- Обмен белков

Содержание
- 2. Аминокислотный состав характеризует пищевую ценность белка . Чем выше содержание незаменимых аминокислот, тем больше пищевая ценность
- 3. Переваривание белков Пищевые белки подвергаются гидролитическому расщеплению под влиянием ПРОТЕОЛИТИЧЕСКИХ ФЕРМЕНТОВ класс – гидролазы, подкласс –
- 4. В ротовой полости белки не подвергаются каким-либо химическим превращениям, т.к. здесь отсутствуют ферменты, действующие на белки
- 5. Под влиянием соляной кислоты ПЕПСИНОГЕН активируется медленно. Более быстро активация происходит под влиянием ПЕПСИНА, т.е. процесс
- 6. СОЛЯНАЯ КИСЛОТА вырабатывается обкладочными клетками слизистой желудка и играет очень важную роль в переваривании белков: 1.
- 7. Образование соляной кислоты в желудке
- 8. Пепсин - это фермент, который является ЭНДОПЕПТИДАЗОЙ, т.е. действует на внутренние ПЕПТИДНЫЕ связи, в образовании которых
- 9. В желудке под действием ПЕПСИНА и ГАСТРИКСИНА сложные белковые молекулы распадаются на высокомолекулярные ПОЛИПЕПТИДЫ, которые поступают
- 10. В кишечнике эти ферменты путем ЧАСТИЧНОГО ПРОТЕОЛИЗА превращаются в активные ферменты: трипсин, химотрипсин, эластаза, карбоксипептидаза. Активация
- 11. Изменение конформации оставшейся части фермента приводит к формированию активного центра и образованию активного трипсина. Образовавшийся трипсин
- 12. ТРИПСИН, ХИМОТРИПСИН, ЭЛАСТАЗА - ЭНДОПЕПТИДАЗЫ. ТРИПСИН разрушает внутренние ПЕПТИДНЫЕ связи, в образовании которых принимают участие лиз
- 13. КАРБОКСИПЕПТИДАЗА разрушает наружные пептидные связи, отщепляя аминокислоты с С-конца полипептидной цепи (ЭКЗОПЕПТИДАЗА)
- 15. Транспорт аминокислот в клетки Аминокислоты, образовавшиеся при гидролизе белков подвергаются всасыванию в кишечнике Аминокислоты поступают в
- 16. Транспорт аминокислот в клетки В настоящее время известно 5 транспортных систем, функционирующих для переноса определенных групп
- 17. Транспорт аминокислот в клетки Одна из специфических транспортных систем функционирует в мозге, кишечнике, почках Она получила
- 18. Этот фермент является гликопротеином и катализирует перенос гаммаглутамильной группы глутатиона на транспортируемую аминокислоту и последующий перенос
- 19. Источники и пути использования аминокислот
- 20. Метаболизм аминокислот Несмотря на то, что почти для каждой аминокислоты выяснены индивидуальные пути обмена, известен ряд
- 21. ТРАНСАМИНИРОВАНИЕ – реакции межмолекулярного переноса аминогруппы от аминокислоты на кетокислоту без промежуточного образования аммиака. Особенности реакций
- 22. Действие аланиновой аминотрансферазы (АЛТ)
- 23. Роль реакций ТРАНСАМИНИРОВАНИЯ: Синтез заменимых аминокислот. При этом происходит перераспределение азота в органах и тканях; Являются
- 24. Реакции ДЕКАРБОКСИЛИРОВАНИЯ – отщепление альфа – карбоксильной группы аминокислот в виде углекислого газа. При этом аминокислоты
- 25. Образование дофамина ДОФА-декарбоксилаза
- 26. Образование серотонина
- 27. Образование гистамина
- 28. Реакции ДЕЗАМИНИРОВАНИЯ – отщепление аминогруппы в виде аммиака ОКИСЛИТЕЛЬНОЕ ДЕЗАМИНИРОВАНИЕ
- 29. НЕПРЯМОЕ ДЕЗАМИНИРОВАНИЕ (ТРАНСДЕЗАМИНИРОВАНИЕ) Если аминокислота не может дезаминироваться прямо, то она может дезаминироваться косвенно с участием
- 30. Это вариант дезаминирования, который протекает в две стадии: а) трансаминирование с участием альфа-кетоглутаровой кислоты; б) окислительное
- 31. БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ РЕАКЦИЙ ДЕЗАМИНИРОВАНИЯ Дезаминирование - путь распада аминокислот Образование аммиака Другой продукт реакций дезаминирования -
- 32. Обезвреживание аммиака Временное обезвреживание (образование амидов дикарбоновых кислот, восстановительное аминирование альфа-кетоглутаровой кислоты) Окончательное обезвреживание аммиака (орнитиновый
- 33. Аминогруппа может присоединяться к аспартату Особенно важны эти реакции в нервной ткани
- 34. Восстановительное аминирование альфа-кетоглутарата
- 35. Окончательное обезвреживание аммиака Орнитиновый цикл (цикл мочевинообразования) (открыт Г.Кребсом) Локализация - матрикс митохондрий циклический процесс
- 36. Первая реакция орнитинового цикла образование карбамоилфосфата путем конденсации NH3, CO2 и АТФ, катализируемое карбамоилфосфатсинтетазой NН3+СО2+2АТФ+Н2О →
- 39. АРГИНАЗА обладает абсолютной специфичностью и содержится только в печени. В составе мочевины содержится два атома азота:
- 40. Образование креатинина КРЕАТИНИН образуется из креатина, который в свою очередь синтезируется в печени из аминокислот, затем
- 41. Креатинфосфат активно синтезируется в покое и распадается при мышечной работе. Это наиболее быстрый способ регенерации АТФ
- 42. Креатинфосфат распадается до креатинина, который является конечным продуктом и выводится с мочой. Количество креатинина прямо пропорционально
- 43. Катаболизм пуриновых нуклеотидов ( образование мочевой кислоты)
- 45. Скачать презентацию

