕНТРАХ)
ОБЛЕГЧЕНИЕ
Афферентный
вход А
допороговый
Не вызывает
возбуждение
Афферентный
вход
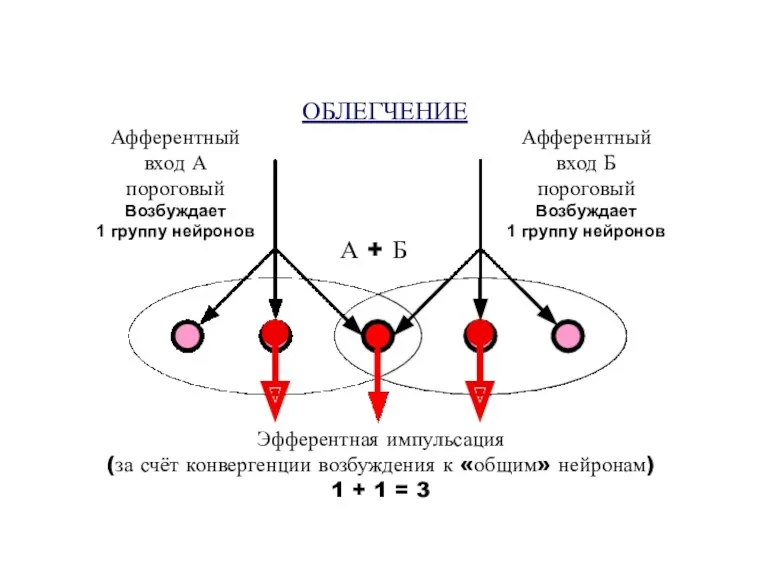
ОБЛЕГЧЕНИЕ
Афферентный
вход А
пороговый
Возбуждает
1 группу нейронов
Афферентный
вход Б
пороговый
Возбуждает
1 группу нейронов
Эфферентная импульсация
(за счёт конвергенции возбуждения
ОБЛЕГЧЕНИЕ
Афферентный
вход А
пороговый
Возбуждает
1 группу нейронов
Афферентный
вход Б
пороговый
Возбуждает
1 группу нейронов
Эфферентная импульсация
(за счёт конвергенции возбуждения
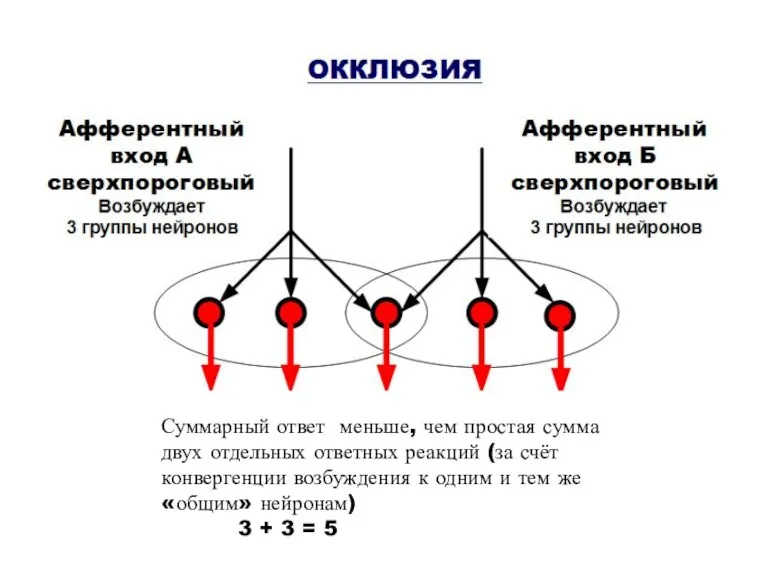
Суммарный ответ меньше, чем простая сумма двух отдельных ответных реакций (за
Суммарный ответ меньше, чем простая сумма двух отдельных ответных реакций (за
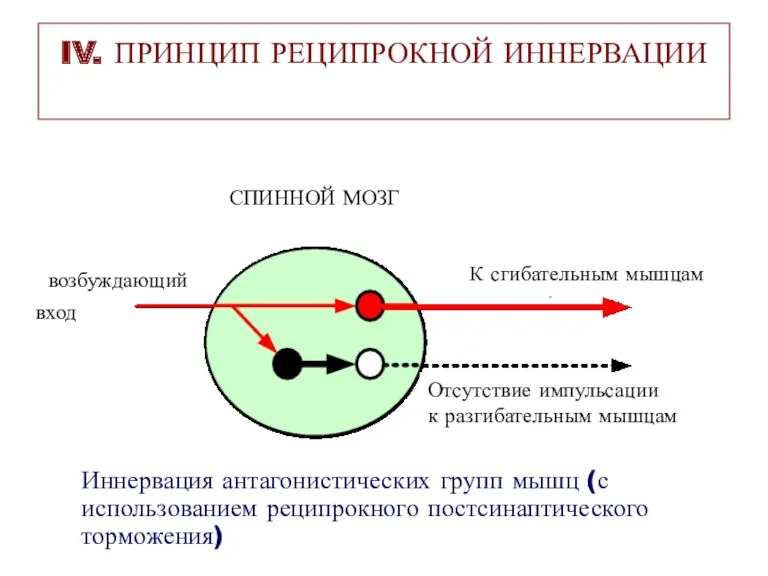
