- Обмен белка и аминокислот

Содержание
- 2. Потребность в белках и нормы белкового питания Белки – незаменимый компонент пищи, практически единственный источник азота
- 3. Потребности в белковой пище Коэффициент Рубнера (коэффициент изнашивания) = 53 мг N2 /кг массы тела. 23
- 4. Потребность в белках Физиологический минимум = 35 -50 г в сутки. Оптимум – 85 -100 г
- 5. Внешний обмен белка (переваривание, гидролиз) Поэтапный протеолиз белков до аминокислот, лишение их видоспецифичности и антигенности. Главными
- 7. Роль соляной кислоты 1. Создает кислую среду в полости желудка (рН 1,5 -2), условия для самоактивации
- 8. Регуляция синтеза соляной кислоты Гистидин ? гистамин – активация аденилатциклазы – активация фосфопротеинкиназы с участием цАМФ
- 9. ПЕПСИНОГЕН - ПЕПСИН Пепсин – простой одноцепочечный белок, карбоксильная (в активном центре асп-асп) эндопротеиназа. Активируется в
- 10. Панкреатические протеиназы Синтезируются в виде проферментов и активируются лимитированным протеолизом в просвете 12 – перстной кишки.
- 11. Панкреатические протеиназы Трипсин обеспечивает активацию проэластазы, прокарбоксипептидазы, химотрипсиногена, отщепляя N –концевые пептиды. Каскад протеолитических эндо- и
- 12. Всасывание аминокислот в кишечнике В мембранах энтероцитов кишечных ворсинок – несколько систем активного транспорта (Na+ -зависимый
- 15. Гниение белков в кишечнике Реакции дезаминирования и декарбоксилирования аминокислот с участием бактериальных ферментов. В кишечнике накапливаются
- 16. Обезвреживание продуктов гниения в печени Неспецифические, индуцибельные ферменты микросом печени: ФАФС–трансфераза, УДФ-трансфераза образуют парные, нетоксичные, растворимые
- 17. Внутриклеточный протеолиз Лизосомы. Кислые гидролазы: тиоловые и аспартатные протеиназы (катепсины В, L, H, D), гидролизующие белки.
- 18. Защита от протеолиза Как в клетках, так и во внеклеточном пространстве, в крови работают и системы
- 19. Промежуточный обмен аминокислот. Общие пути катаболизма аминокислот (дезаминирование,трансаминирование, декарбоксилирование) Частные реакции превращений аминокислот. Пути синтеза заменимых
- 20. Метаболические функции аминокислот Кроме участия в синтезе пептидов и белков, у большинства аминокислот активная метаболическая «судьба»:
- 21. СИНТЕЗ ЗАМЕНИМЫХ АМИНОКИСЛОТ Источник углеродных скелетов – глюкоза, азота – NH2 – группы аминокислот, NH3. Реакции
- 22. Дезаминирование аминокислот Механизмы: восстановительный; гидролитический; внутримолекулярный, окислительный. В клетках млекопитающих гис подвергается внутримолекулярному дезаминированию. Сер и
- 24. Окислительное дезаминирование Для каждой аминокислоты есть специфическая оксидаза. FMN –зависимые оксидазы L-аминокислот имеют оптимум рН в
- 25. Окислительное дезаминирование Наиболее активной дезаминазой является глутаматдегидрогеназа (NAD- зависимая) Реакция идет в две стадии: ферментативное окисление
- 27. Трансаминирование Обратимая реакция между кетокислотами и аминокислотами (кофактор – пиридоксальфосфат переносит аминогруппу). На основе кетокислот возникают
- 34. Декарбоксилирование аминокислот При участии пиридоксальзависимых декарбоксилаз образуются биогенные амины. Глу ? γ − аминомасляная кислота Гис
- 38. Конечные продукты белкового обмена Аммиак образуется как результат: дезаминирования аминокислот окисления биогенных аминов утилизация азотистых оснований
- 39. АММИАК В сутки в норме образуется до 20г аммиака, т.е. 4г/л , тем не менее концентрация
- 40. Механизмы токсичности аммиака NH3 проникает через клеточные и митохондриальные мембраны. Увеличение скорости восстановительного аминирования α –кетоглутарата
- 41. Пути обезвреживания аммиака Восстановительное аминирование Образование амидов Синтез азотистых оснований (пиримидинов) Аммониогенез Синтез мочевины «Косвенные» пути,
- 42. Образование амидов дикарбоновых кислот Глутамин- и аспарагин-синтетазы включают аммиак в состав амидов, образуя временную, транспортную нетоксичную
- 43. Восстановительное аминирование кетокислот NADF- зависимая редуктаза восстанавливает кетокислоты до аминокислот. Это путь образования заменимых аминокислот и
- 44. Синтез азотистых оснований (пиримидинов) Синтез пиримидинов начинается с карбамоилсинтетазной реакции: NH3+ CO2+ ATP? NH2COPO32-. Синтез пуринов
- 45. Аммониогенез в почках Глутамин в почках вновь освобождает аммиак (гидролиз глутаминазой) Образовавшася с помощью карбангидразы Н2СО3
- 46. Синтез креатинина Осуществляется при участии ферментов почек и печени из глицина, аргинина и метионина. Креатин фосфорилируется
- 51. Конечные продукты азотистого обмена У организмов разных видов с мочой выделяются разные продукты: Аммонийтелический тип (NH3)
- 52. Орнитиновый цикл синтеза мочевины (цикл Кребса, Ханзеляйта) Гепатоциты, митохондрии, аэробные условия. АТР, СО2, орнитин, цитруллин, аспартат,
- 61. Конечные продукты азотистого обмена Фракции «остаточного» азота в крови: Мочевина (50% N2 крови и 90% N2
- 63. Скачать презентацию

Потребность в белках и нормы белкового питания
Белки – незаменимый компонент пищи,
Потребность в белках и нормы белкового питания
Белки – незаменимый компонент пищи,

Потребности в белковой пище
Коэффициент Рубнера (коэффициент изнашивания) = 53 мг N2
Потребности в белковой пище
Коэффициент Рубнера (коэффициент изнашивания) = 53 мг N2

Потребность в белках
Физиологический минимум = 35 -50 г в сутки.
Оптимум –
Потребность в белках
Физиологический минимум = 35 -50 г в сутки.
Оптимум –

