- Обмен белков - 2

Содержание
- 2. Катаболизм аминокислот
- 3. Метионин
- 4. Обмен метионина
- 5. S-аденозилметионин (SAМ)
- 6. Трансметилирование – реакции переноса метильной группы от донора к акцептору А-СН3 + В А + В-СН3
- 7. Использование СН3-радикалов
- 8. Синтез креатина Креатин поступает в мышцы, перефосфорилируется с АТФ и в виде креатин-фосфата является резервной формой
- 9. Синтез адреналина
- 10. Синтез холина Холин входит в состав: фосфатидилхолина – компонента клеточных мембран ацетилхолина – нейромедиатора
- 11. Синтез тимина Тимин – азотистое основание, входящее в состав ДНК метилтранс-фераза
- 12. Инактивация гистамина метилтранс-фераза
- 13. Обезвреживание никотинамида метилтранс-фераза
- 14. Обезвреживание ксенобиотиков
- 15. Тетрагидрофолиевая кислота (ТГФК - Н4-фолат)
- 16. Одноуглеродные радикалы
- 17. Доноры одноуглеродных групп
- 18. Антивитамины фолиевой кислоты
- 19. Обмен ароматических аминокислот
- 20. Синтез тирозина
- 21. Нарушения обмена фенилаланина
- 22. Скрининг-тест – это исследование, которое: Проводится для 100% популяции (н., для всех новорожденных), Это доклиническое исследование,
- 23. Синтез катехоламинов
- 24. Йодтиронины Йодтиронины – гормоны щитовидной железы, участвующие в общем обмене, регулирующие нервную деятельность, рост и дифференцировку
- 25. Синтез меланина Меланин – пигмент, входящий в состав волос, кожи, радужной оболочки глаза. Защищает организм от
- 26. Распад тирозина
- 27. Нарушения обмена ароматических аминокислот
- 28. Алкаптонурия
- 29. Альбинизм
- 30. Конечные продукты обмена белков
- 31. Основные источники аммиака
- 32. Обезвреживание аммиака в месте образования 1. Восстановительное аминирование α-кетоглутарата α-кетоглутарат глутамат
- 33. Обезвреживание аммиака в месте образования 2. Амидирование аминокислот (синтез амидов)
- 34. 3. Амидирование карбоксильных групп белков Обезвреживание аммиака в месте образования
- 35. 4. Синтез карбамоилфосфата Обезвреживание аммиака в месте образования
- 36. Транспортные формы аммиака Глутаминовая кислота Амиды аминокислот (аспарагин и глутамин) Амидированные белки
- 37. Роль глутамина
- 38. Синтез аммонийных солей (аммониегенез)
- 39. Биологическая роль аммониегенеза Обезвреживание и выведение токсического аммиака Поддержание кислотно-основного состояния (КОС) за счёт выведения избыточных
- 40. Орнитиновый цикл (цикл мочевинообразования)
- 42. Особенности цикла мочевинообразования На синтез одной молекулы мочевины затрачивается 3 молекулы АТФ (но 4 макро-эргических связи)
- 43. Биологическая роль орнитинового цикла Конечное обезвреживание токсического аммиака Синтез аргинина и пополнение его фонда в организме
- 44. Связь цикла мочевинообразования и ЦТК Оба цикла протекают в одном и том же месте – в
- 45. АМФ + РРi
- 46. Нарушения синтеза мочевины аргиназа
- 47. Нарушение синтеза и выведения мочевины
- 48. Соотношение азотсодержащих веществ в моче (%) при нормальном белковом питании
- 49. КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ КАФЕДРА ФУНДАМЕНТАЛЬНОЙ И КЛИНИЧЕСКОЙ БИОХИМИИ Лекция по биохимии Тема: «Обмен белков–3» Краснодар
- 50. Строение мононуклеотида азотистое основание пентоза остаток фосфорной кислоты нуклеозид НУКЛЕОТИД
- 51. Пуриновые основания нуклеиновых кислот
- 52. Пиримидиновые основания нуклеиновых кислот
- 53. Углеводы нуклеотидов (пентозы) (РНК) (ДНК) рибоза дезоксирибоза
- 54. Гидролиз нуклеопротеинов
- 55. Катаболизм пуриновых оснований В норме содержание мочевой кислоты в крови 0,14-0,5 ммоль/л
- 56. Нарушения обмена пуринов
- 57. Ингибитор ксантиноксидазы
- 58. Распад пиримидиновых оснований
- 59. Особенности синтеза нуклеотидов Синтез идёт из обычных простых предшественников (ак, углекислого газа и т.п.) Синтезируются не
- 60. (фосфорибозиопирофосфат) Синтез пуриновых нуклеотидов
- 61. Происхождение атомов пуринового кольца СН≡ТГФК
- 62. Синтез пуриновых нуклеотидов
- 63. Синтез пиримидиновых нуклеотидов глу-NH2 оротидинмонофосфат (ОМФ) уридинмонофосфат (УМФ)
- 64. Синтез пиримидиновых нуклеотидов
- 65. Синтез дезоксирибонуклеотидов
- 66. Ингибиторы синтеза нуклеиновых кислот
- 67. Нарушения обмена нуклеотидов Нарушение синтеза пиримидинов – оротацидурия – дефект ОМФ-декарбоксилазы (недостаток синтеза пиримидиновых нуклеотидов, снижение
- 68. основание фосфодиэфирная связь Первичная структура нуклеиновых кислот основание основание
- 69. Вторичная структура ДНК
- 70. Вторичная структура р-РНК
- 71. Вторичная структура т-РНК
- 72. Строение нуклеосомы
- 73. Клеточный цикл
- 74. Строение мононуклеотида
- 75. Направление роста цепи 5'→3' Поступающий нуклеотид он х х х Растущая цепь НК Матрица – цепь
- 76. Биосинтез ДНК (репликация) является: матричным (матрица – обе нити ДНК) комплиментарным фрагментарным (нити ДНК синтезируются в
- 77. Полуконсервативность биосинтеза ДНК
- 79. Особенности репликации ДНК-полимеразы δ и ε не могут соединять между собой два мононуклеотида, а только достраивают
- 80. Этапы репликации 1. Инициация: Топоизомераза находит точку начала репликации, гидролизует одну фосфодиэфирную связь и даёт возможность
- 81. Этапы репликации ДНК-связывающие белки (SSB-белки) стабилизируют репликативную вилку, не давая восстанавливаться водородным связям между комплиментарными нуклеотидами
- 82. Инициация репликации ДНК-полимераза α ДНК-полимераза δ ДНК-полимераза ε
- 83. Ориджин репликации репликон Инициация репликации
- 84. 2. Элонгация ДНК-полимераза δ продолжает удлинять нить из дезоксирибонуклеотидов на лидирующей нити, а ДНК-полимераза ε –
- 85. Элонгация репликации
- 86. Терминация ДНК-полимераза β (фермент репарации) удаляет праймеры и достраивает фрагменты ДНК ДНК-лигаза соединяет фрагменты между собой
- 87. Функции ДНК-полимеразы β
- 88. Репарация ДНК
- 89. Для биосинтеза РНК (транскрипции) необходимы: МАТРИЦА – участок одной из нитей ДНК – (транскриптон) СТРОИТЕЛЬНЫЙ МАТЕРИАЛ:
- 90. Биосинтез РНК
- 91. Транскрибируемая нить Нетранскрибируемая нить РНК-полимераза ДНК пре-РНК (первичный транскрипт) Биосинтез РНК мононуклеотиды Сайт терминации
- 92. Процессинг РНК (1. сплайсинг)
- 93. м-РНК 7-метилгуанозин полиаденилат 5' 3' Процессинг (2. модификация концов м-РНК)
- 95. Скачать презентацию
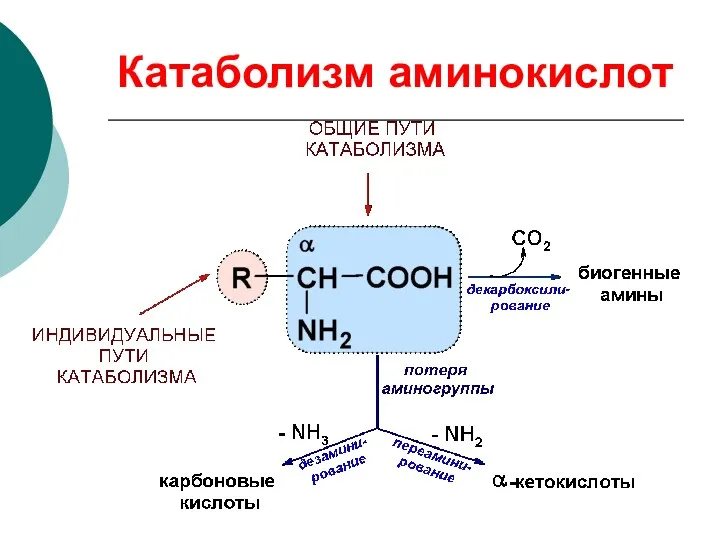
Катаболизм аминокислот
Катаболизм аминокислот
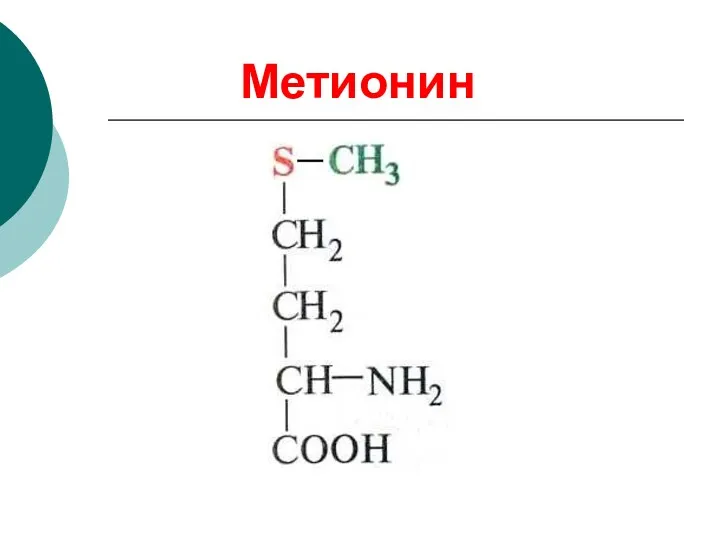
Метионин
Метионин
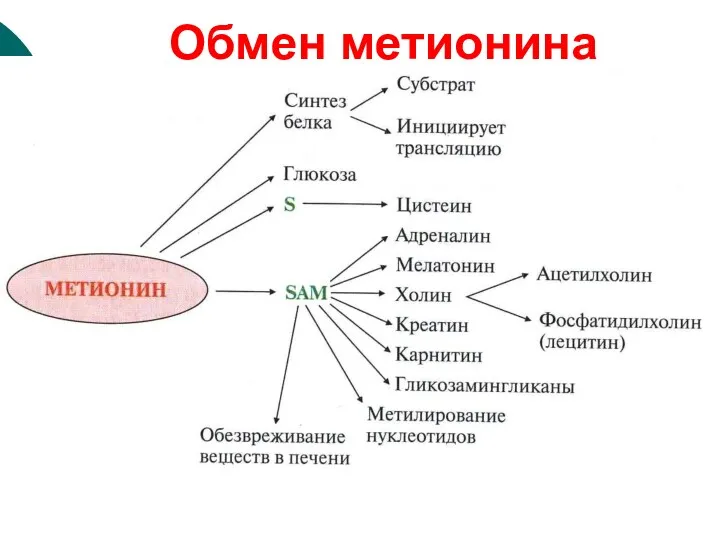
Обмен метионина
Обмен метионина
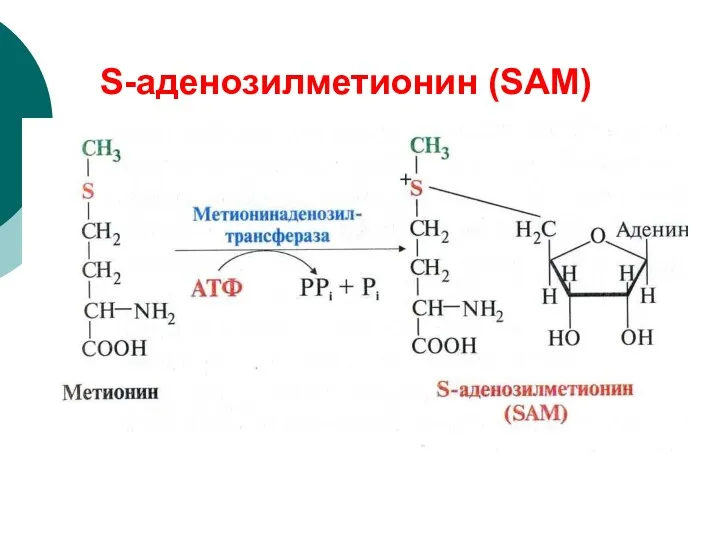
S-аденозилметионин (SAМ)
S-аденозилметионин (SAМ)