IV. ПРИНЦИП РЕЦИПРОКНОЙ ИННЕРВАЦИИ
СПИННОЙ МОЗГ
вход
возбуждающий
К сгибательным мышцам
Отсутствие импульсации
к разгибательным мышцам
Иннервация
IV. ПРИНЦИП РЕЦИПРОКНОЙ ИННЕРВАЦИИ
СПИННОЙ МОЗГ
вход
возбуждающий
К сгибательным мышцам
Отсутствие импульсации
к разгибательным мышцам
Иннервация
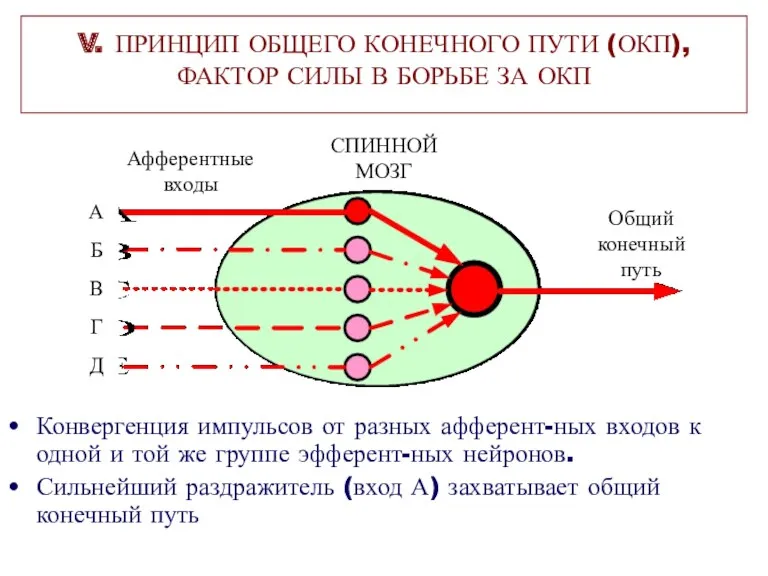
V. ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ (ОКП),
ФАКТОР СИЛЫ В БОРЬБЕ ЗА ОКП
Конвергенция
V. ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ (ОКП),
ФАКТОР СИЛЫ В БОРЬБЕ ЗА ОКП
Конвергенция
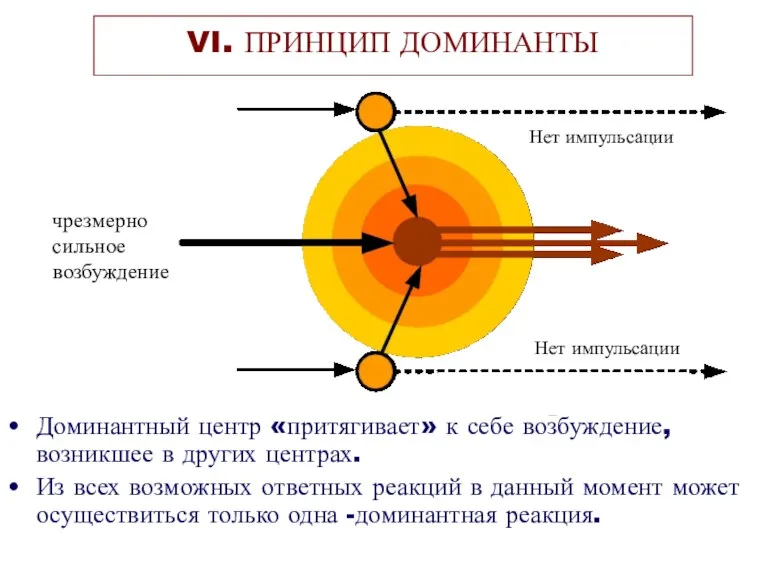
Доминантный центр «притягивает» к себе возбуждение, возникшее в других центрах.
Из всех
Доминантный центр «притягивает» к себе возбуждение, возникшее в других центрах.