Внешний обмен белка (переваривание, гидролиз)
Поэтапный протеолиз белков до аминокислот, лишение их
Внешний обмен белка (переваривание, гидролиз)
Поэтапный протеолиз белков до аминокислот, лишение их
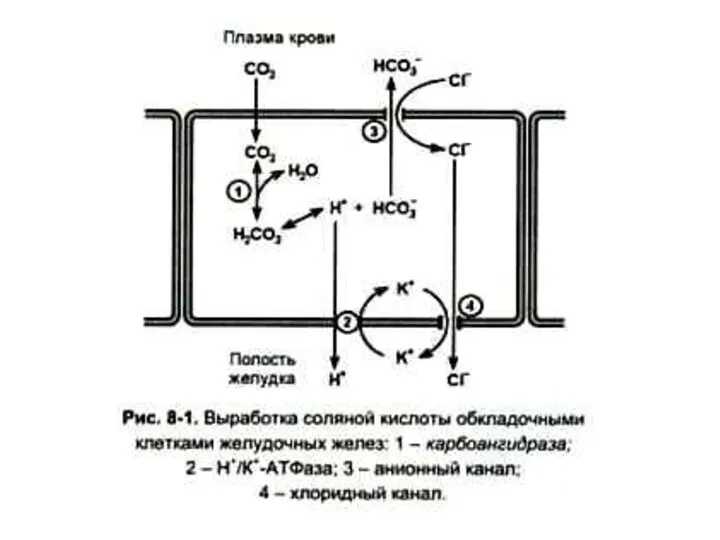

Роль соляной кислоты
1. Создает кислую среду в полости желудка (рН 1,5
Роль соляной кислоты
1. Создает кислую среду в полости желудка (рН 1,5

Регуляция синтеза соляной кислоты
Гистидин ? гистамин – активация аденилатциклазы – активация
Регуляция синтеза соляной кислоты
Гистидин ? гистамин – активация аденилатциклазы – активация

ПЕПСИНОГЕН - ПЕПСИН
Пепсин – простой одноцепочечный белок, карбоксильная (в активном центре
ПЕПСИНОГЕН - ПЕПСИН
Пепсин – простой одноцепочечный белок, карбоксильная (в активном центре

Панкреатические протеиназы
Синтезируются в виде проферментов и активируются лимитированным протеолизом в просвете
Панкреатические протеиназы
Синтезируются в виде проферментов и активируются лимитированным протеолизом в просвете

Панкреатические протеиназы
Трипсин обеспечивает активацию проэластазы, прокарбоксипептидазы, химотрипсиногена, отщепляя N –концевые пептиды.
Каскад
Панкреатические протеиназы
Трипсин обеспечивает активацию проэластазы, прокарбоксипептидазы, химотрипсиногена, отщепляя N –концевые пептиды.
Каскад

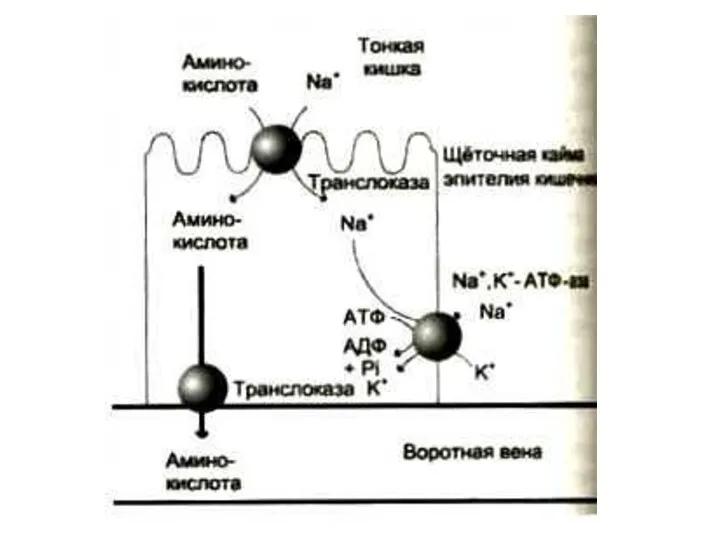
Всасывание аминокислот в кишечнике
В мембранах энтероцитов кишечных ворсинок – несколько
Всасывание аминокислот в кишечнике
В мембранах энтероцитов кишечных ворсинок – несколько
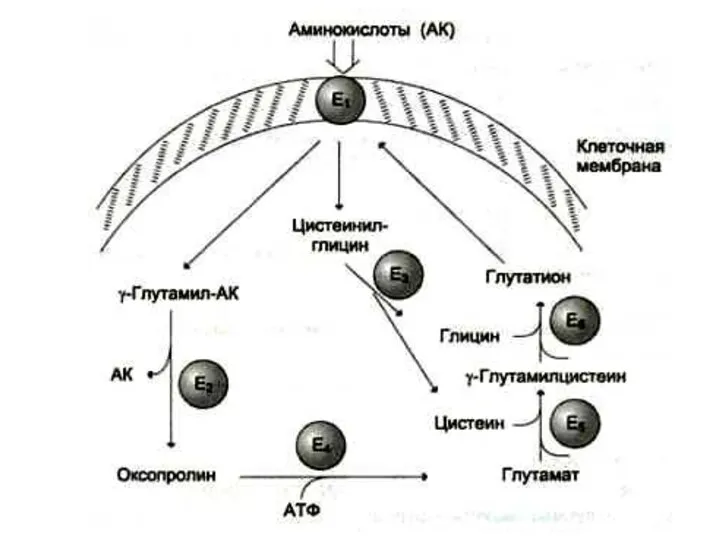

Гниение белков в кишечнике
Реакции дезаминирования и декарбоксилирования аминокислот с участием бактериальных
Гниение белков в кишечнике
Реакции дезаминирования и декарбоксилирования аминокислот с участием бактериальных

Обезвреживание продуктов гниения в печени
Неспецифические, индуцибельные ферменты микросом печени:
ФАФС–трансфераза, УДФ-трансфераза
Обезвреживание продуктов гниения в печени
Неспецифические, индуцибельные ферменты микросом печени:
ФАФС–трансфераза, УДФ-трансфераза

Внутриклеточный протеолиз
Лизосомы. Кислые гидролазы: тиоловые и аспартатные протеиназы (катепсины В, L,
Внутриклеточный протеолиз
Лизосомы. Кислые гидролазы: тиоловые и аспартатные протеиназы (катепсины В, L,