Трансметилирование – реакции переноса метильной группы от донора к акцептору
А-СН3 +
Трансметилирование – реакции переноса метильной группы от донора к акцептору
А-СН3 +
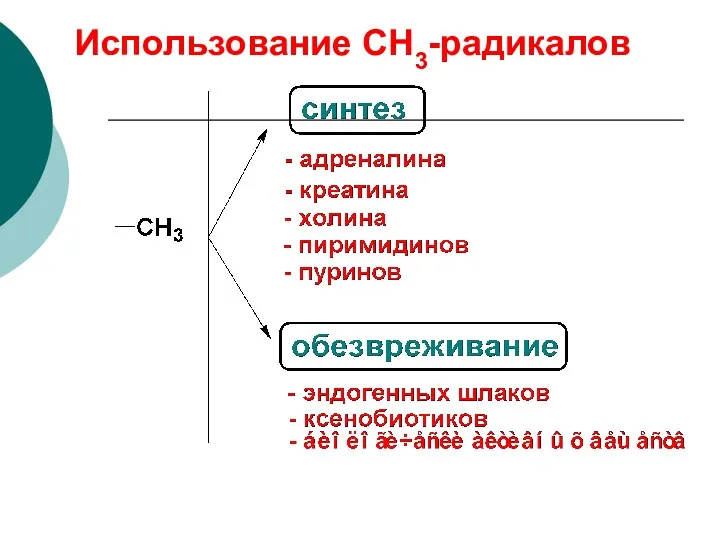
Использование СН3-радикалов
Использование СН3-радикалов
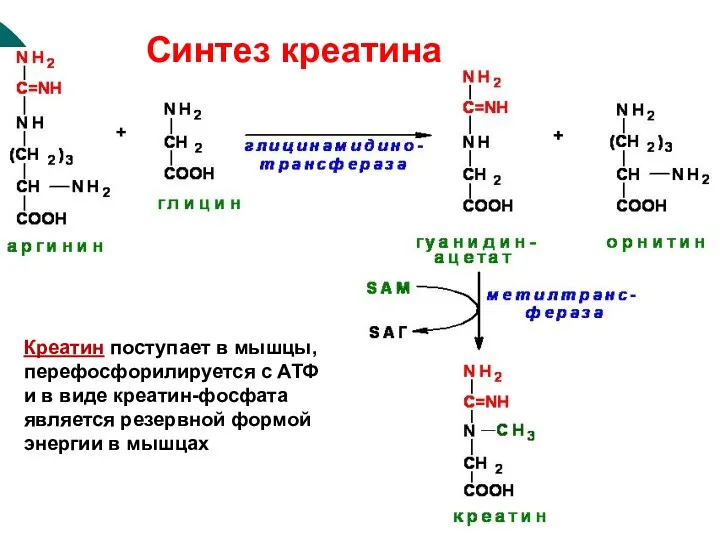
Синтез креатина
Креатин поступает в мышцы, перефосфорилируется с АТФ и в виде
Синтез креатина
Креатин поступает в мышцы, перефосфорилируется с АТФ и в виде
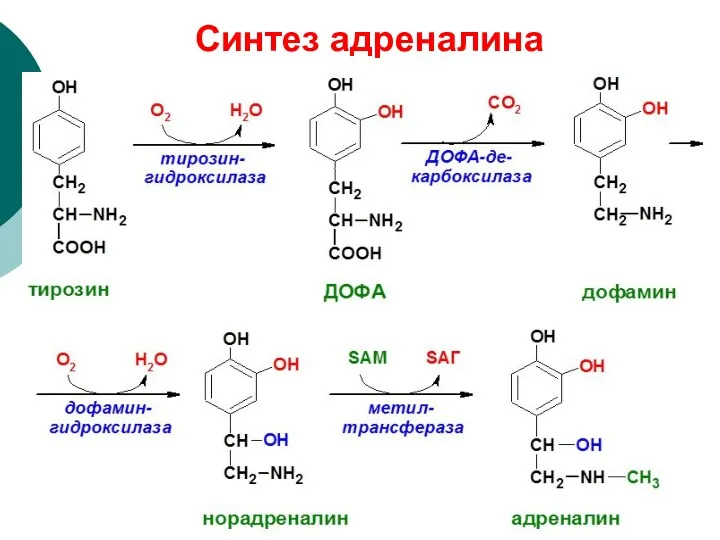
Синтез адреналина
Синтез адреналина
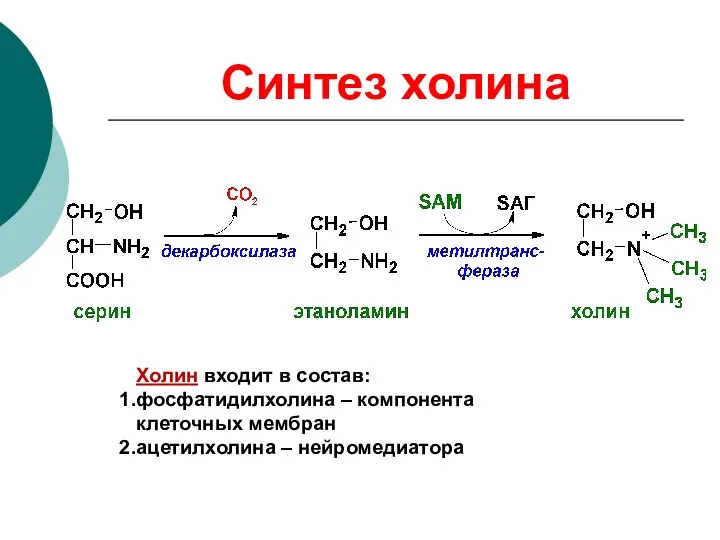
Синтез холина
Холин входит в состав:
фосфатидилхолина – компонента клеточных мембран
ацетилхолина – нейромедиатора
Синтез холина
Холин входит в состав:
фосфатидилхолина – компонента клеточных мембран
ацетилхолина – нейромедиатора
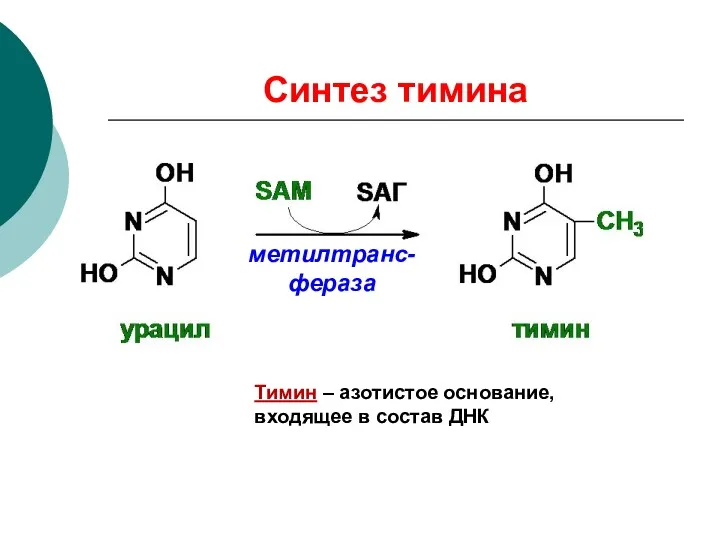
Синтез тимина
Тимин – азотистое основание, входящее в состав ДНК
метилтранс-фераза
Синтез тимина
Тимин – азотистое основание, входящее в состав ДНК
метилтранс-фераза
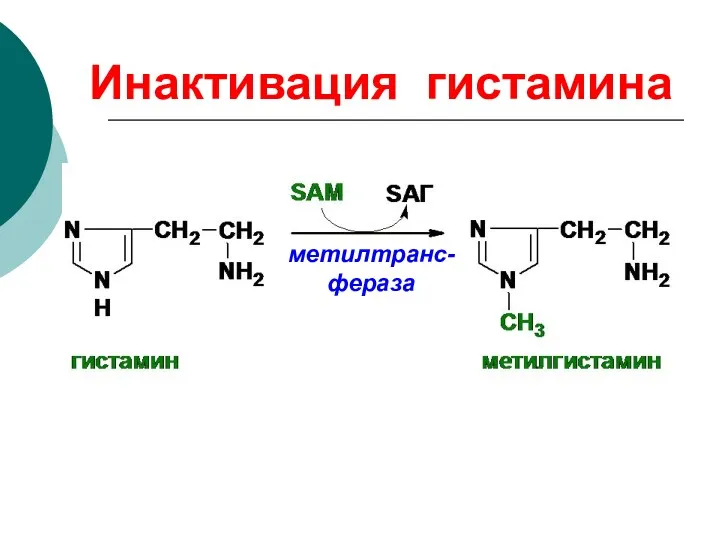
Инактивация гистамина
метилтранс-фераза
Инактивация гистамина
метилтранс-фераза
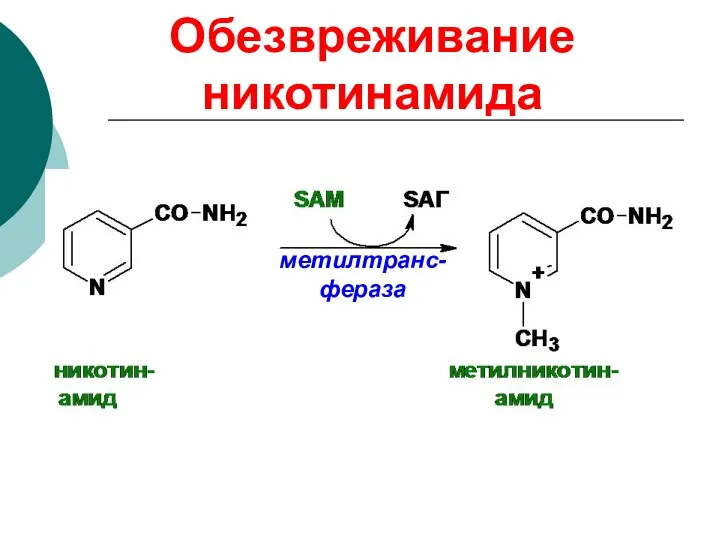
Обезвреживание никотинамида
метилтранс-фераза
Обезвреживание никотинамида
метилтранс-фераза
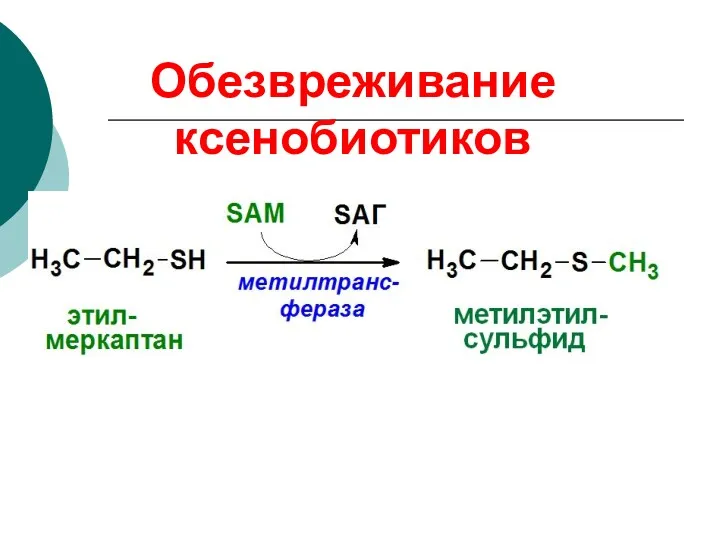
Обезвреживание ксенобиотиков
Обезвреживание ксенобиотиков
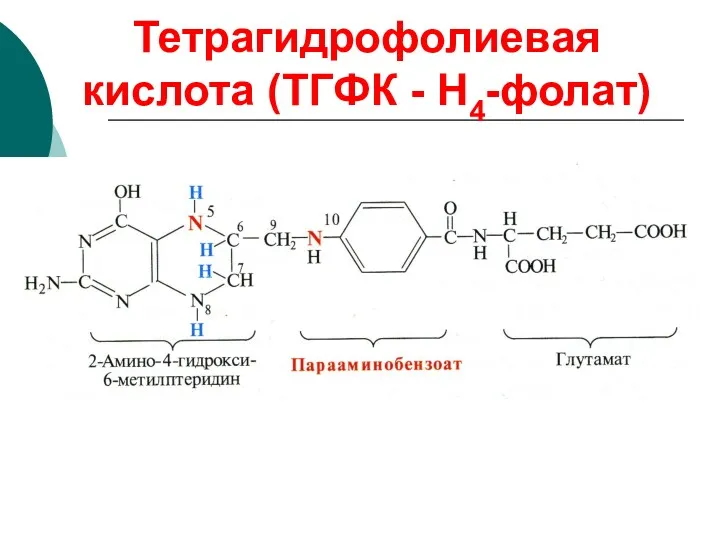
Тетрагидрофолиевая кислота (ТГФК - Н4-фолат)
Тетрагидрофолиевая кислота (ТГФК - Н4-фолат)
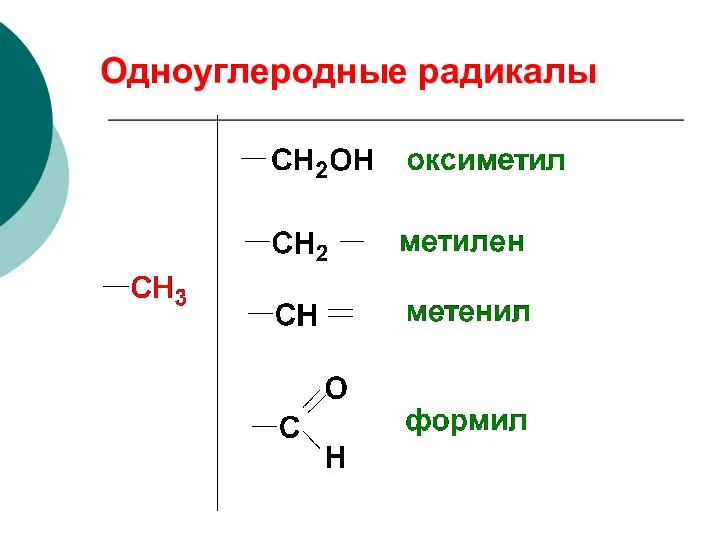
Одноуглеродные радикалы
Одноуглеродные радикалы
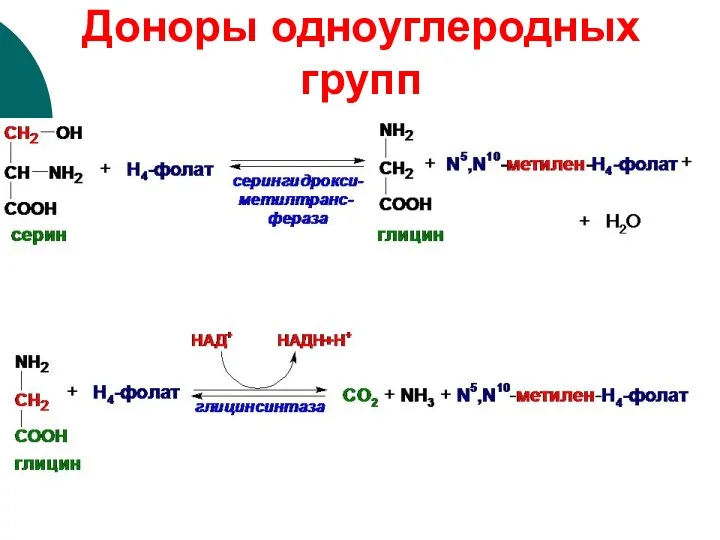
Доноры одноуглеродных групп
Доноры одноуглеродных групп
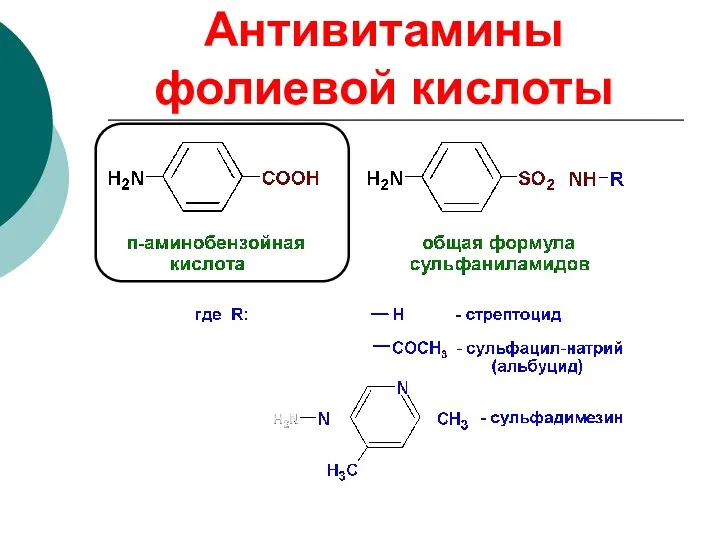
Антивитамины фолиевой кислоты
Антивитамины фолиевой кислоты
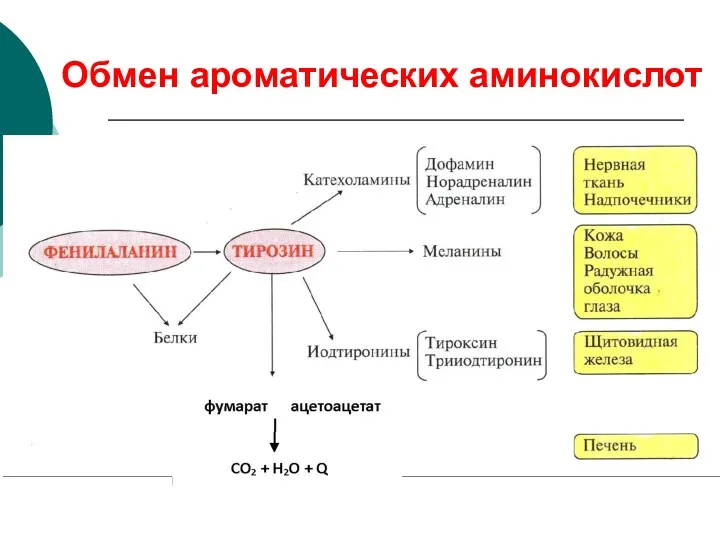
Обмен ароматических аминокислот
Обмен ароматических аминокислот
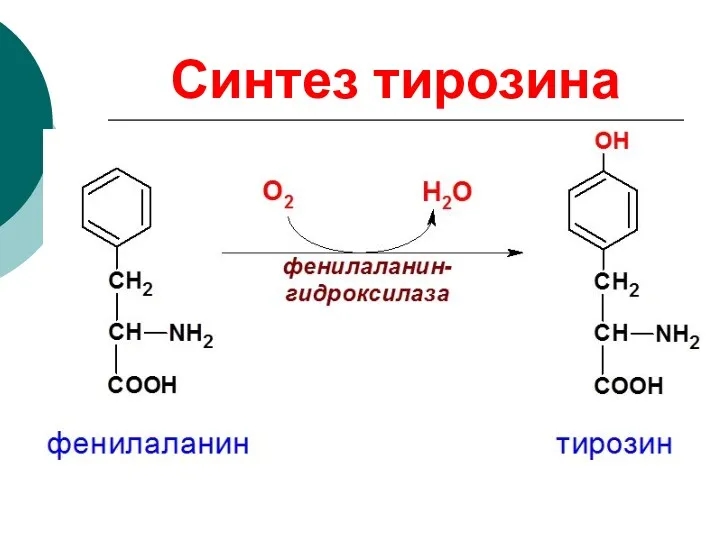
Синтез тирозина
Синтез тирозина
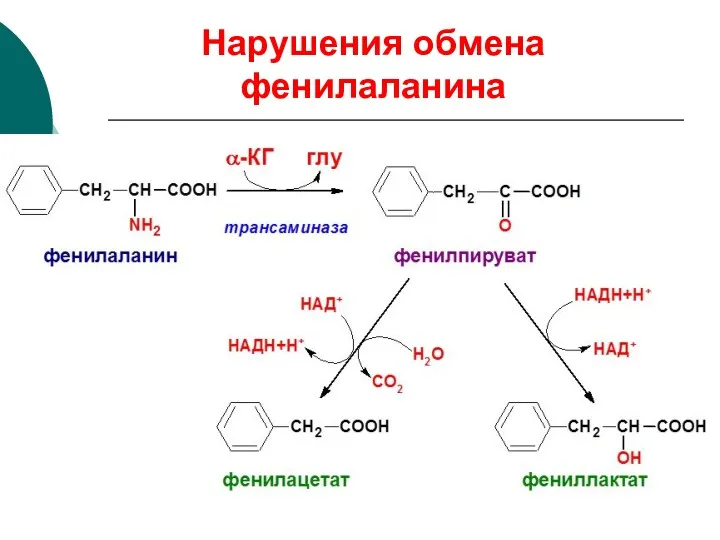
Нарушения обмена фенилаланина
Нарушения обмена фенилаланина