Аминокислотный состав характеризует пищевую ценность белка . Чем выше содержание
Аминокислотный состав характеризует пищевую ценность белка . Чем выше содержание

Переваривание белков
Пищевые белки подвергаются гидролитическому расщеплению под влиянием ПРОТЕОЛИТИЧЕСКИХ ФЕРМЕНТОВ
класс –
Переваривание белков
Пищевые белки подвергаются гидролитическому расщеплению под влиянием ПРОТЕОЛИТИЧЕСКИХ ФЕРМЕНТОВ
класс –

В ротовой полости белки не подвергаются каким-либо химическим превращениям, т.к. здесь
В ротовой полости белки не подвергаются каким-либо химическим превращениям, т.к. здесь

Под влиянием соляной кислоты ПЕПСИНОГЕН активируется медленно. Более быстро активация происходит
Под влиянием соляной кислоты ПЕПСИНОГЕН активируется медленно. Более быстро активация происходит

СОЛЯНАЯ КИСЛОТА вырабатывается обкладочными клетками слизистой желудка и играет очень важную
СОЛЯНАЯ КИСЛОТА вырабатывается обкладочными клетками слизистой желудка и играет очень важную
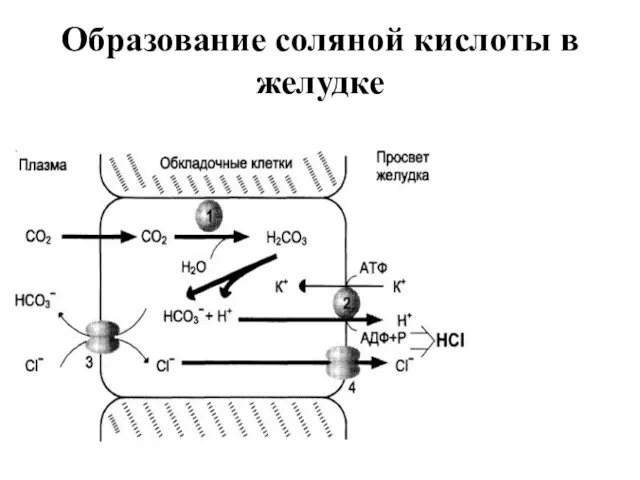
Образование соляной кислоты в желудке
Образование соляной кислоты в желудке

Пепсин - это фермент, который является ЭНДОПЕПТИДАЗОЙ, т.е. действует на внутренние
Пепсин - это фермент, который является ЭНДОПЕПТИДАЗОЙ, т.е. действует на внутренние