Защита от протеолиза
Как в клетках, так и во внеклеточном пространстве, в
Защита от протеолиза
Как в клетках, так и во внеклеточном пространстве, в

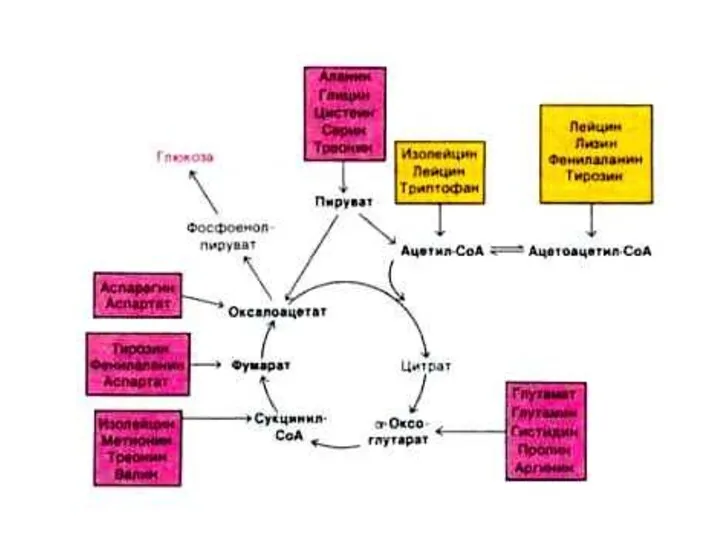
Промежуточный обмен аминокислот.
Общие пути катаболизма аминокислот (дезаминирование,трансаминирование, декарбоксилирование)
Частные реакции превращений
Промежуточный обмен аминокислот.
Общие пути катаболизма аминокислот (дезаминирование,трансаминирование, декарбоксилирование)
Частные реакции превращений

Метаболические функции аминокислот
Кроме участия в синтезе пептидов и белков, у большинства
Метаболические функции аминокислот
Кроме участия в синтезе пептидов и белков, у большинства

СИНТЕЗ ЗАМЕНИМЫХ АМИНОКИСЛОТ
Источник углеродных скелетов – глюкоза, азота – NH2 –
СИНТЕЗ ЗАМЕНИМЫХ АМИНОКИСЛОТ
Источник углеродных скелетов – глюкоза, азота – NH2 –

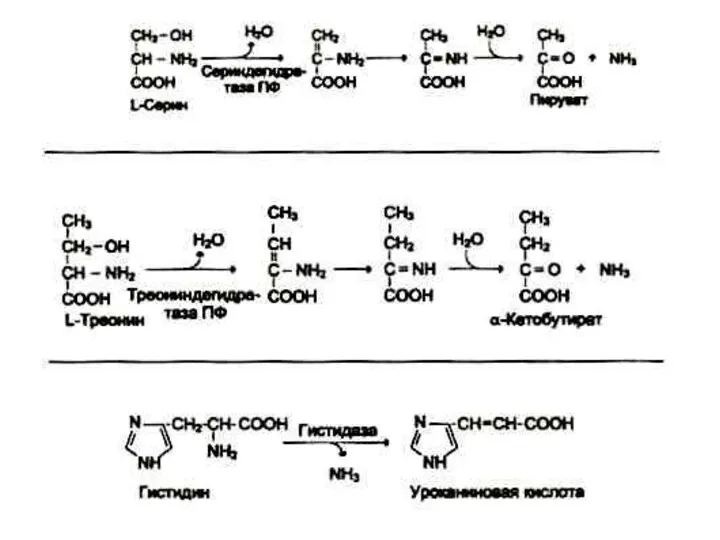
Дезаминирование аминокислот
Механизмы: восстановительный; гидролитический; внутримолекулярный, окислительный.
В клетках млекопитающих гис подвергается внутримолекулярному
Дезаминирование аминокислот
Механизмы: восстановительный; гидролитический; внутримолекулярный, окислительный.
В клетках млекопитающих гис подвергается внутримолекулярному

Окислительное дезаминирование
Для каждой аминокислоты есть специфическая оксидаза.
FMN –зависимые оксидазы L-аминокислот имеют
Окислительное дезаминирование
Для каждой аминокислоты есть специфическая оксидаза.
FMN –зависимые оксидазы L-аминокислот имеют

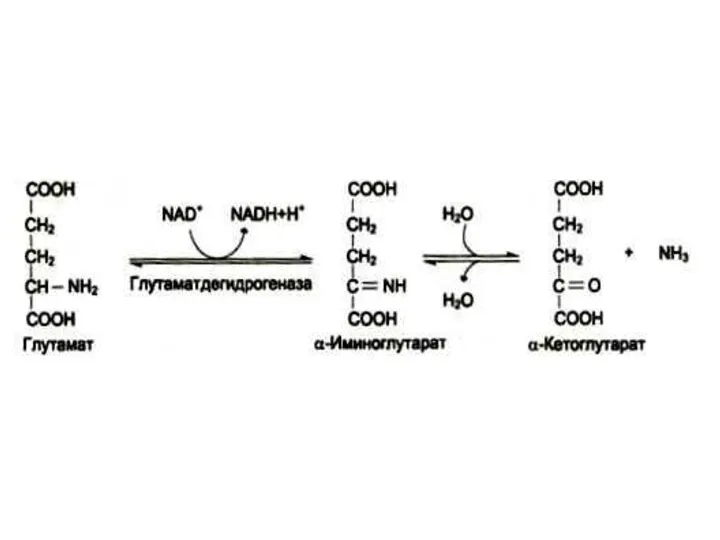
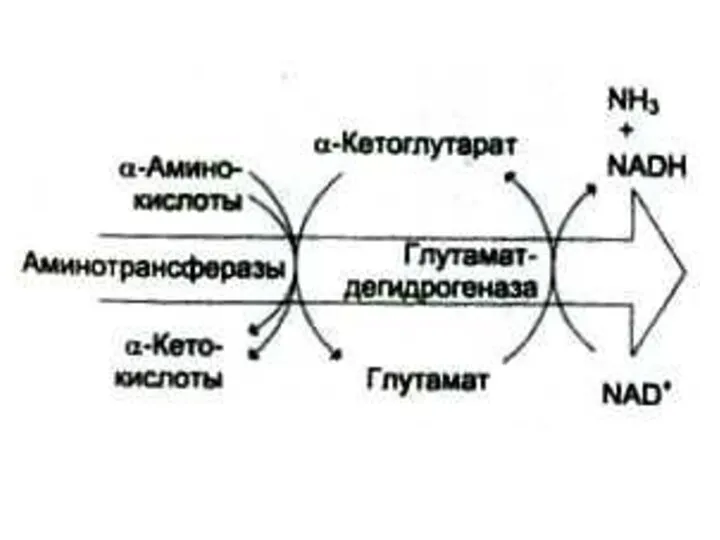
Окислительное дезаминирование
Наиболее активной дезаминазой является глутаматдегидрогеназа (NAD- зависимая)
Реакция идет в две
Окислительное дезаминирование
Наиболее активной дезаминазой является глутаматдегидрогеназа (NAD- зависимая)
Реакция идет в две