Скрининг-тест – это исследование, которое:
Проводится для 100% популяции (н., для всех
Скрининг-тест – это исследование, которое:
Проводится для 100% популяции (н., для всех
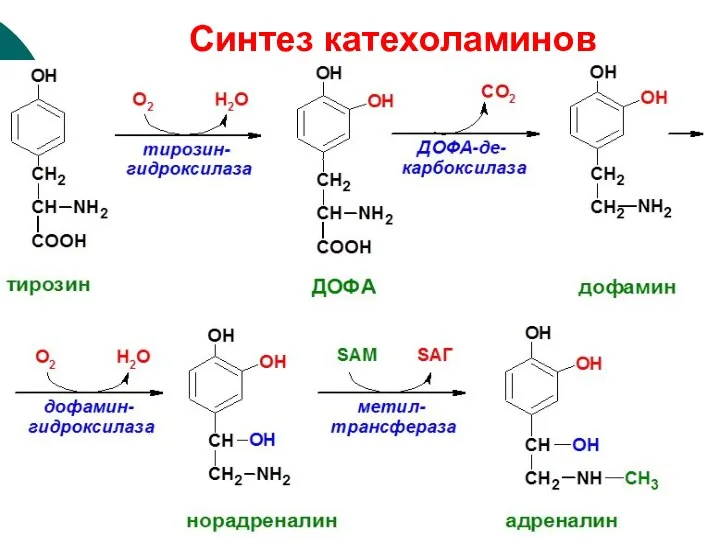
Синтез катехоламинов
Синтез катехоламинов
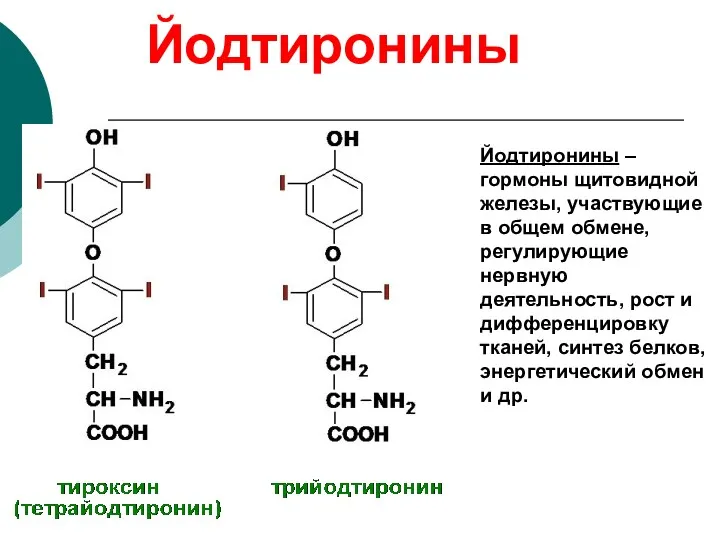
Йодтиронины
Йодтиронины – гормоны щитовидной железы, участвующие в общем обмене, регулирующие нервную
Йодтиронины
Йодтиронины – гормоны щитовидной железы, участвующие в общем обмене, регулирующие нервную
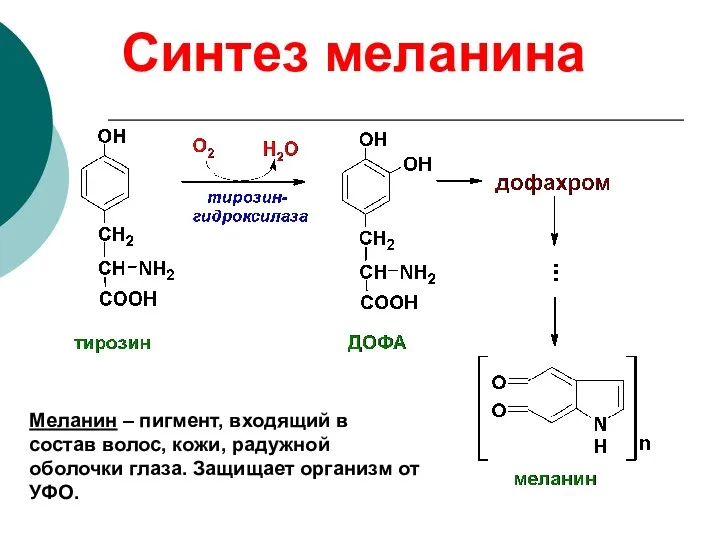
Синтез меланина
Меланин – пигмент, входящий в состав волос, кожи, радужной оболочки
Синтез меланина
Меланин – пигмент, входящий в состав волос, кожи, радужной оболочки
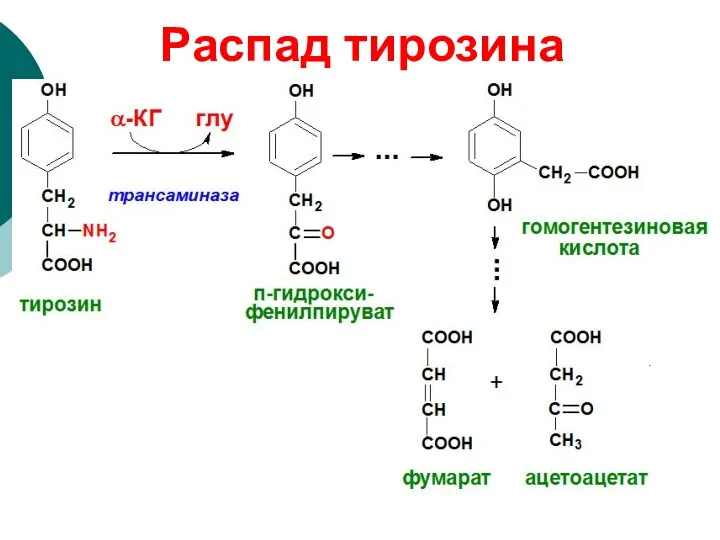
Распад тирозина
Распад тирозина
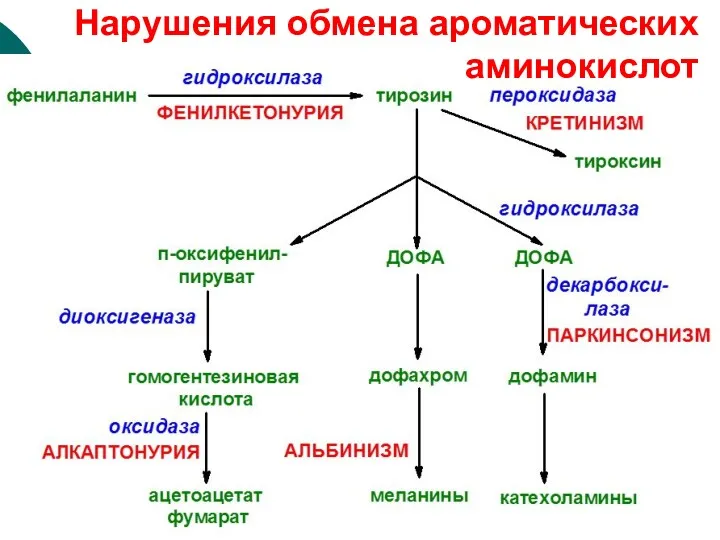
Нарушения обмена ароматических аминокислот
Нарушения обмена ароматических аминокислот
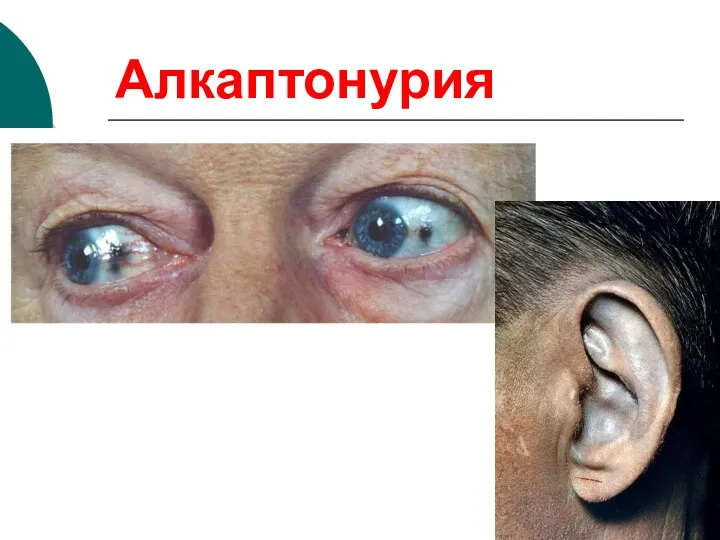
Алкаптонурия
Алкаптонурия