В желудке под действием ПЕПСИНА и ГАСТРИКСИНА сложные белковые молекулы распадаются на высокомолекулярные ПОЛИПЕПТИДЫ, которые поступают в тонкий кишечник
В тонком кишечнике эти ПОЛИПЕПТИДЫ
В желудке под действием ПЕПСИНА и ГАСТРИКСИНА сложные белковые молекулы распадаются на высокомолекулярные ПОЛИПЕПТИДЫ, которые поступают в тонкий кишечник
В тонком кишечнике эти ПОЛИПЕПТИДЫ

В кишечнике эти ферменты путем ЧАСТИЧНОГО ПРОТЕОЛИЗА превращаются в активные ферменты:
В кишечнике эти ферменты путем ЧАСТИЧНОГО ПРОТЕОЛИЗА превращаются в активные ферменты:

Изменение конформации оставшейся части фермента приводит к формированию активного центра и
Изменение конформации оставшейся части фермента приводит к формированию активного центра и

ТРИПСИН, ХИМОТРИПСИН, ЭЛАСТАЗА - ЭНДОПЕПТИДАЗЫ.
ТРИПСИН разрушает внутренние ПЕПТИДНЫЕ связи, в образовании
ТРИПСИН, ХИМОТРИПСИН, ЭЛАСТАЗА - ЭНДОПЕПТИДАЗЫ.
ТРИПСИН разрушает внутренние ПЕПТИДНЫЕ связи, в образовании

КАРБОКСИПЕПТИДАЗА разрушает наружные пептидные связи, отщепляя аминокислоты с С-конца полипептидной цепи
КАРБОКСИПЕПТИДАЗА разрушает наружные пептидные связи, отщепляя аминокислоты с С-конца полипептидной цепи

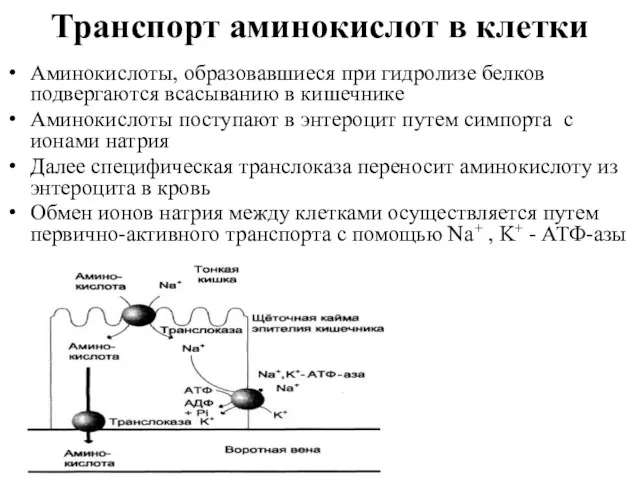
Транспорт аминокислот в клетки
Аминокислоты, образовавшиеся при гидролизе белков подвергаются всасыванию в
Транспорт аминокислот в клетки
Аминокислоты, образовавшиеся при гидролизе белков подвергаются всасыванию в

Транспорт аминокислот в клетки
В настоящее время известно 5 транспортных систем, функционирующих
Транспорт аминокислот в клетки
В настоящее время известно 5 транспортных систем, функционирующих