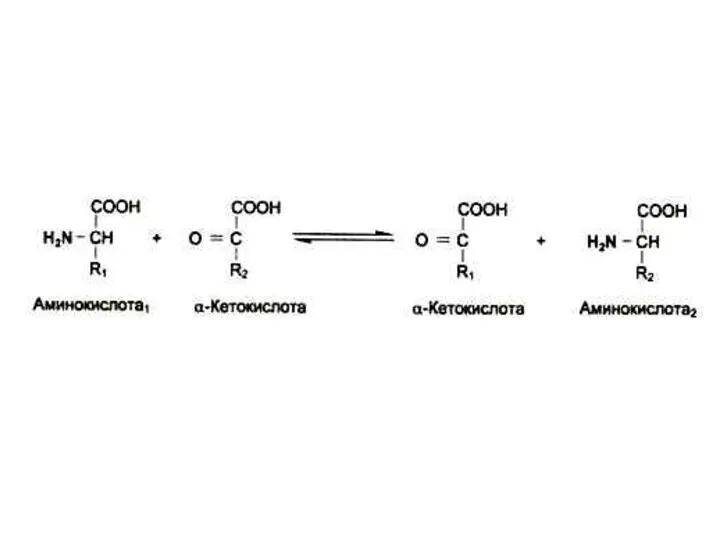
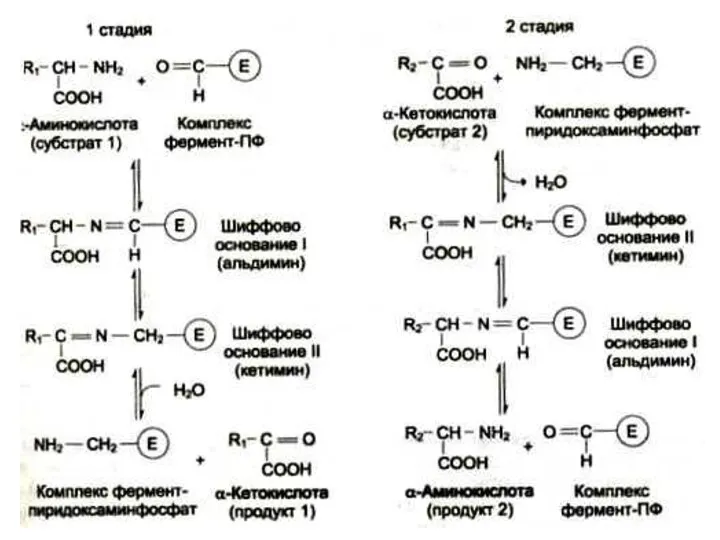
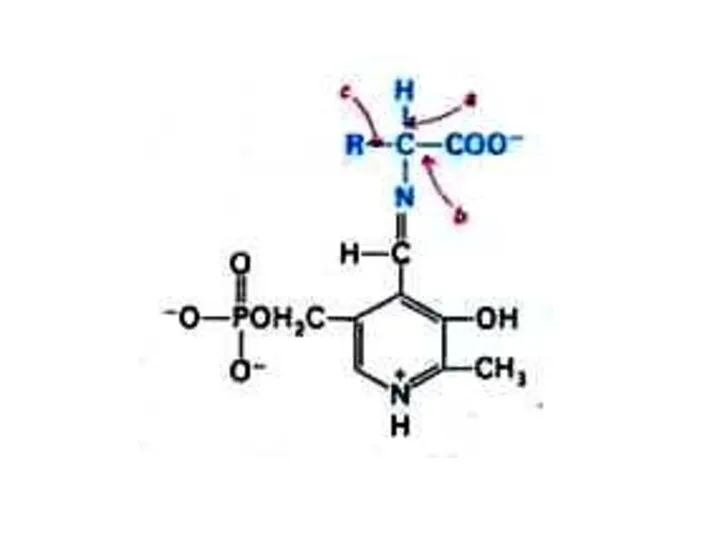
Трансаминирование
Обратимая реакция между кетокислотами и аминокислотами (кофактор – пиридоксальфосфат переносит
Трансаминирование
Обратимая реакция между кетокислотами и аминокислотами (кофактор – пиридоксальфосфат переносит
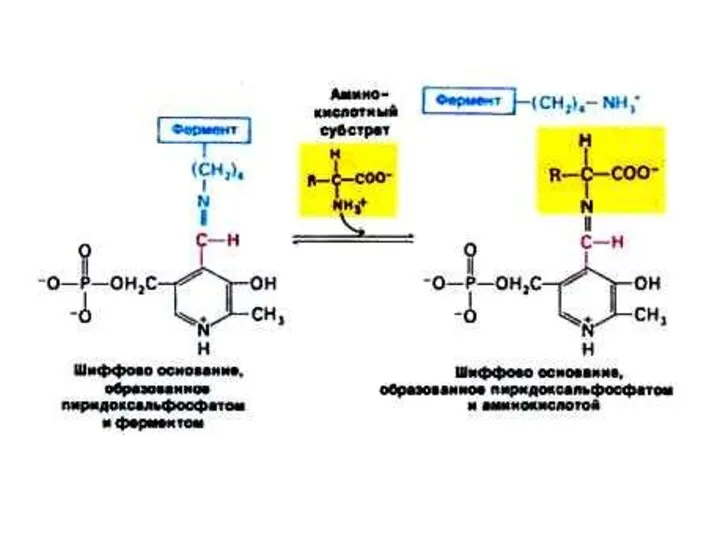
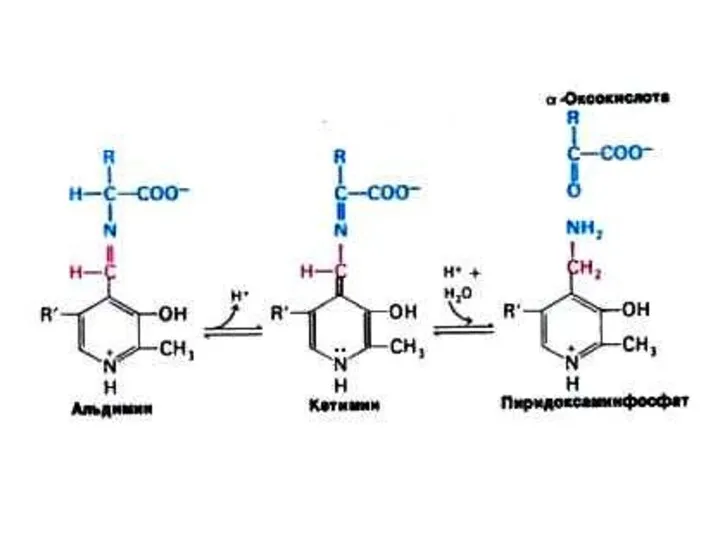