Альбинизм
Альбинизм
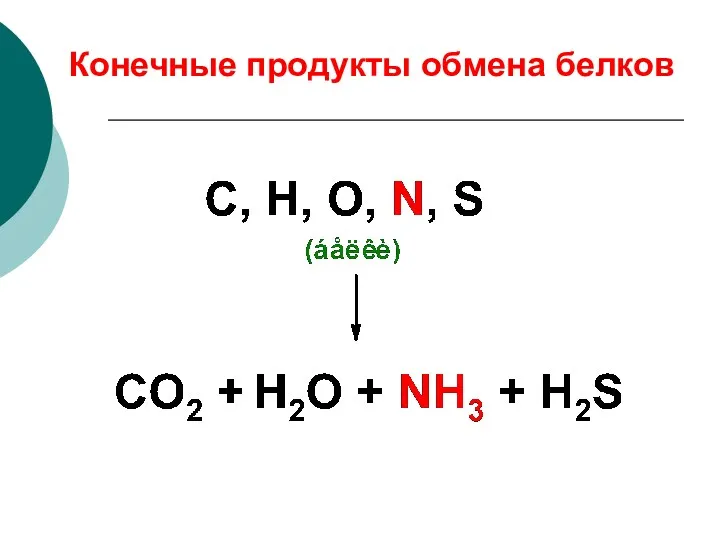
Конечные продукты обмена белков
Конечные продукты обмена белков
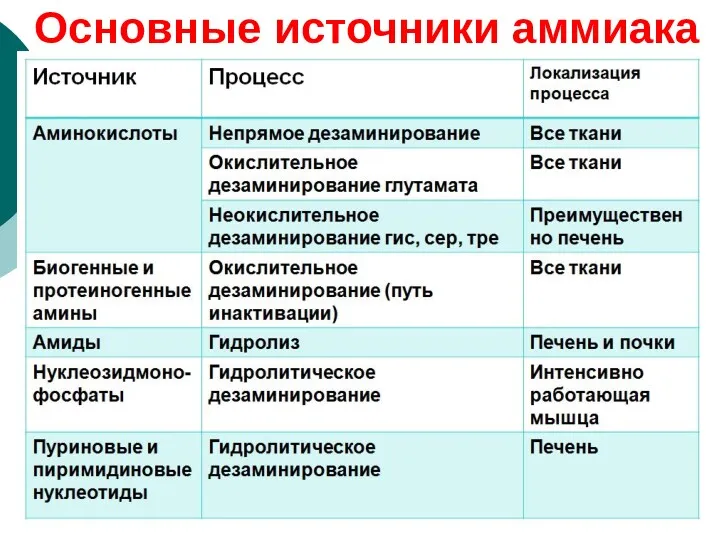
Основные источники аммиака
Основные источники аммиака
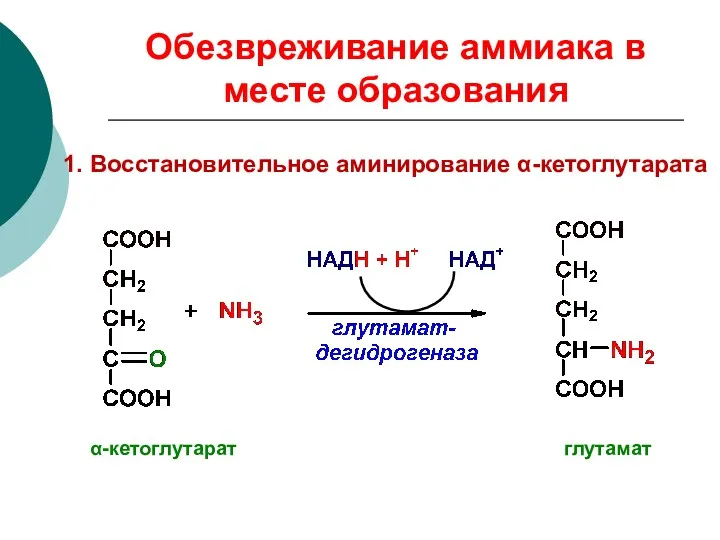
Обезвреживание аммиака в месте образования
1. Восстановительное аминирование α-кетоглутарата
α-кетоглутарат глутамат
Обезвреживание аммиака в месте образования
1. Восстановительное аминирование α-кетоглутарата
α-кетоглутарат глутамат
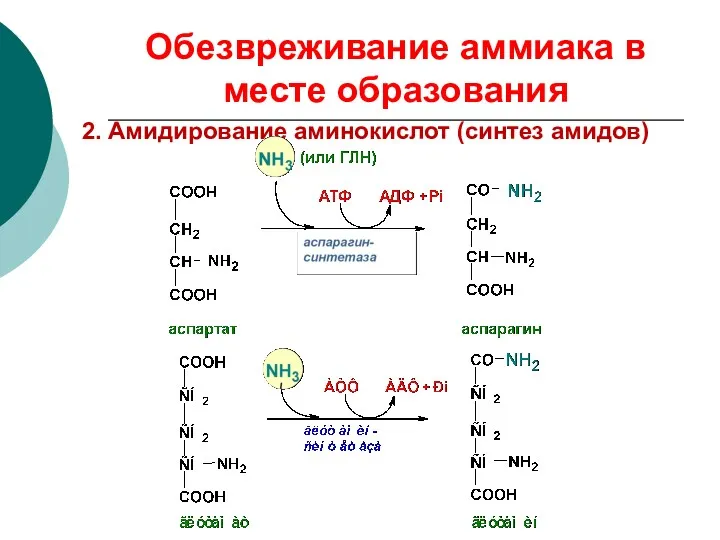
Обезвреживание аммиака в месте образования
2. Амидирование аминокислот (синтез амидов)
Обезвреживание аммиака в месте образования
2. Амидирование аминокислот (синтез амидов)
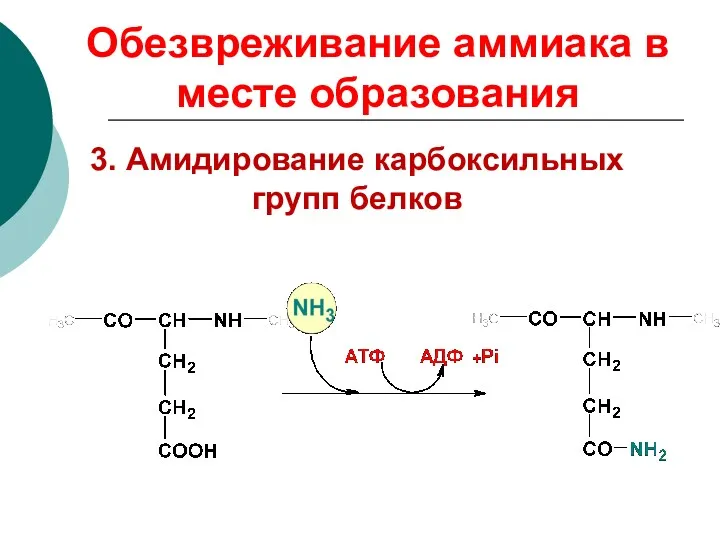
3. Амидирование карбоксильных групп белков
Обезвреживание аммиака в месте образования
3. Амидирование карбоксильных групп белков
Обезвреживание аммиака в месте образования
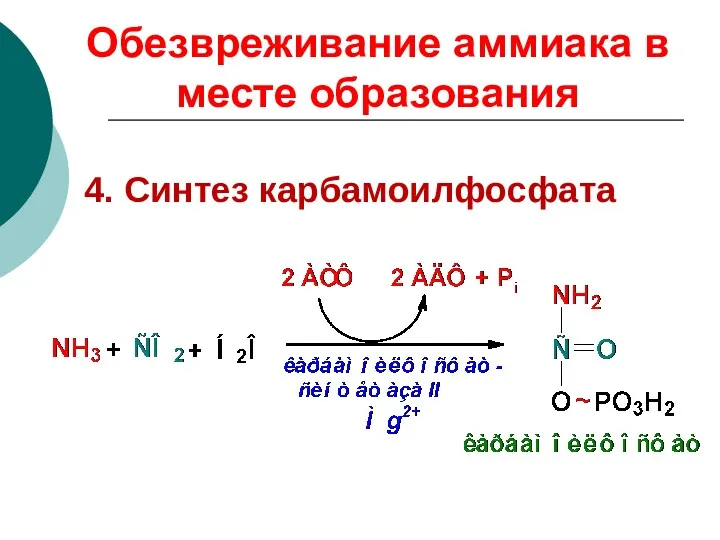
4. Синтез карбамоилфосфата
Обезвреживание аммиака в месте образования
4. Синтез карбамоилфосфата
Обезвреживание аммиака в месте образования

Транспортные формы аммиака
Глутаминовая кислота
Амиды аминокислот (аспарагин и глутамин)
Амидированные белки
Транспортные формы аммиака
Глутаминовая кислота
Амиды аминокислот (аспарагин и глутамин)
Амидированные белки
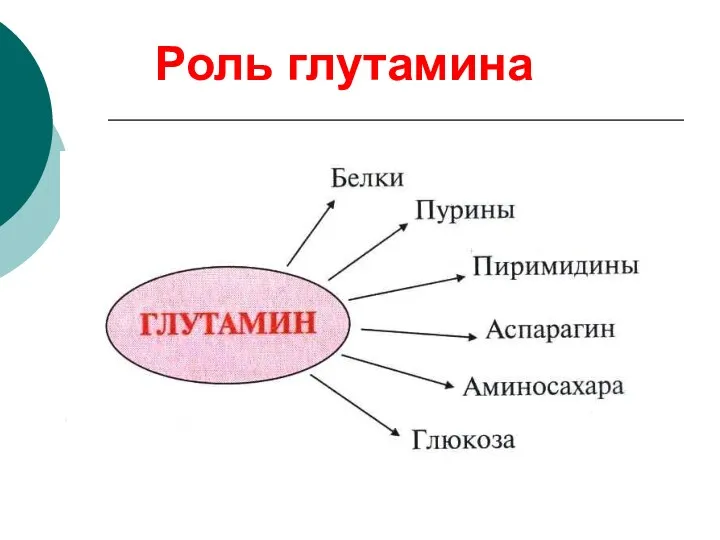
Роль глутамина
Роль глутамина
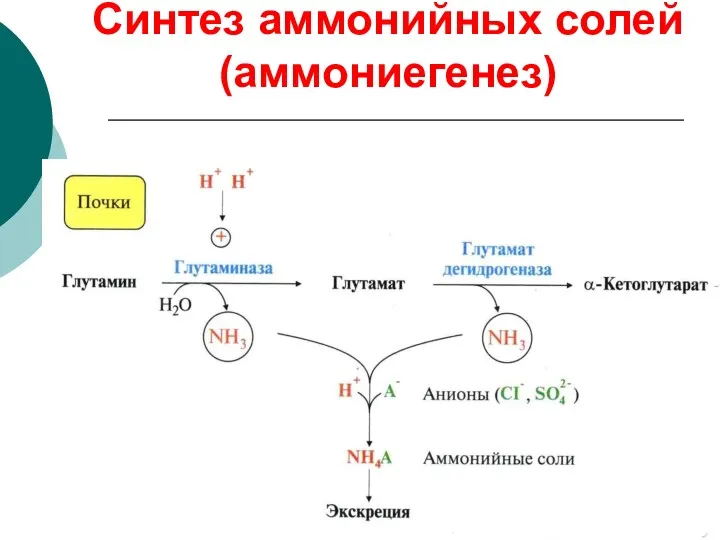
Синтез аммонийных солей (аммониегенез)
Синтез аммонийных солей (аммониегенез)

Биологическая роль аммониегенеза
Обезвреживание и выведение токсического аммиака
Поддержание кислотно-основного состояния (КОС) за
Биологическая роль аммониегенеза
Обезвреживание и выведение токсического аммиака
Поддержание кислотно-основного состояния (КОС) за
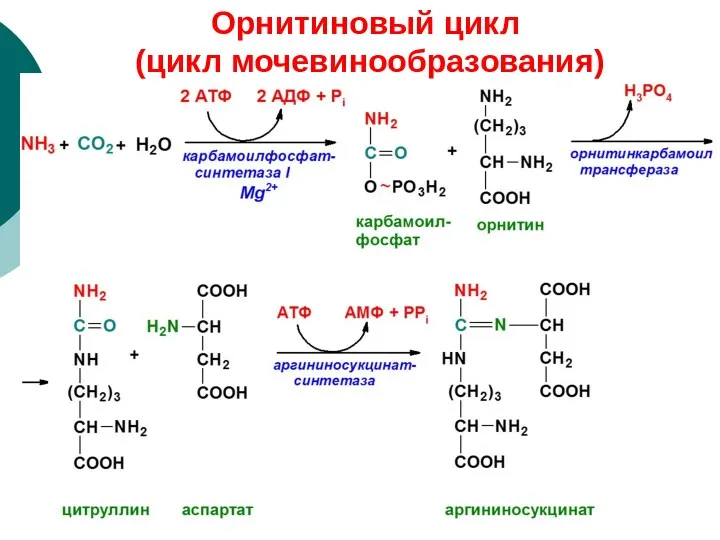
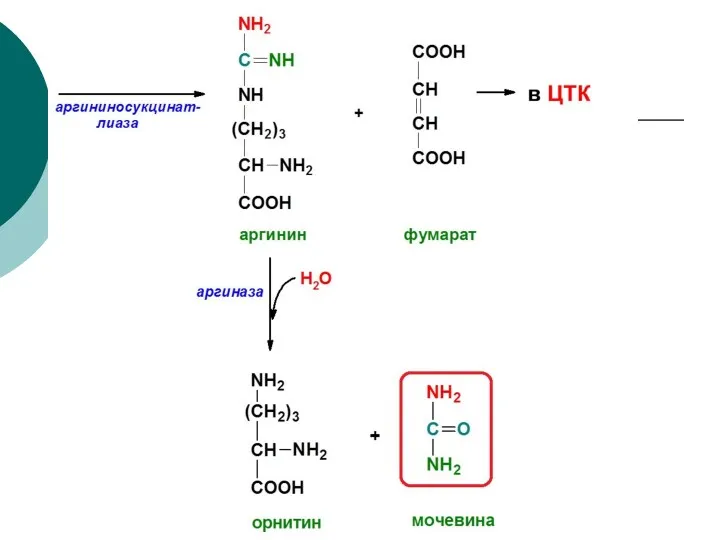
Орнитиновый цикл (цикл мочевинообразования)
Орнитиновый цикл (цикл мочевинообразования)

Особенности цикла мочевинообразования
На синтез одной молекулы мочевины затрачивается 3 молекулы АТФ
Особенности цикла мочевинообразования
На синтез одной молекулы мочевины затрачивается 3 молекулы АТФ

Биологическая роль орнитинового цикла
Конечное обезвреживание токсического аммиака
Синтез аргинина и пополнение его
Биологическая роль орнитинового цикла
Конечное обезвреживание токсического аммиака
Синтез аргинина и пополнение его