Транспорт аминокислот в клетки
Одна из специфических транспортных систем функционирует в мозге,
Транспорт аминокислот в клетки
Одна из специфических транспортных систем функционирует в мозге,
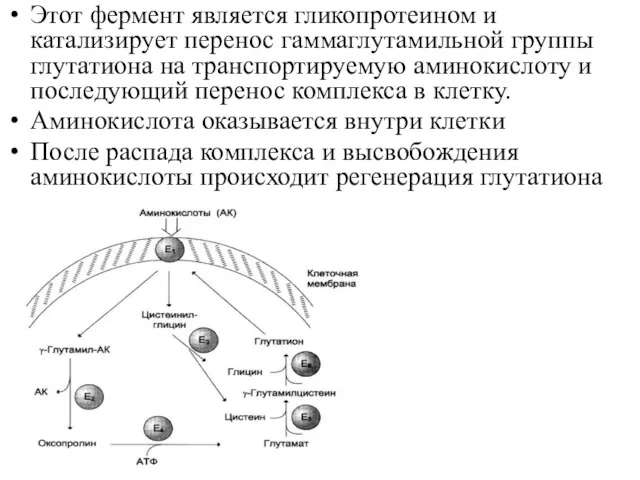
Этот фермент является гликопротеином и катализирует перенос гаммаглутамильной группы глутатиона на
Этот фермент является гликопротеином и катализирует перенос гаммаглутамильной группы глутатиона на
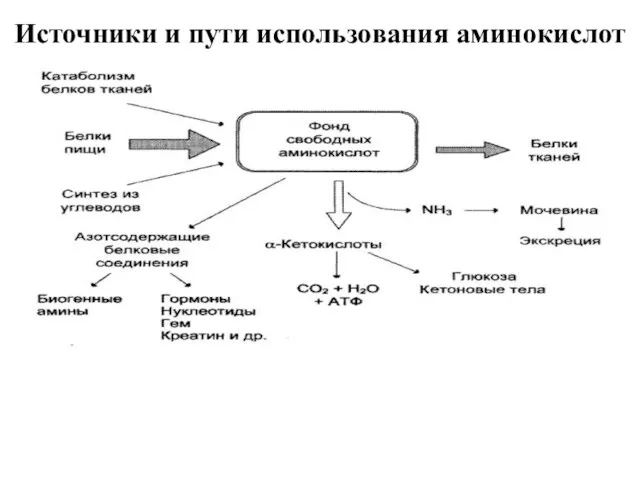
Источники и пути использования аминокислот
Источники и пути использования аминокислот

Метаболизм аминокислот
Несмотря на то, что почти для каждой аминокислоты выяснены индивидуальные
Метаболизм аминокислот
Несмотря на то, что почти для каждой аминокислоты выяснены индивидуальные

ТРАНСАМИНИРОВАНИЕ
– реакции межмолекулярного переноса аминогруппы от аминокислоты на кетокислоту без промежуточного
ТРАНСАМИНИРОВАНИЕ
– реакции межмолекулярного переноса аминогруппы от аминокислоты на кетокислоту без промежуточного
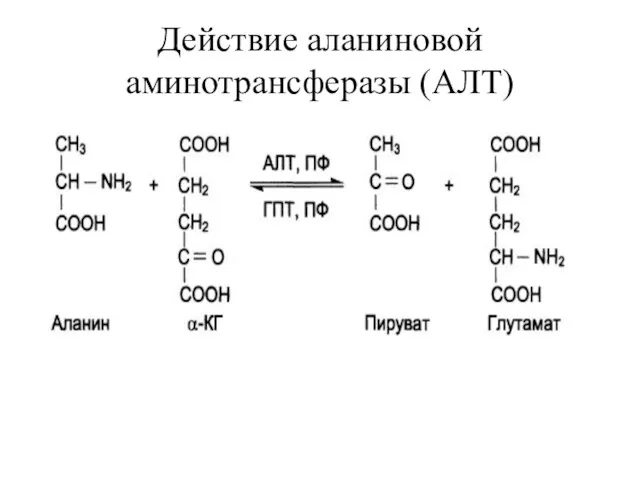
Действие аланиновой аминотрансферазы (АЛТ)
Действие аланиновой аминотрансферазы (АЛТ)

Роль реакций ТРАНСАМИНИРОВАНИЯ:
Синтез заменимых аминокислот.
При этом происходит перераспределение азота в
Роль реакций ТРАНСАМИНИРОВАНИЯ:
Синтез заменимых аминокислот.
При этом происходит перераспределение азота в