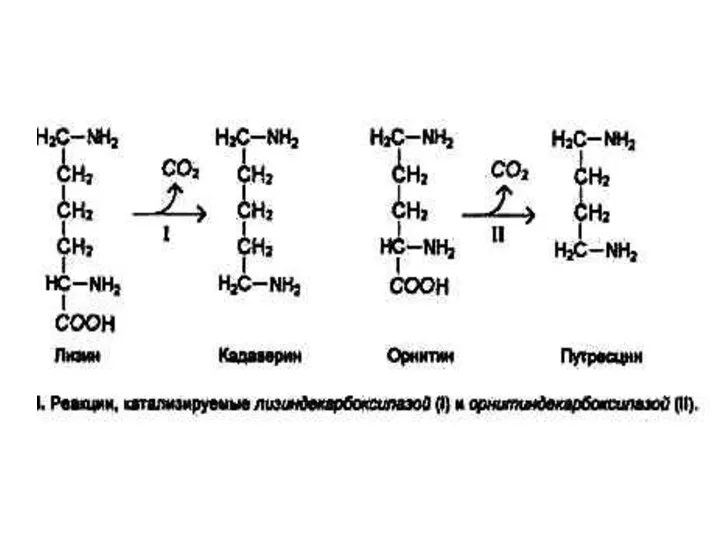
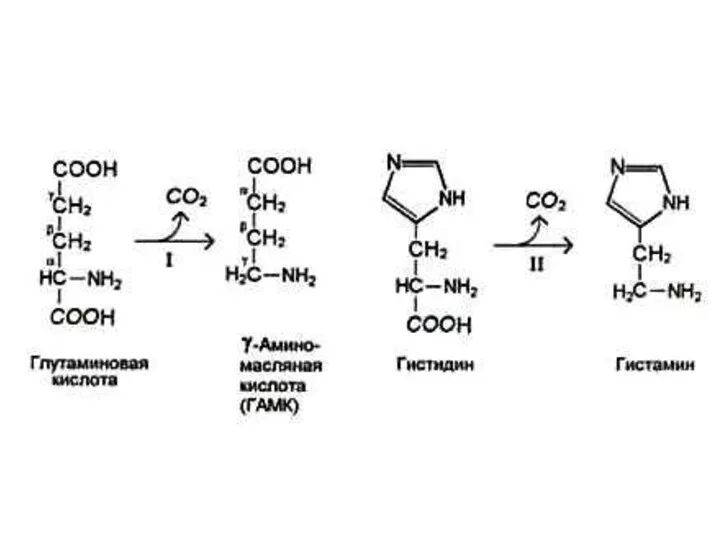
Декарбоксилирование аминокислот
При участии пиридоксальзависимых декарбоксилаз образуются биогенные амины.
Глу ? γ −
Декарбоксилирование аминокислот
При участии пиридоксальзависимых декарбоксилаз образуются биогенные амины.
Глу ? γ −

Конечные продукты белкового обмена
Аммиак образуется как результат:
дезаминирования аминокислот
окисления биогенных аминов
Конечные продукты белкового обмена
Аммиак образуется как результат:
дезаминирования аминокислот
окисления биогенных аминов

АММИАК
В сутки в норме образуется до 20г аммиака, т.е. 4г/л ,
АММИАК
В сутки в норме образуется до 20г аммиака, т.е. 4г/л ,

Механизмы токсичности аммиака
NH3 проникает через клеточные и митохондриальные мембраны.
Увеличение скорости восстановительного
Механизмы токсичности аммиака
NH3 проникает через клеточные и митохондриальные мембраны.
Увеличение скорости восстановительного

Пути обезвреживания аммиака
Восстановительное аминирование
Образование амидов
Синтез азотистых оснований (пиримидинов)
Аммониогенез
Синтез мочевины
«Косвенные» пути, реакции
Пути обезвреживания аммиака
Восстановительное аминирование
Образование амидов
Синтез азотистых оснований (пиримидинов)
Аммониогенез
Синтез мочевины
«Косвенные» пути, реакции

Образование амидов дикарбоновых кислот
Глутамин- и аспарагин-синтетазы включают аммиак в состав амидов,
Образование амидов дикарбоновых кислот
Глутамин- и аспарагин-синтетазы включают аммиак в состав амидов,

Восстановительное аминирование кетокислот
NADF- зависимая редуктаза восстанавливает кетокислоты до аминокислот.
Это путь образования
Восстановительное аминирование кетокислот
NADF- зависимая редуктаза восстанавливает кетокислоты до аминокислот.
Это путь образования