Связь цикла мочевинообразования и ЦТК
Оба цикла протекают в одном и том
Связь цикла мочевинообразования и ЦТК
Оба цикла протекают в одном и том
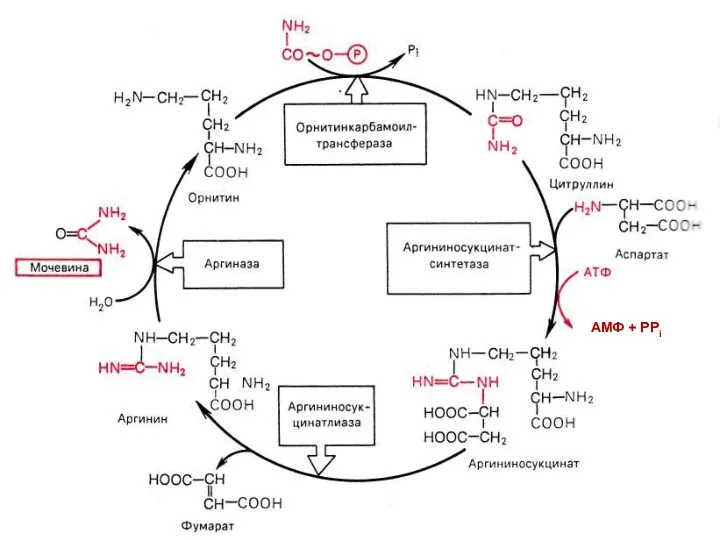
АМФ + РРi
АМФ + РРi
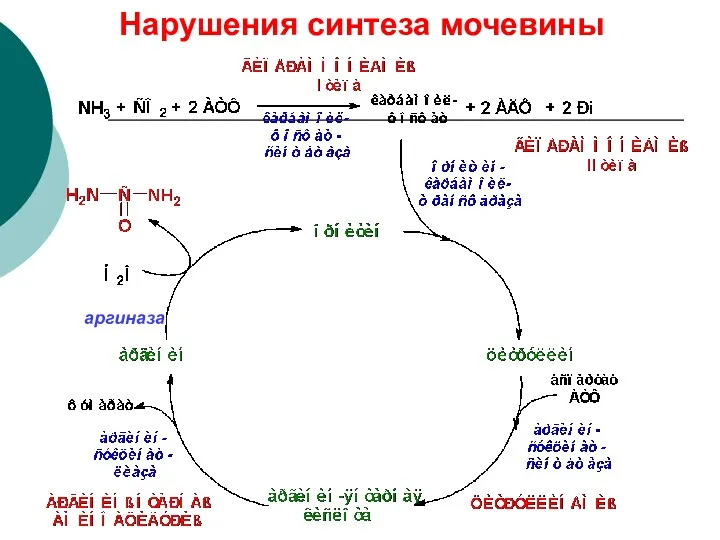
Нарушения синтеза мочевины
аргиназа
Нарушения синтеза мочевины
аргиназа
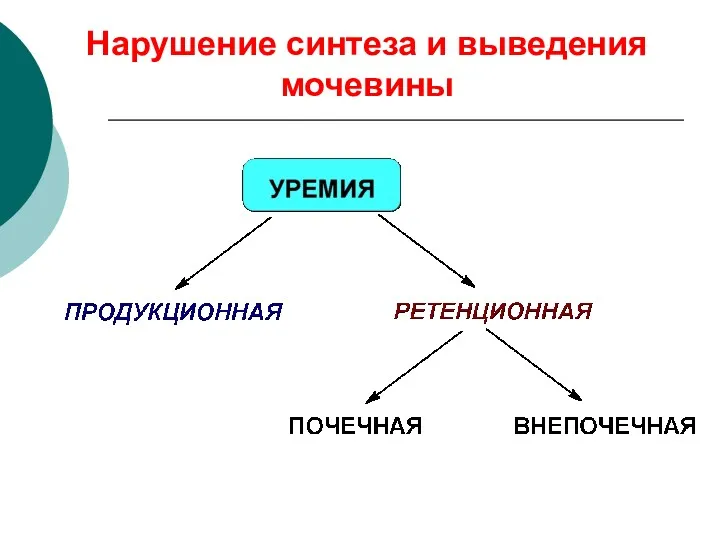
Нарушение синтеза и выведения мочевины
Нарушение синтеза и выведения мочевины
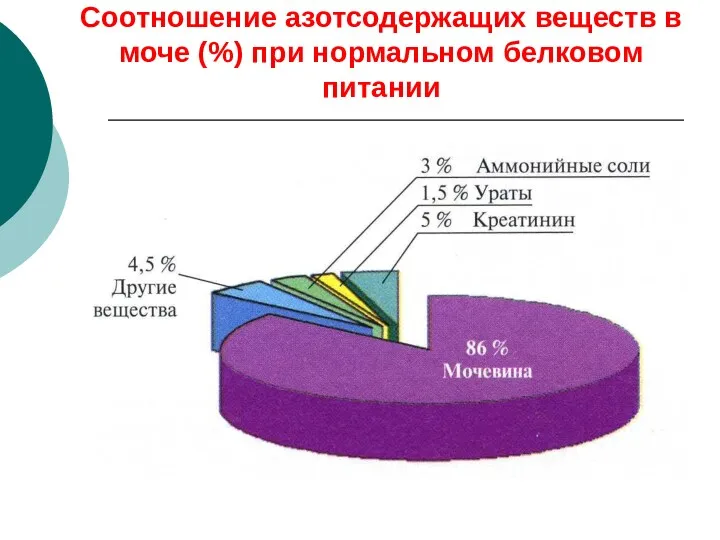
Соотношение азотсодержащих веществ в моче (%) при нормальном белковом питании
Соотношение азотсодержащих веществ в моче (%) при нормальном белковом питании

КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ
КАФЕДРА ФУНДАМЕНТАЛЬНОЙ И КЛИНИЧЕСКОЙ БИОХИМИИ
Лекция по биохимии
Тема:
«Обмен белков–3»
Краснодар
2017
КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ
КАФЕДРА ФУНДАМЕНТАЛЬНОЙ И КЛИНИЧЕСКОЙ БИОХИМИИ
Лекция по биохимии
Тема:
«Обмен белков–3»
Краснодар
2017
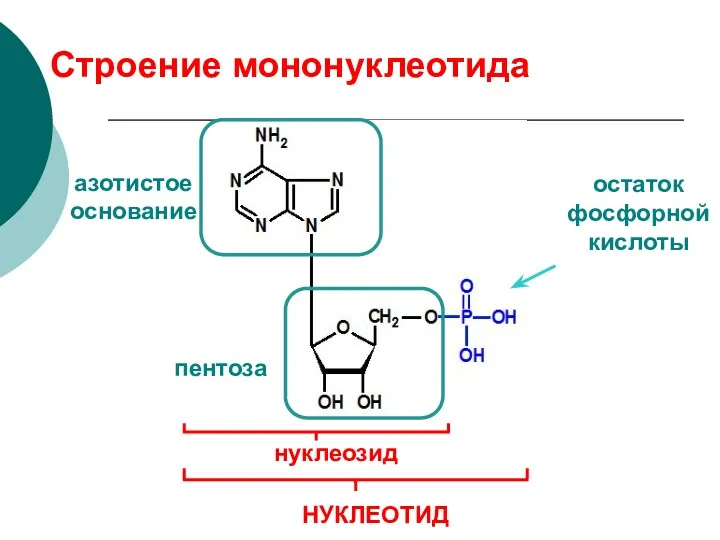
Строение мононуклеотида
азотистое основание
пентоза
остаток фосфорной кислоты
нуклеозид
НУКЛЕОТИД
Строение мононуклеотида
азотистое основание
пентоза
остаток фосфорной кислоты
нуклеозид
НУКЛЕОТИД
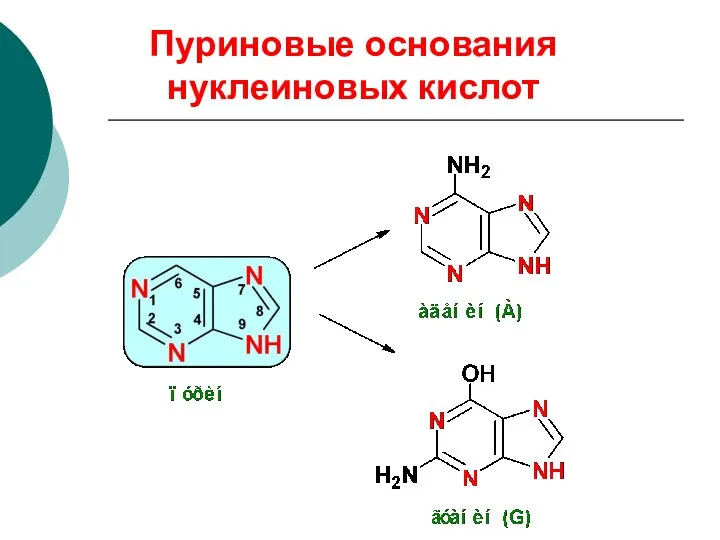
Пуриновые основания нуклеиновых кислот
Пуриновые основания нуклеиновых кислот
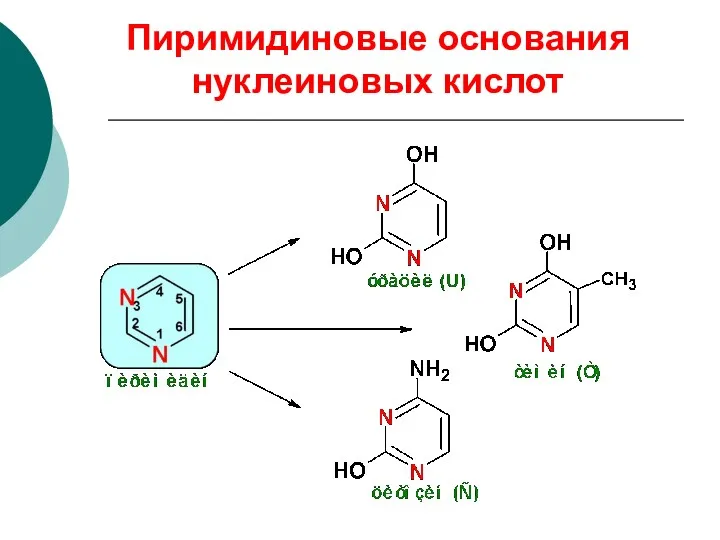
Пиримидиновые основания нуклеиновых кислот
Пиримидиновые основания нуклеиновых кислот
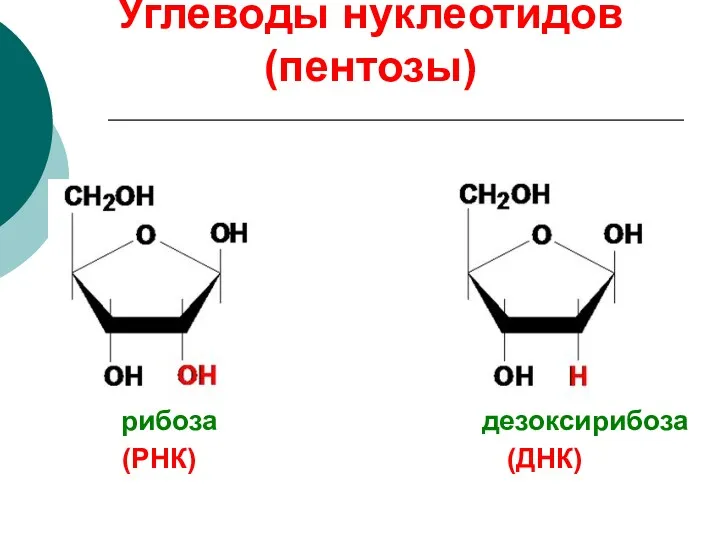
Углеводы нуклеотидов (пентозы)
(РНК) (ДНК)
рибоза дезоксирибоза
Углеводы нуклеотидов (пентозы)
(РНК) (ДНК)
рибоза дезоксирибоза
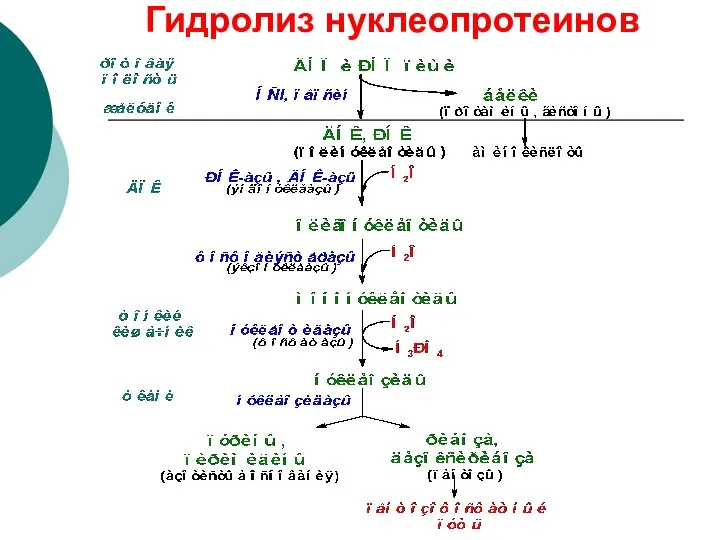
Гидролиз нуклеопротеинов
Гидролиз нуклеопротеинов
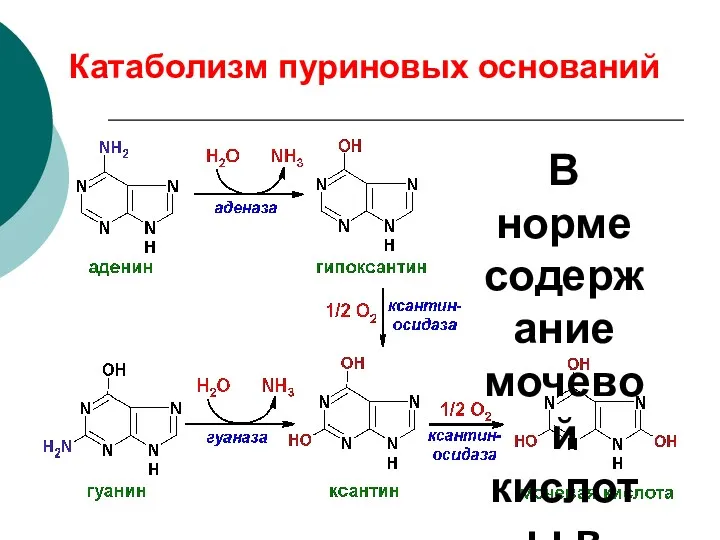
Катаболизм пуриновых оснований
В норме содержание мочевой кислоты в крови 0,14-0,5 ммоль/л
Катаболизм пуриновых оснований
В норме содержание мочевой кислоты в крови 0,14-0,5 ммоль/л
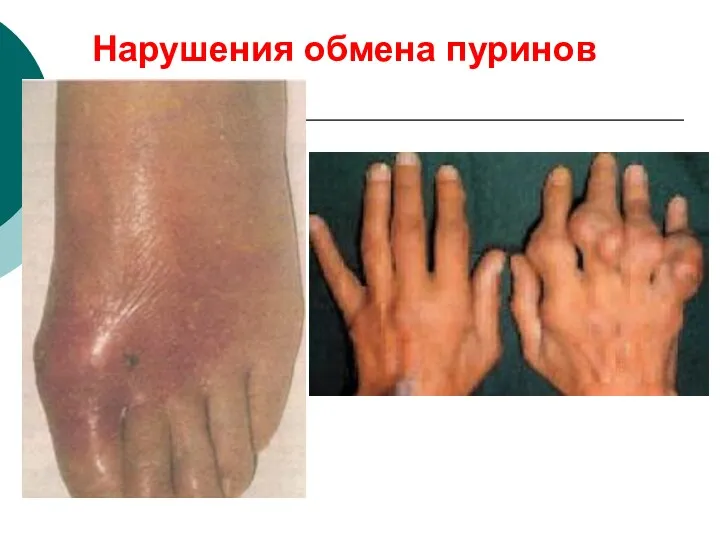
Нарушения обмена пуринов
Нарушения обмена пуринов
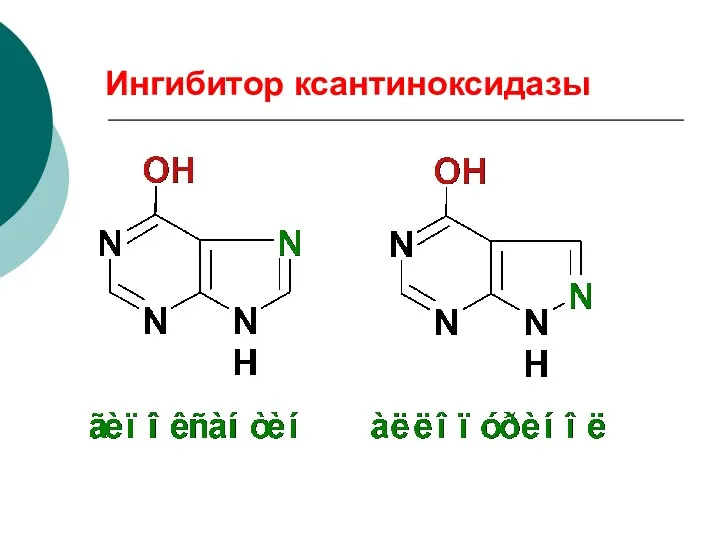
Ингибитор ксантиноксидазы
Ингибитор ксантиноксидазы
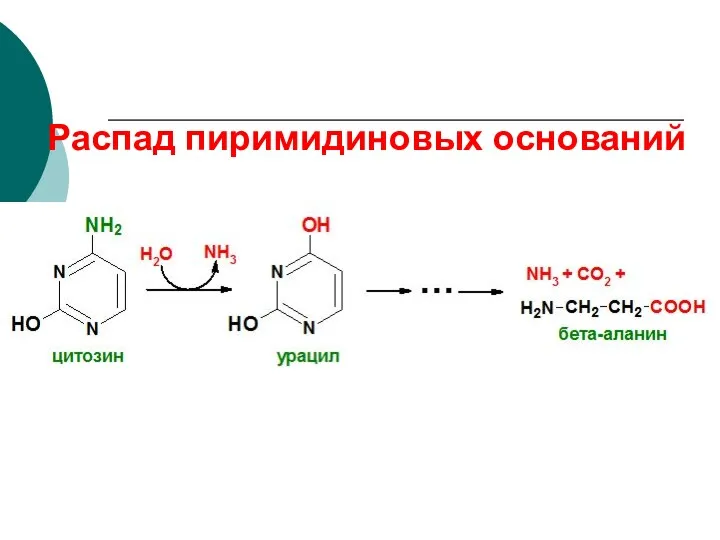
Распад пиримидиновых оснований
Распад пиримидиновых оснований