Реакции ДЕКАРБОКСИЛИРОВАНИЯ
– отщепление альфа – карбоксильной группы аминокислот в виде углекислого
Реакции ДЕКАРБОКСИЛИРОВАНИЯ
– отщепление альфа – карбоксильной группы аминокислот в виде углекислого
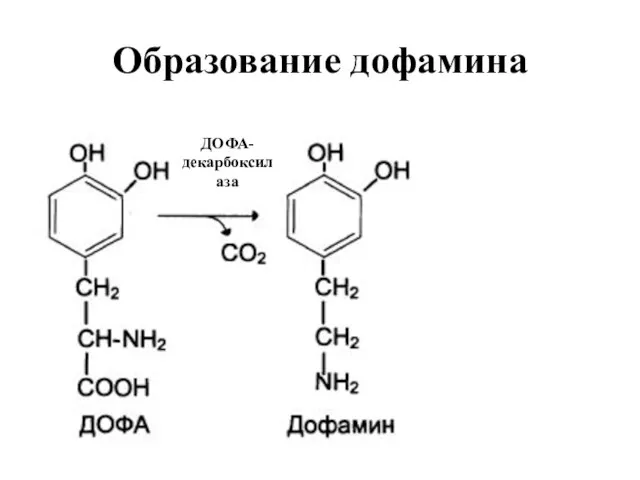
Образование дофамина
ДОФА-декарбоксилаза
Образование дофамина
ДОФА-декарбоксилаза
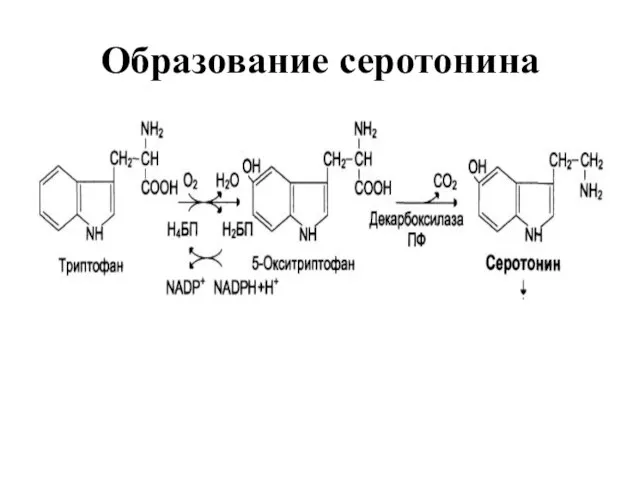
Образование серотонина
Образование серотонина
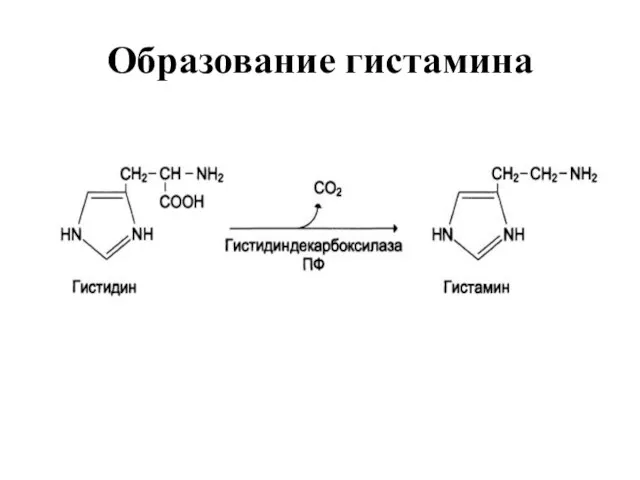
Образование гистамина
Образование гистамина
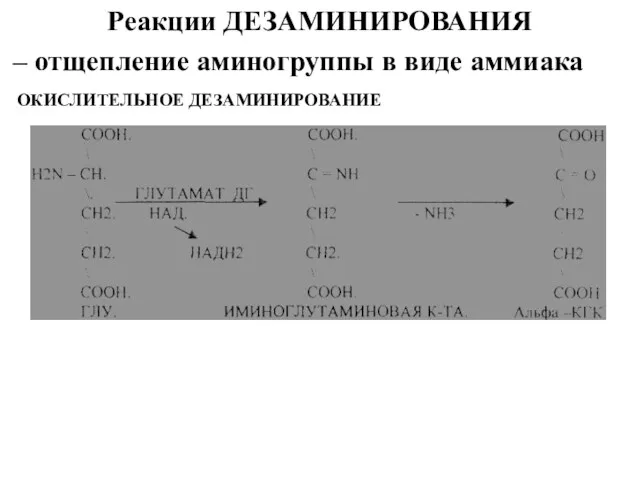
Реакции ДЕЗАМИНИРОВАНИЯ
– отщепление аминогруппы в виде аммиака
ОКИСЛИТЕЛЬНОЕ ДЕЗАМИНИРОВАНИЕ
Реакции ДЕЗАМИНИРОВАНИЯ
– отщепление аминогруппы в виде аммиака
ОКИСЛИТЕЛЬНОЕ ДЕЗАМИНИРОВАНИЕ