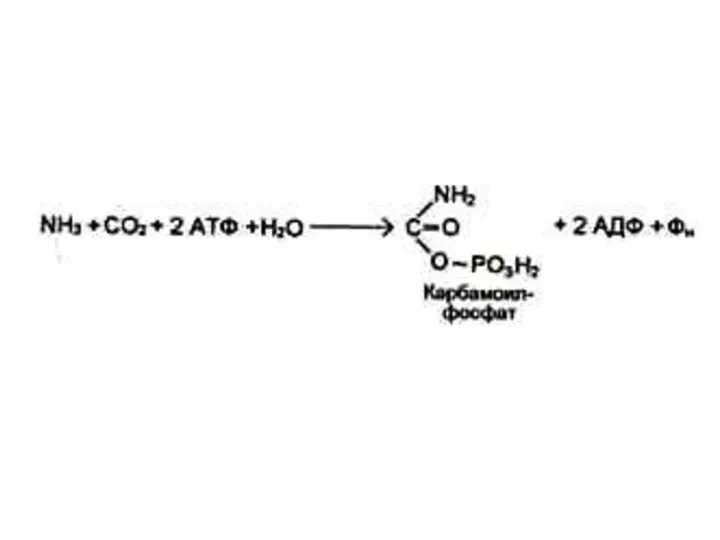
Синтез азотистых оснований (пиримидинов)
Синтез пиримидинов начинается с карбамоилсинтетазной реакции:
NH3+ CO2+ ATP?
Синтез азотистых оснований (пиримидинов)
Синтез пиримидинов начинается с карбамоилсинтетазной реакции:
NH3+ CO2+ ATP?

Аммониогенез в почках
Глутамин в почках вновь освобождает аммиак (гидролиз глутаминазой)
Образовавшася с
Аммониогенез в почках
Глутамин в почках вновь освобождает аммиак (гидролиз глутаминазой)
Образовавшася с

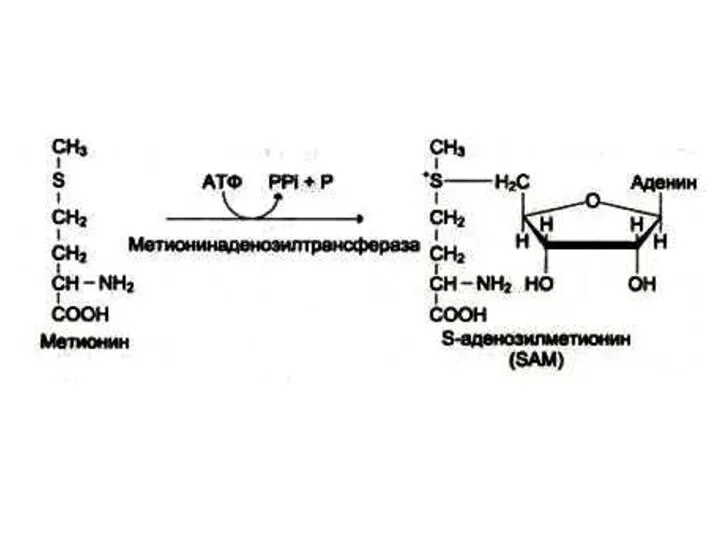
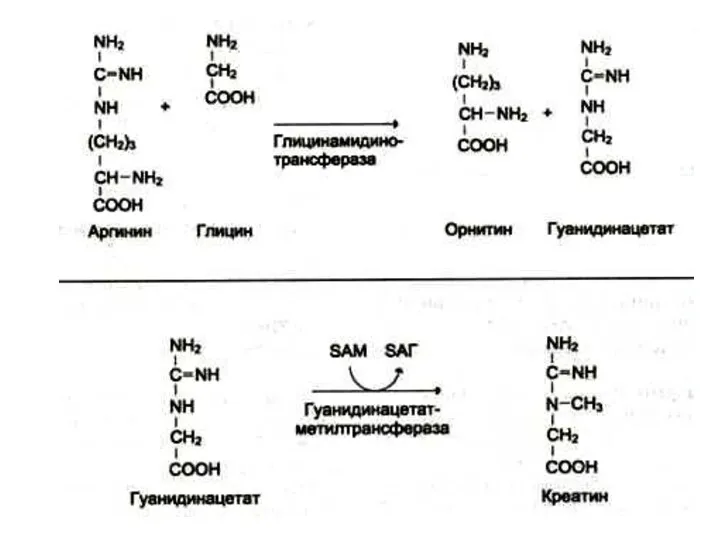
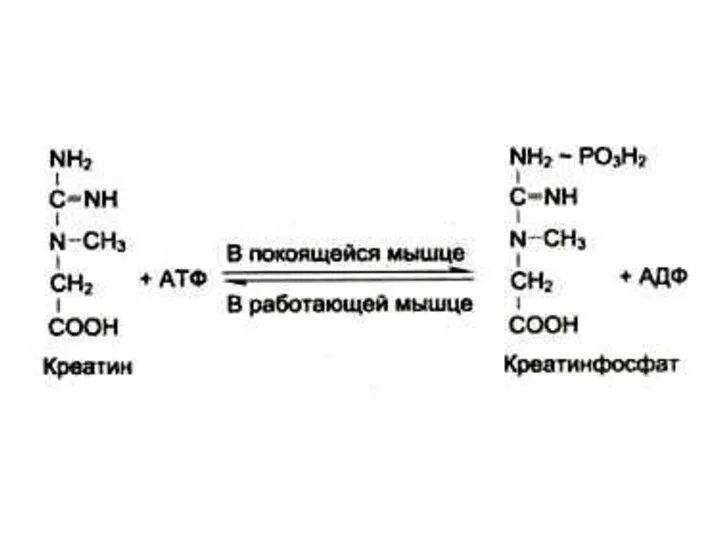
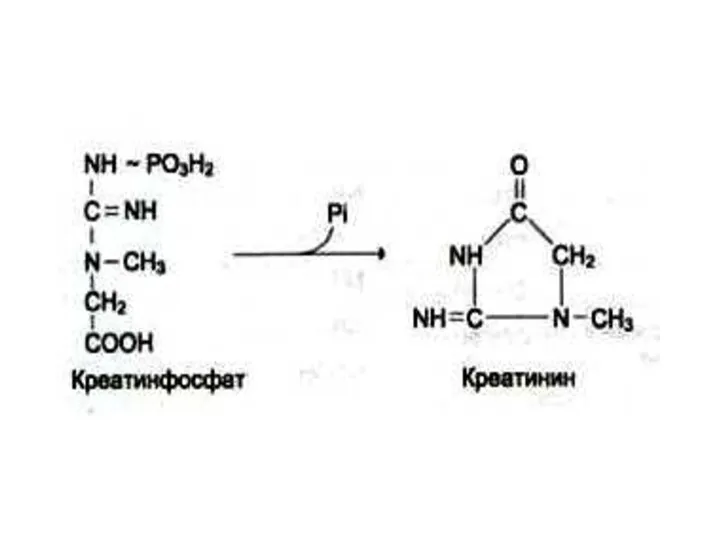
Синтез креатинина
Осуществляется при участии ферментов почек и печени из глицина, аргинина
Синтез креатинина
Осуществляется при участии ферментов почек и печени из глицина, аргинина