Особенности синтеза нуклеотидов
Синтез идёт из обычных простых предшественников (ак, углекислого газа
Особенности синтеза нуклеотидов
Синтез идёт из обычных простых предшественников (ак, углекислого газа
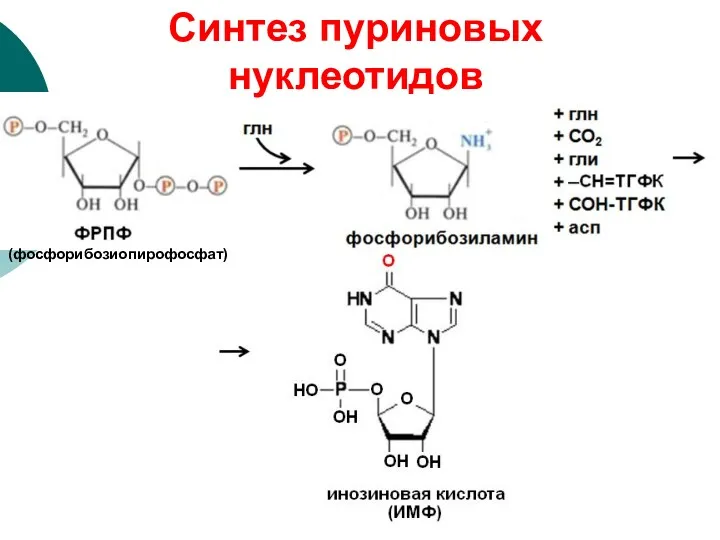
(фосфорибозиопирофосфат)
Синтез пуриновых нуклеотидов
(фосфорибозиопирофосфат)
Синтез пуриновых нуклеотидов
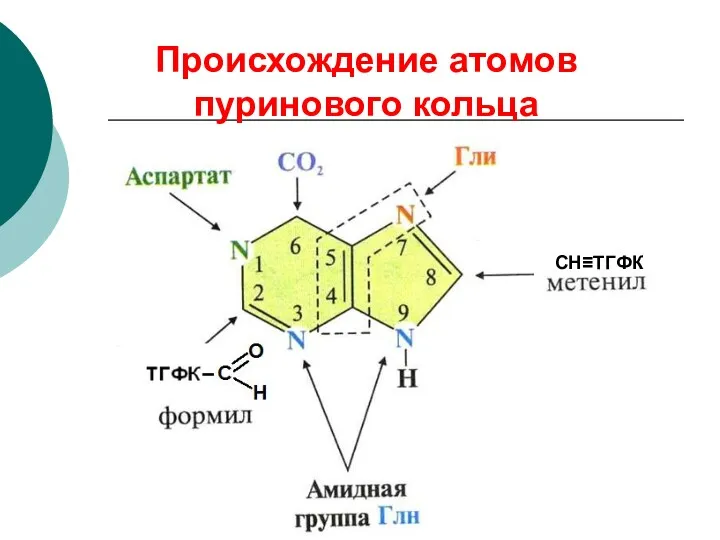
Происхождение атомов пуринового кольца
СН≡ТГФК
Происхождение атомов пуринового кольца
СН≡ТГФК
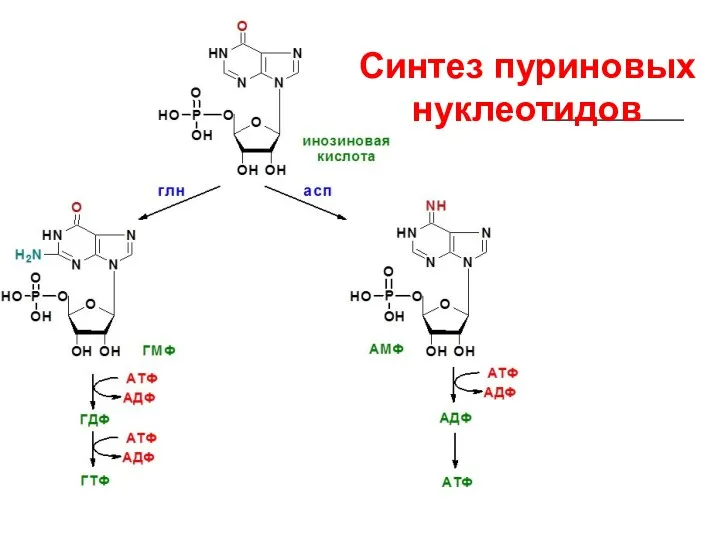
Синтез пуриновых нуклеотидов
Синтез пуриновых нуклеотидов
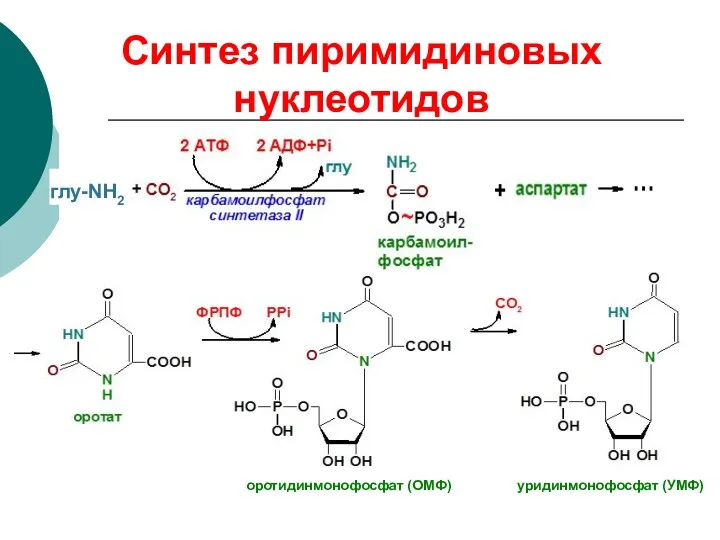
Синтез пиримидиновых нуклеотидов
глу-NH2
оротидинмонофосфат (ОМФ) уридинмонофосфат (УМФ)
Синтез пиримидиновых нуклеотидов
глу-NH2
оротидинмонофосфат (ОМФ) уридинмонофосфат (УМФ)
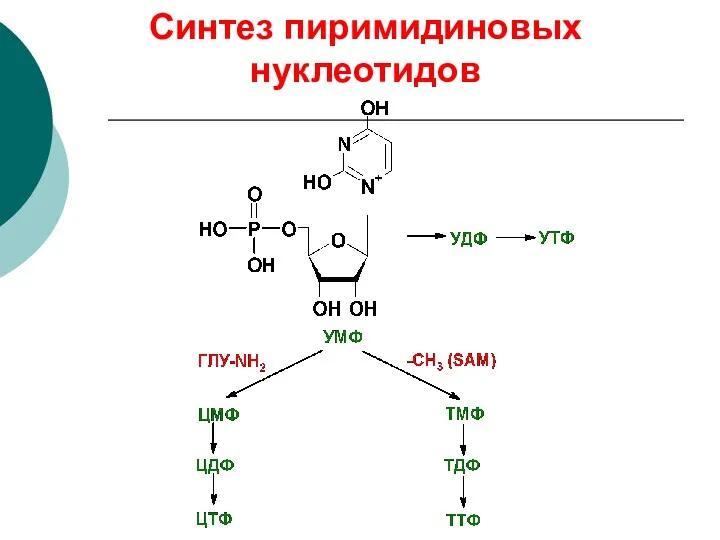
Синтез пиримидиновых нуклеотидов
Синтез пиримидиновых нуклеотидов
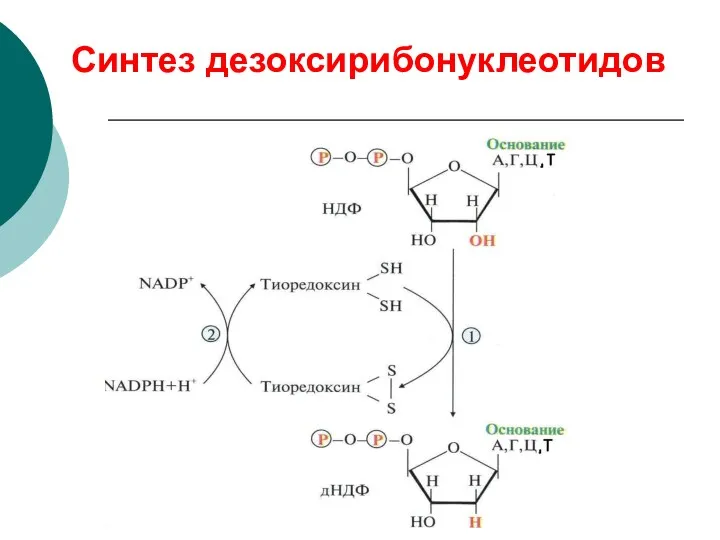
Синтез дезоксирибонуклеотидов
Синтез дезоксирибонуклеотидов
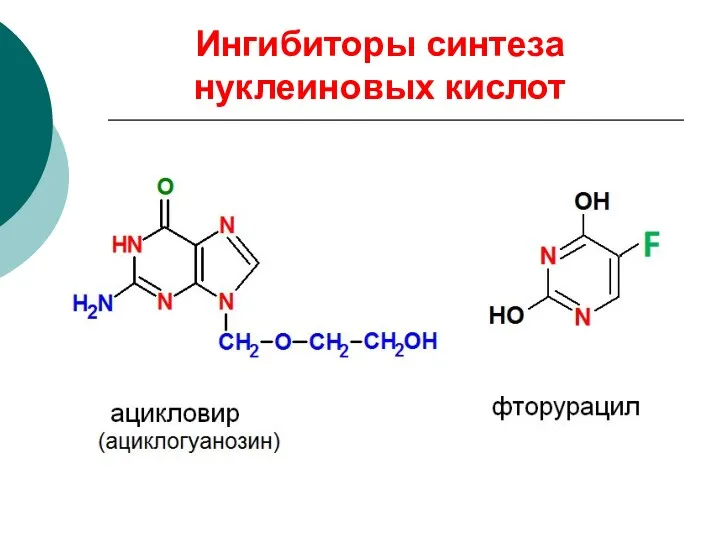
Ингибиторы синтеза нуклеиновых кислот
Ингибиторы синтеза нуклеиновых кислот