Из всех
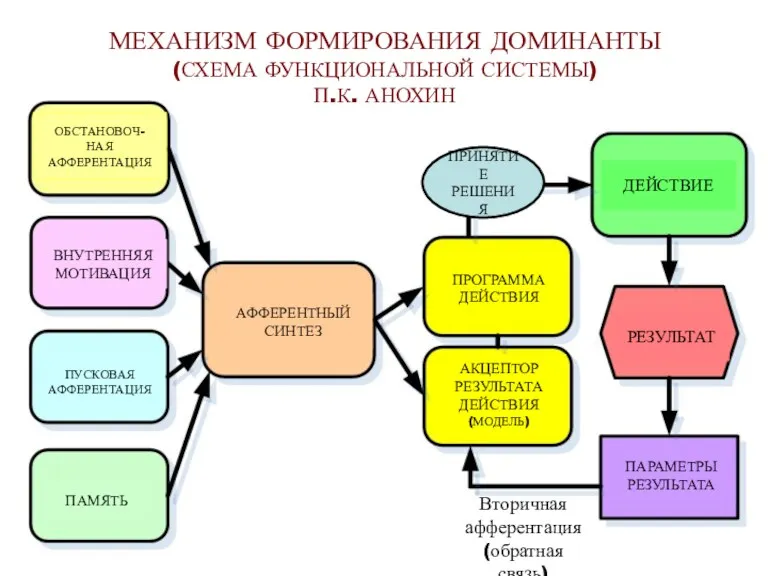
МЕХАНИЗМ ФОРМИРОВАНИЯ ДОМИНАНТЫ
(СХЕМА ФУНКЦИОНАЛЬНОЙ СИСТЕМЫ)
П.К. АНОХИН
ОБСТАНОВОЧ-
НАЯ
АФФЕРЕНТАЦИЯ
ВНУТРЕННЯЯ
МОТИВАЦИЯ
ПУСКОВАЯ
АФФЕРЕНТАЦИЯ
ПАМЯТЬ
АФФЕРЕНТНЫЙ
СИНТЕЗ
ПРОГРАММА
ДЕЙСТВИЯ
АКЦЕПТОР
РЕЗУЛЬТАТА
ДЕЙСТВИЯ
(МОДЕЛЬ)
ДЕЙСТВИЕ
РЕЗУЛЬТАТ
ПАРАМЕТРЫ
РЕЗУЛЬТАТА
Вторичная
афферентация
(обратная связь)
ПРИНЯТИЕ
РЕШЕНИЯ
МЕХАНИЗМ ФОРМИРОВАНИЯ ДОМИНАНТЫ
(СХЕМА ФУНКЦИОНАЛЬНОЙ СИСТЕМЫ)
П.К. АНОХИН
ОБСТАНОВОЧ-
НАЯ
АФФЕРЕНТАЦИЯ
ВНУТРЕННЯЯ
МОТИВАЦИЯ
ПУСКОВАЯ
АФФЕРЕНТАЦИЯ
ПАМЯТЬ
АФФЕРЕНТНЫЙ
СИНТЕЗ
ПРОГРАММА
ДЕЙСТВИЯ
АКЦЕПТОР
РЕЗУЛЬТАТА
ДЕЙСТВИЯ
(МОДЕЛЬ)
ДЕЙСТВИЕ
РЕЗУЛЬТАТ
ПАРАМЕТРЫ
РЕЗУЛЬТАТА
Вторичная
афферентация
(обратная связь)
ПРИНЯТИЕ
РЕШЕНИЯ
 Теоретическая, функциональная и возрастная анатомия костной системы
Теоретическая, функциональная и возрастная анатомия костной системы Общие свойства живого
Общие свойства живого Водоросли. 6 класс
Водоросли. 6 класс Конечный мозг (thelencephalon, cerebrum)
Конечный мозг (thelencephalon, cerebrum) Тайская кошка
Тайская кошка Красная книга
Красная книга Введение в биомеханику. Общая, дифференциальная и частная биомеханика
Введение в биомеханику. Общая, дифференциальная и частная биомеханика Ноосфера
Ноосфера Сон и его значение
Сон и его значение Микрофлора организма человека. Дисбактериоз. Экология микроорганизмов
Микрофлора организма человека. Дисбактериоз. Экология микроорганизмов Основы селекции организмов
Основы селекции организмов Зимующие птицы
Зимующие птицы Биологические ресурсы
Биологические ресурсы Презентация по биологии на тему Абиотические факторы среды. Влажность для 11 класса
Презентация по биологии на тему Абиотические факторы среды. Влажность для 11 класса Авторская дидактическая игра Угадай, кто спрятался?
Авторская дидактическая игра Угадай, кто спрятался? Биологические ритмы
Биологические ритмы Закон гомологических рядов
Закон гомологических рядов Семейство кошачьих
Семейство кошачьих Размножение - свойство живых организмов
Размножение - свойство живых организмов Введение в биоинформатику. Биологические базы данных. Лекция 1
Введение в биоинформатику. Биологические базы данных. Лекция 1 Организация генома. Прокариот и эукариот
Организация генома. Прокариот и эукариот Многолетние растения: флоксы, пионы, розы
Многолетние растения: флоксы, пионы, розы Готовимся к ЕГЭ по биологии
Готовимся к ЕГЭ по биологии Презентация к выступлению на научной конференции Совершенствование естественнонаучного школьного образования
Презентация к выступлению на научной конференции Совершенствование естественнонаучного школьного образования Пептиды. Белки. (Лекция 16)
Пептиды. Белки. (Лекция 16) Классификация царства грибы. Низшие и высшие грибы
Классификация царства грибы. Низшие и высшие грибы Редкие животные нашего округа
Редкие животные нашего округа Интегративная деятельность организма. Рефлексы
Интегративная деятельность организма. Рефлексы