Конечные продукты азотистого обмена
У организмов разных видов с мочой выделяются разные
Конечные продукты азотистого обмена
У организмов разных видов с мочой выделяются разные

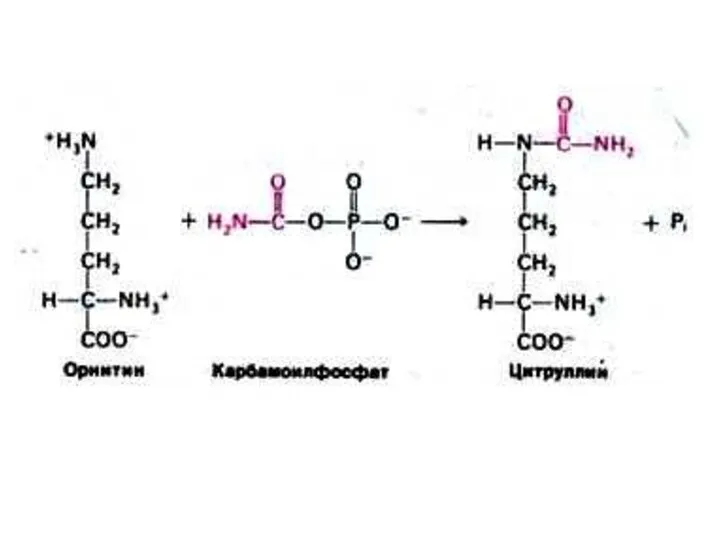
Орнитиновый цикл синтеза мочевины (цикл Кребса, Ханзеляйта)
Гепатоциты, митохондрии, аэробные условия.
АТР, СО2,
Орнитиновый цикл синтеза мочевины (цикл Кребса, Ханзеляйта)
Гепатоциты, митохондрии, аэробные условия.
АТР, СО2,
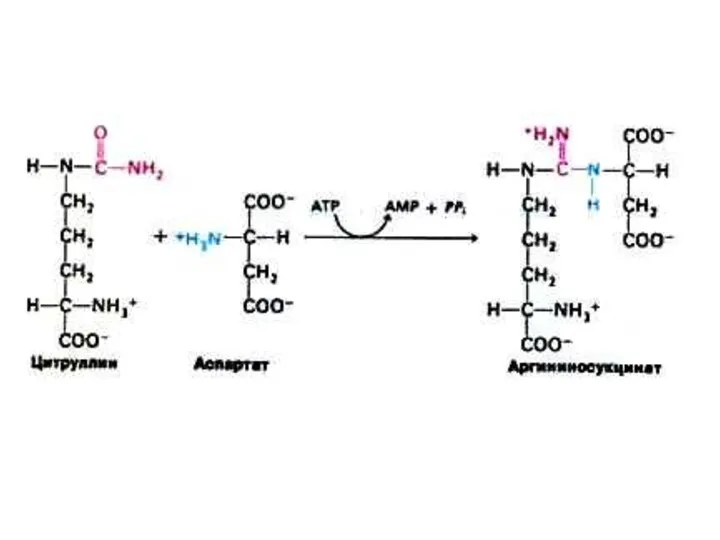
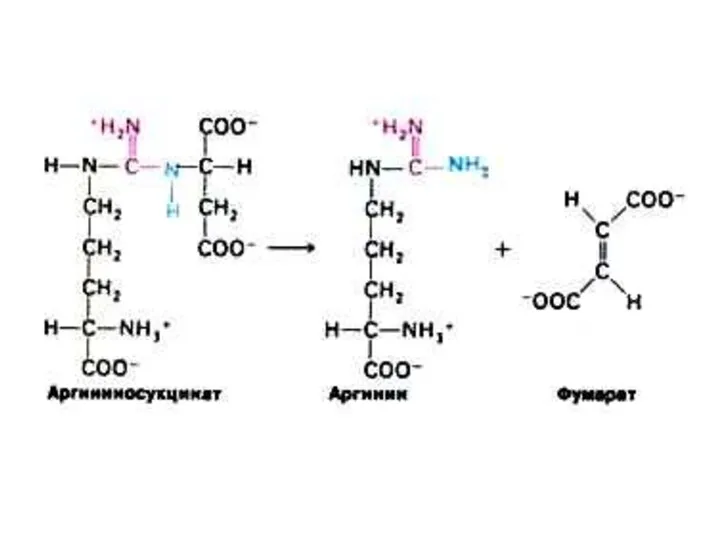
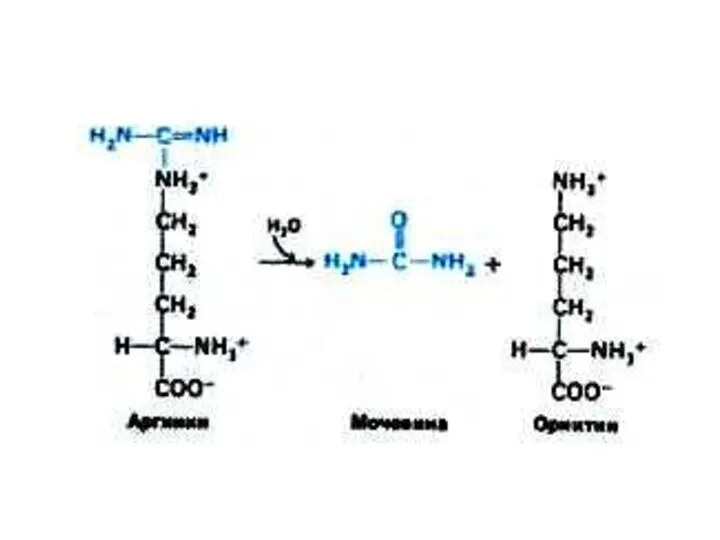
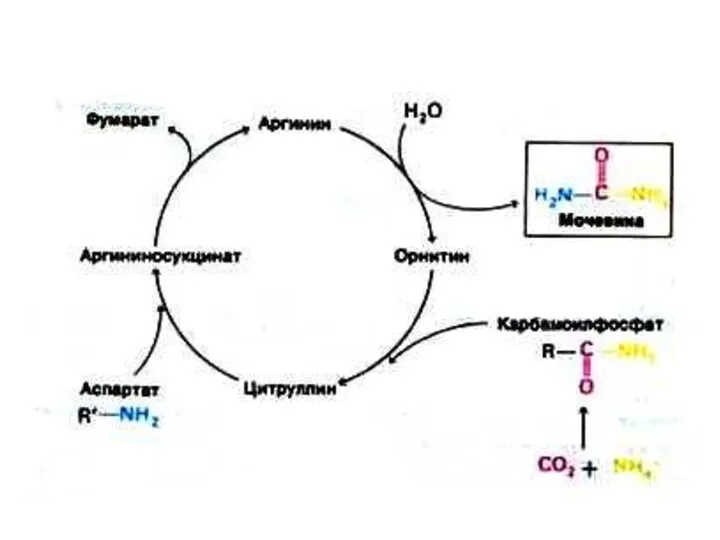
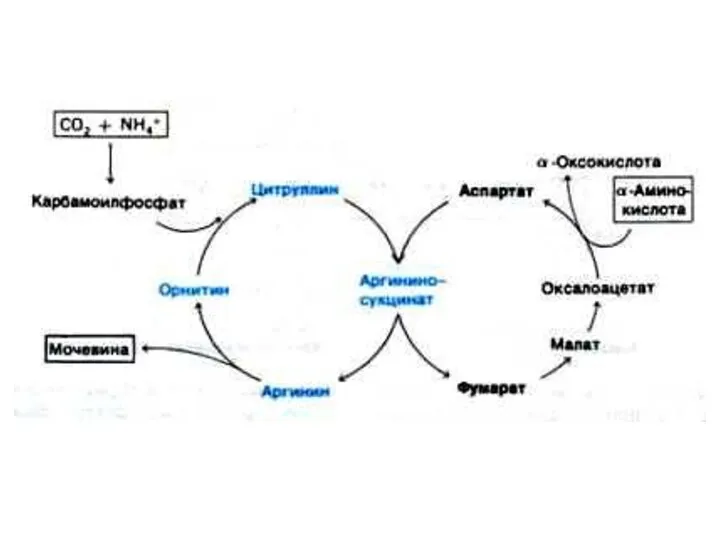
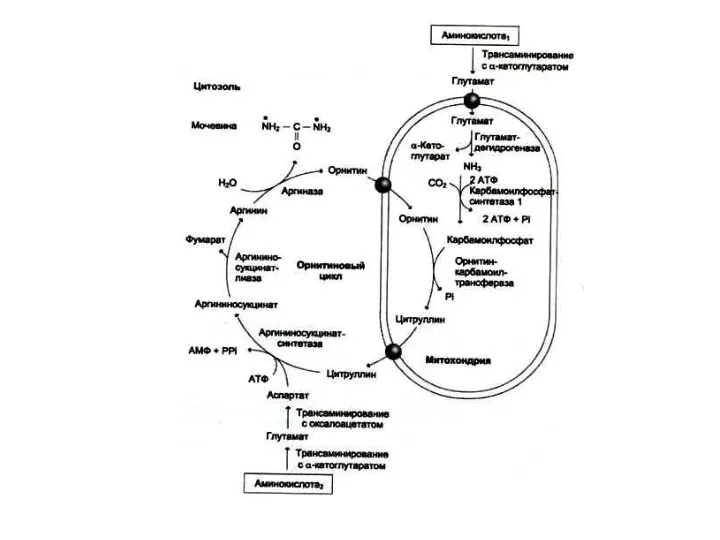