Нарушения обмена нуклеотидов
Нарушение синтеза пиримидинов – оротацидурия – дефект ОМФ-декарбоксилазы (недостаток
Нарушения обмена нуклеотидов
Нарушение синтеза пиримидинов – оротацидурия – дефект ОМФ-декарбоксилазы (недостаток
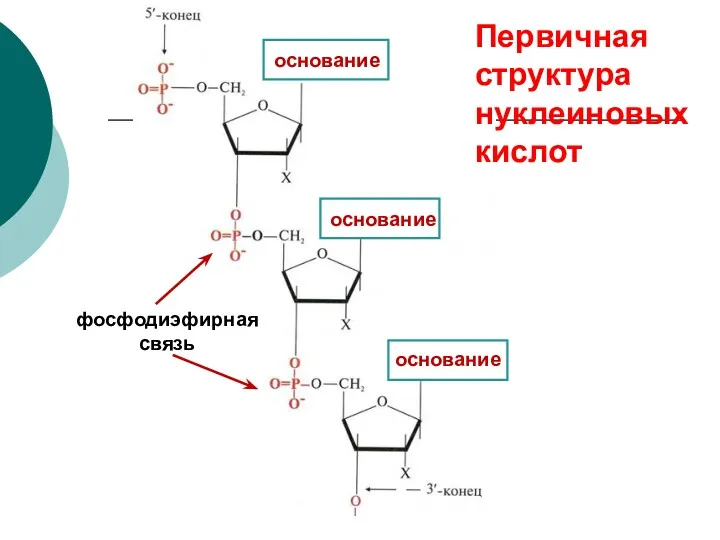
основание
фосфодиэфирная связь
Первичная структура нуклеиновых кислот
основание
основание
основание
фосфодиэфирная связь
Первичная структура нуклеиновых кислот
основание
основание
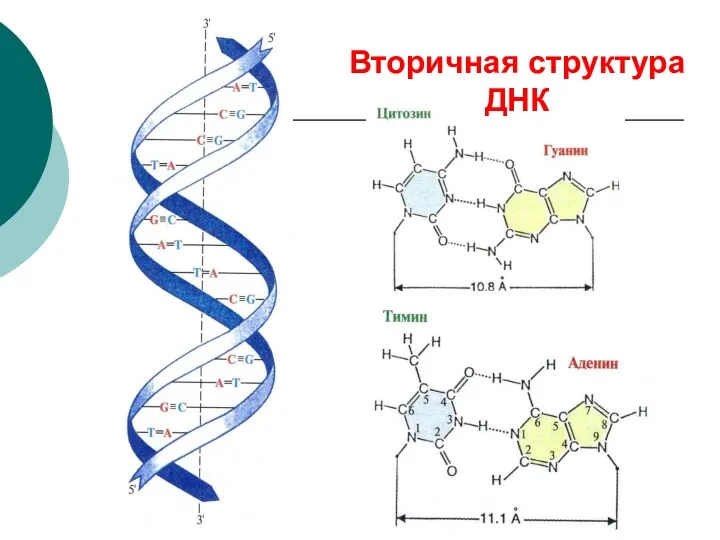
Вторичная структура ДНК
Вторичная структура ДНК
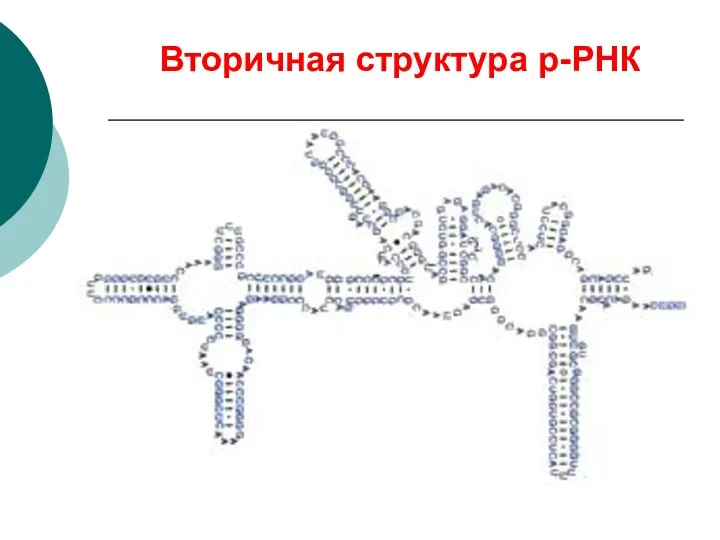
Вторичная структура р-РНК
Вторичная структура р-РНК
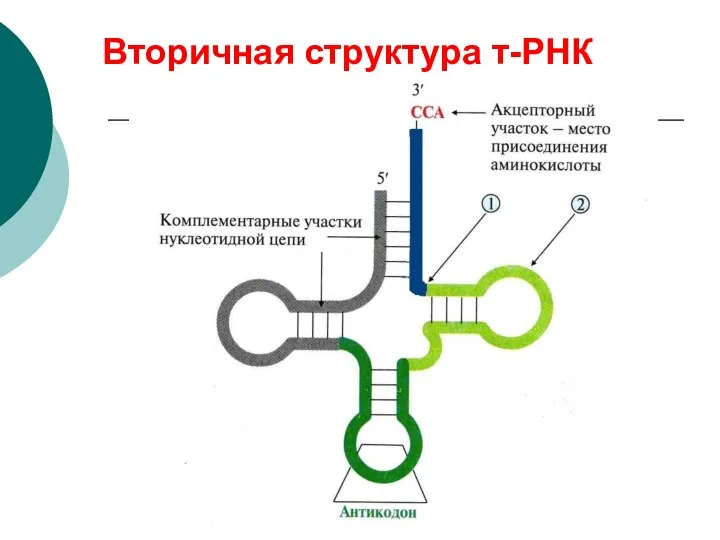
Вторичная структура т-РНК
Вторичная структура т-РНК
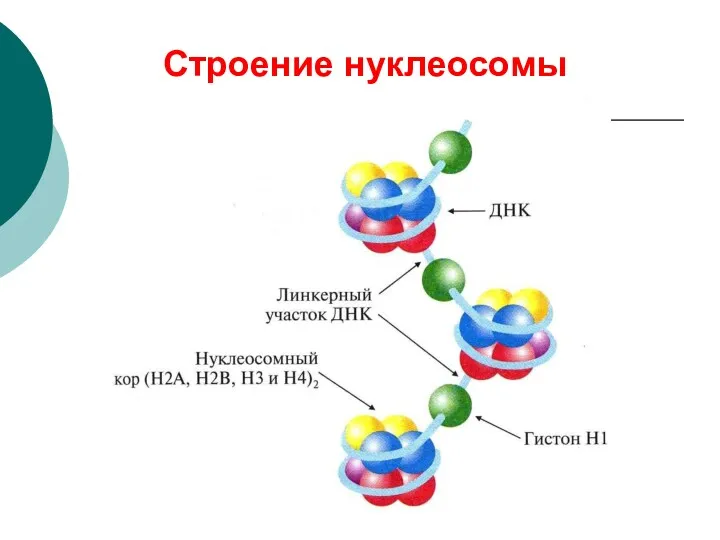
Строение нуклеосомы
Строение нуклеосомы
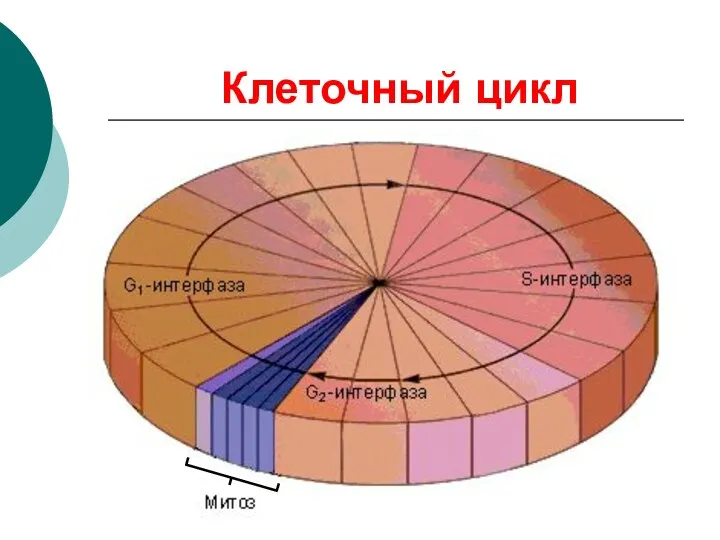
Клеточный цикл
Клеточный цикл
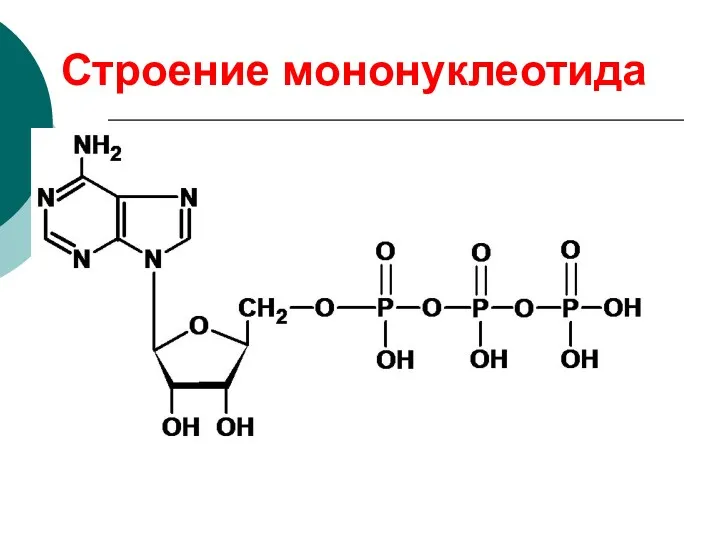
Строение мононуклеотида
Строение мононуклеотида
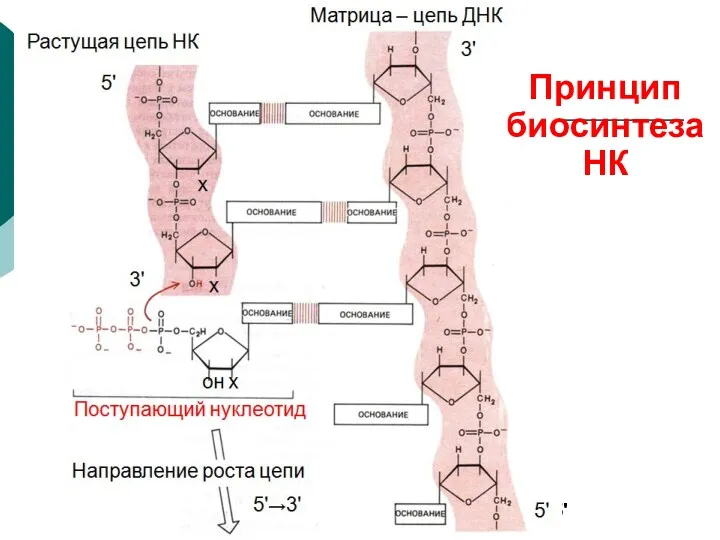
Направление роста цепи
5'→3'
Поступающий нуклеотид
он х
х
х
Растущая цепь НК
Матрица – цепь ДНК
5'
5'
3'
3'
Принцип биосинтеза
НК
Направление роста цепи
5'→3'
Поступающий нуклеотид
он х
х
х
Растущая цепь НК
Матрица – цепь ДНК
5'
5'
3'
3'
Принцип биосинтеза
НК

Биосинтез ДНК (репликация) является:
матричным (матрица – обе нити ДНК)
комплиментарным
фрагментарным (нити ДНК
Биосинтез ДНК (репликация) является:
матричным (матрица – обе нити ДНК)
комплиментарным
фрагментарным (нити ДНК
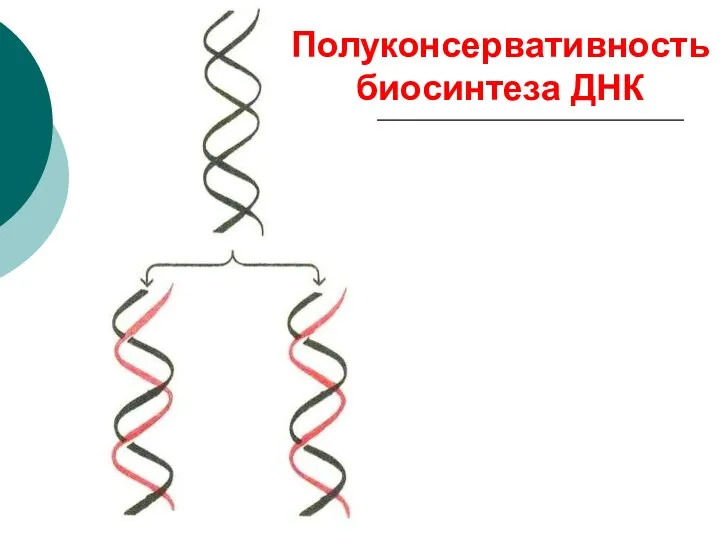
Полуконсервативность биосинтеза ДНК
Полуконсервативность биосинтеза ДНК


Особенности репликации
ДНК-полимеразы δ и ε не могут соединять между собой два
Особенности репликации
ДНК-полимеразы δ и ε не могут соединять между собой два

Этапы репликации
1. Инициация:
Топоизомераза находит точку начала репликации, гидролизует одну фосфодиэфирную
Этапы репликации
1. Инициация:
Топоизомераза находит точку начала репликации, гидролизует одну фосфодиэфирную

Этапы репликации
ДНК-связывающие белки (SSB-белки) стабилизируют репликативную вилку, не давая восстанавливаться водородным
Этапы репликации
ДНК-связывающие белки (SSB-белки) стабилизируют репликативную вилку, не давая восстанавливаться водородным
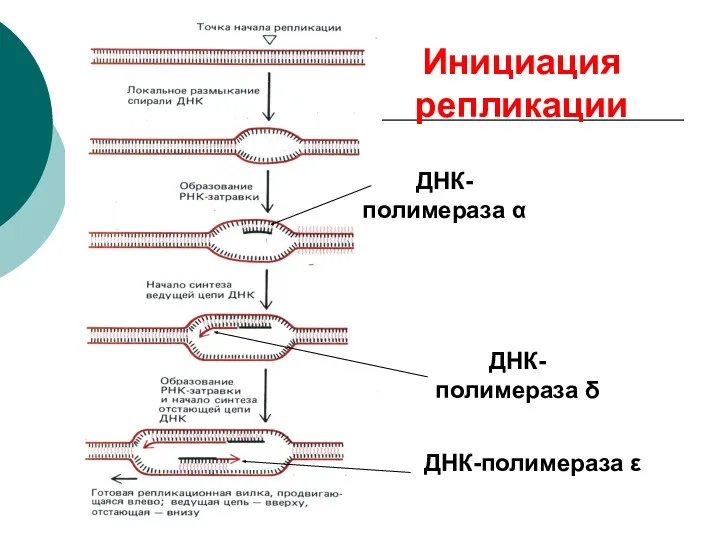
Инициация репликации
ДНК-полимераза α
ДНК-полимераза δ
ДНК-полимераза ε
Инициация репликации
ДНК-полимераза α
ДНК-полимераза δ
ДНК-полимераза ε
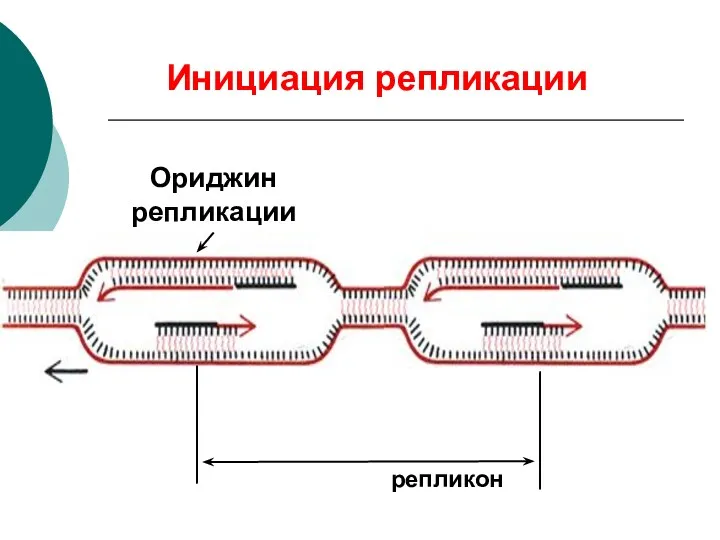
Ориджин репликации
репликон
Инициация репликации
Ориджин репликации
репликон
Инициация репликации

2. Элонгация
ДНК-полимераза δ продолжает удлинять нить из дезоксирибонуклеотидов на лидирующей
2. Элонгация
ДНК-полимераза δ продолжает удлинять нить из дезоксирибонуклеотидов на лидирующей
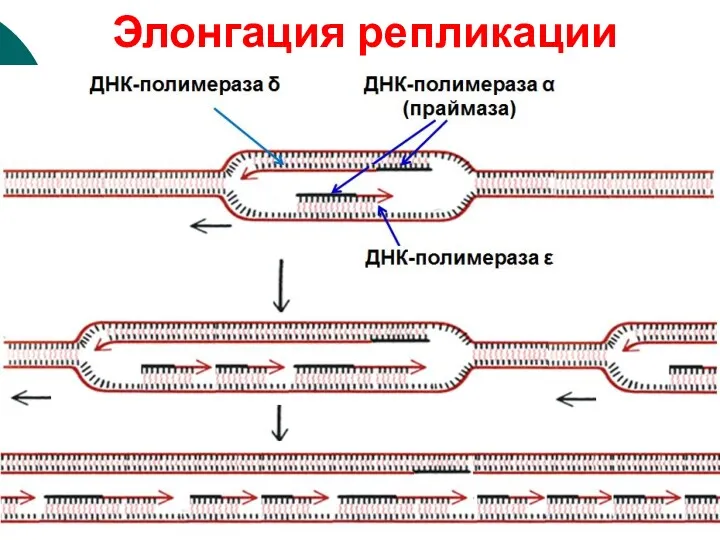
Элонгация репликации
Элонгация репликации

Терминация
ДНК-полимераза β (фермент репарации) удаляет праймеры и достраивает фрагменты ДНК
ДНК-лигаза
Терминация
ДНК-полимераза β (фермент репарации) удаляет праймеры и достраивает фрагменты ДНК
ДНК-лигаза
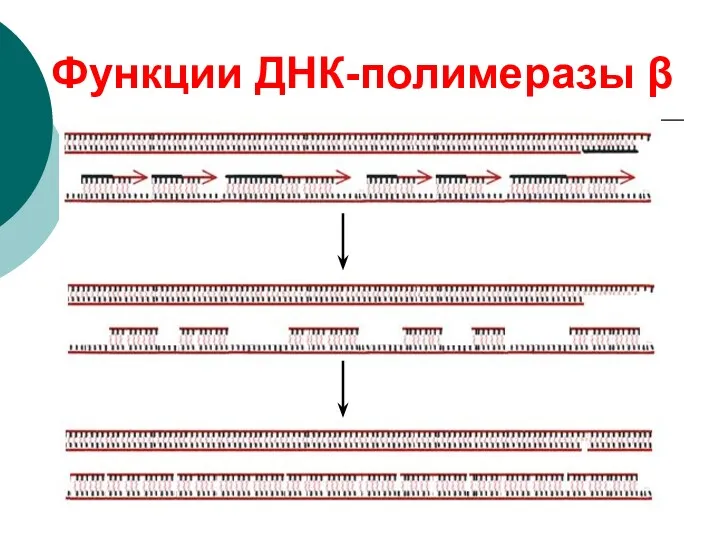
Функции ДНК-полимеразы β
Функции ДНК-полимеразы β
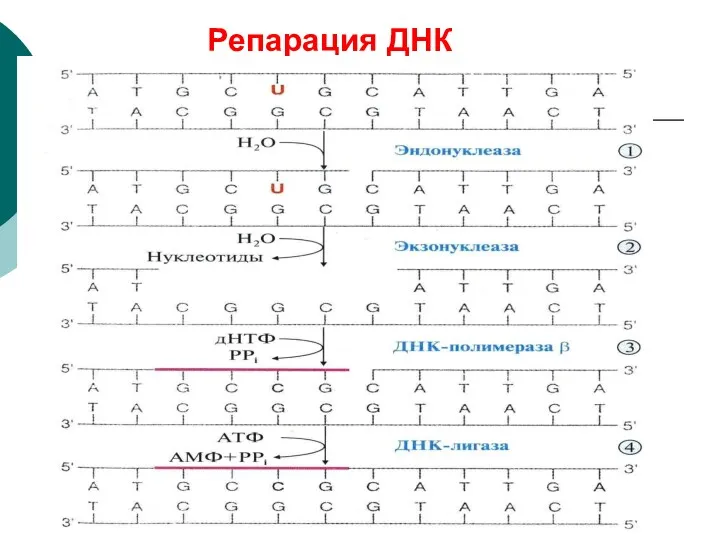
Репарация ДНК
Репарация ДНК

Для биосинтеза РНК (транскрипции) необходимы:
МАТРИЦА – участок одной из нитей ДНК
Для биосинтеза РНК (транскрипции) необходимы:
МАТРИЦА – участок одной из нитей ДНК
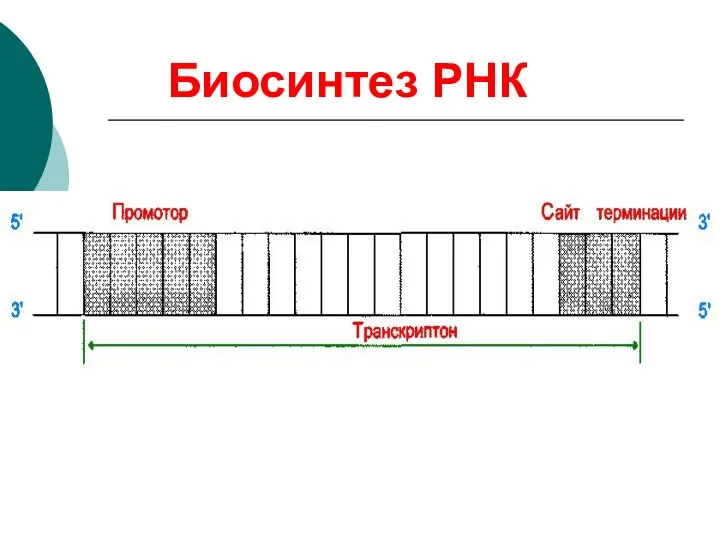
Биосинтез РНК
Биосинтез РНК
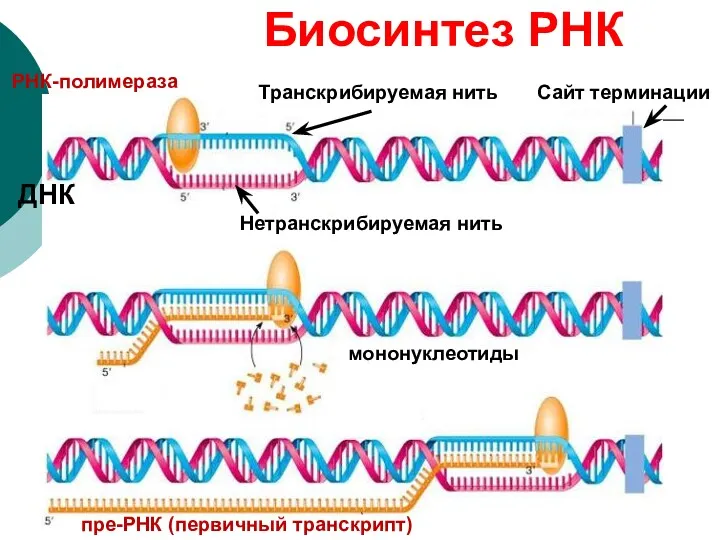
Транскрибируемая нить
Нетранскрибируемая нить
РНК-полимераза
ДНК
пре-РНК (первичный транскрипт)
Биосинтез РНК
мононуклеотиды
Сайт терминации
Транскрибируемая нить
Нетранскрибируемая нить
РНК-полимераза
ДНК
пре-РНК (первичный транскрипт)
Биосинтез РНК
мононуклеотиды
Сайт терминации
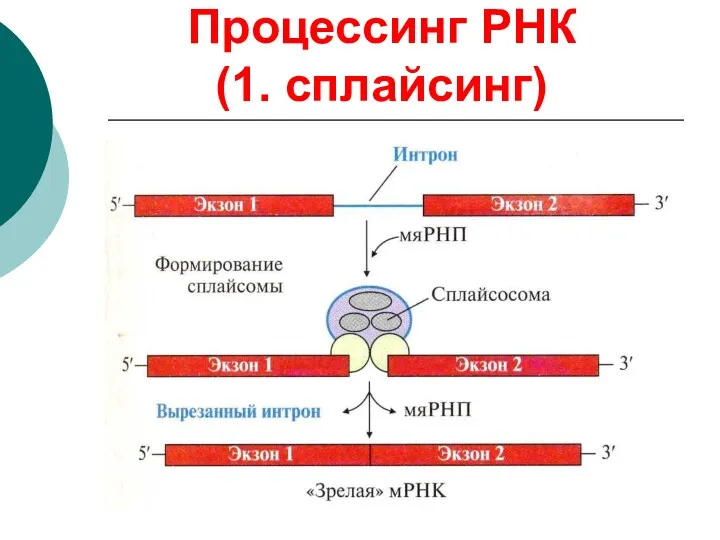
Процессинг РНК
(1. сплайсинг)
Процессинг РНК
(1. сплайсинг)
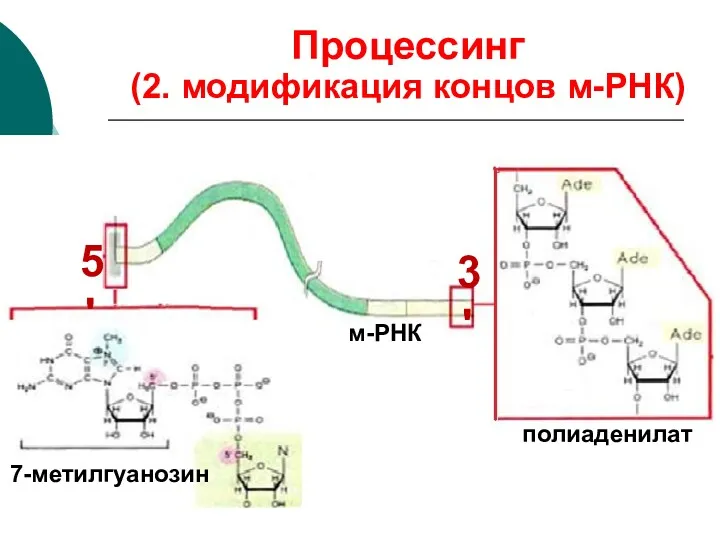
м-РНК
7-метилгуанозин
полиаденилат
5'
3'
Процессинг
(2. модификация концов м-РНК)
м-РНК
7-метилгуанозин
полиаденилат
5'
3'
Процессинг
(2. модификация концов м-РНК)
 Учение академика В.И. Вернадского
Учение академика В.И. Вернадского Проект по биологии В защиту незаслуженно гонимых
Проект по биологии В защиту незаслуженно гонимых Углеводы. Функции углеводов
Углеводы. Функции углеводов Бактерии. Общая характеристика. Взаимоотношения с другими организмами
Бактерии. Общая характеристика. Взаимоотношения с другими организмами Транспорт веществ у растений и животных
Транспорт веществ у растений и животных Химический состав клетки
Химический состав клетки Презентация Как растения показывают погоду и время.
Презентация Как растения показывают погоду и время. Углеводы. Классификация углеводов
Углеводы. Классификация углеводов Игра – акция Зимовка по науке
Игра – акция Зимовка по науке водоросли биология
водоросли биология Предмет и задачи генетики
Предмет и задачи генетики Презентация Витамины биология 8 класс
Презентация Витамины биология 8 класс Растениеводство. Жизненный цикл зерновых хлебов
Растениеводство. Жизненный цикл зерновых хлебов Голонасінні. Ялина європейська
Голонасінні. Ялина європейська Вирусы. Бактерии. Простейшие. Грибы. Лишайники
Вирусы. Бактерии. Простейшие. Грибы. Лишайники Строение клетки. Эндоплазматическая сеть. Комплекс гольджи. Лизосомы. Клеточные включения. Митохондрии. Пластиды
Строение клетки. Эндоплазматическая сеть. Комплекс гольджи. Лизосомы. Клеточные включения. Митохондрии. Пластиды Иммобилизованные ферменты, способы иммобилизации
Иммобилизованные ферменты, способы иммобилизации Строение и жизнедеятельность бактерий
Строение и жизнедеятельность бактерий Водоросли. Разнообразие водорослей
Водоросли. Разнообразие водорослей Царство грибы
Царство грибы Теңіз балдырлары
Теңіз балдырлары Живые клетки. Методы их изучения. Изучение строения клетки растения с помощью микроскопа. 5 класс
Живые клетки. Методы их изучения. Изучение строения клетки растения с помощью микроскопа. 5 класс Белки. Качественный состав белков
Белки. Качественный состав белков Генетические основы селекции собак
Генетические основы селекции собак Они рядом с нами редкие и исчезающие животные Кемеровской области
Они рядом с нами редкие и исчезающие животные Кемеровской области Электрические свойства тканей организма. (Лекция 6)
Электрические свойства тканей организма. (Лекция 6) Введение в общую биологию. (критерии и уровни организации жизни)
Введение в общую биологию. (критерии и уровни организации жизни) Viruses. Procaryotes. Cyanobacteria
Viruses. Procaryotes. Cyanobacteria