Конечные продукты азотистого обмена
Фракции «остаточного» азота в крови:
Мочевина (50% N2 крови
Конечные продукты азотистого обмена
Фракции «остаточного» азота в крови:
Мочевина (50% N2 крови
 Подготовка обучающихся 9-х классов к ОГЭ по биологии 2018 года
Подготовка обучающихся 9-х классов к ОГЭ по биологии 2018 года Микробиология как наука, предмет ее изучения, исторические этапы развития, задачи медицинской микробиологии
Микробиология как наука, предмет ее изучения, исторические этапы развития, задачи медицинской микробиологии Віруси людини
Віруси людини Vitamins. Classes of Vitamins
Vitamins. Classes of Vitamins Понятие о гене. Биосинтез белка. Транскрипция
Понятие о гене. Биосинтез белка. Транскрипция Модификационная изменчивость
Модификационная изменчивость Мышление и деятельность
Мышление и деятельность Эволюция приматов
Эволюция приматов Отчет- презентация О подготовке учащихся к сдаче ЕНТ
Отчет- презентация О подготовке учащихся к сдаче ЕНТ Взаимосвязи компонентов природы. Природный комплекс. (проект Урок в Москве)
Взаимосвязи компонентов природы. Природный комплекс. (проект Урок в Москве) Кошки
Кошки Зачем мы едим овощи и фрукты
Зачем мы едим овощи и фрукты Творческий проект по технологии. Комнатное растение в интерьере гостиной
Творческий проект по технологии. Комнатное растение в интерьере гостиной Открытый урок Внешнее строение Насекомых
Открытый урок Внешнее строение Насекомых Вопросы к викторине по охране природы
Вопросы к викторине по охране природы Листопад. Причини опадання листя
Листопад. Причини опадання листя Дикорослі та культурні рослини
Дикорослі та культурні рослини The endangered animals
The endangered animals Молекулярная биология. (Лекция 5-6)
Молекулярная биология. (Лекция 5-6) Использование фитонцидоносных растений для очистки воздуха
Использование фитонцидоносных растений для очистки воздуха Репродуктивная система человека
Репродуктивная система человека Углеводы и их роль и значение в жизни человека
Углеводы и их роль и значение в жизни человека iPavlov: Conversational Intelligence Project
iPavlov: Conversational Intelligence Project Основные понятия генетики
Основные понятия генетики Живые изгороди
Живые изгороди Класс Млекопитающие. Mammalia. Лекция14
Класс Млекопитающие. Mammalia. Лекция14 Пищеварительная система человека
Пищеварительная система человека Хищные животные Хабаровского края
Хищные животные Хабаровского края