НЕПРЯМОЕ ДЕЗАМИНИРОВАНИЕ (ТРАНСДЕЗАМИНИРОВАНИЕ)
Если аминокислота не может дезаминироваться прямо, то она
НЕПРЯМОЕ ДЕЗАМИНИРОВАНИЕ (ТРАНСДЕЗАМИНИРОВАНИЕ)
Если аминокислота не может дезаминироваться прямо, то она

Это вариант дезаминирования, который протекает в две стадии:
а) трансаминирование с
Это вариант дезаминирования, который протекает в две стадии:
а) трансаминирование с

БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ РЕАКЦИЙ ДЕЗАМИНИРОВАНИЯ
Дезаминирование - путь распада аминокислот
Образование аммиака
Другой продукт
БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ РЕАКЦИЙ ДЕЗАМИНИРОВАНИЯ
Дезаминирование - путь распада аминокислот
Образование аммиака
Другой продукт

Обезвреживание аммиака
Временное обезвреживание
(образование амидов дикарбоновых кислот, восстановительное аминирование альфа-кетоглутаровой кислоты)
Окончательное обезвреживание
Обезвреживание аммиака
Временное обезвреживание
(образование амидов дикарбоновых кислот, восстановительное аминирование альфа-кетоглутаровой кислоты)
Окончательное обезвреживание
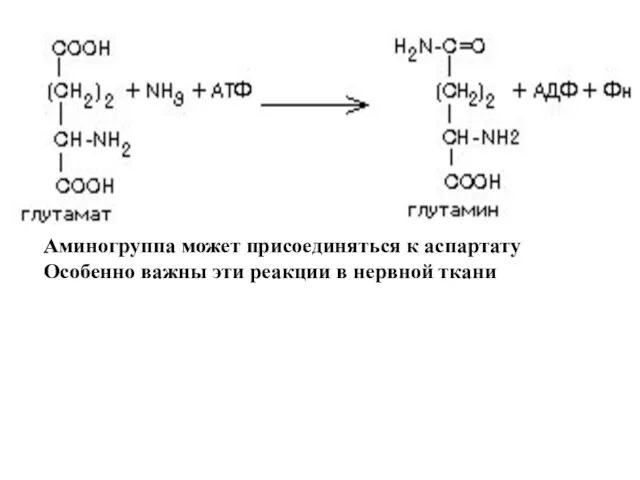
Аминогруппа может присоединяться к аспартату
Особенно важны эти реакции в нервной ткани
Аминогруппа может присоединяться к аспартату
Особенно важны эти реакции в нервной ткани
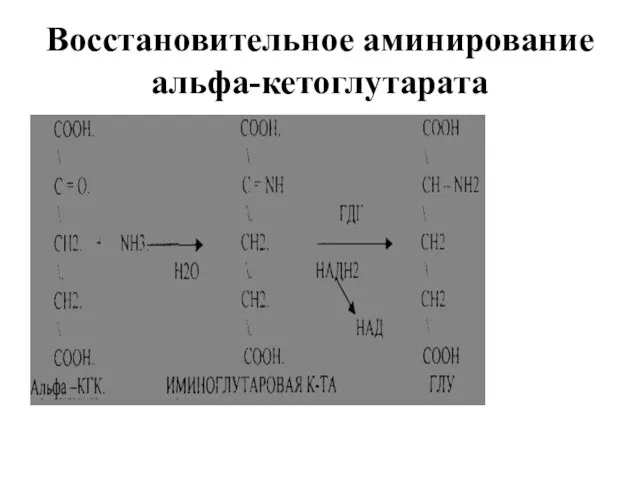
Восстановительное аминирование альфа-кетоглутарата
Восстановительное аминирование альфа-кетоглутарата

Окончательное обезвреживание аммиака
Орнитиновый цикл (цикл мочевинообразования) (открыт Г.Кребсом)
Локализация - матрикс митохондрий
циклический
Окончательное обезвреживание аммиака
Орнитиновый цикл (цикл мочевинообразования) (открыт Г.Кребсом)
Локализация - матрикс митохондрий
циклический

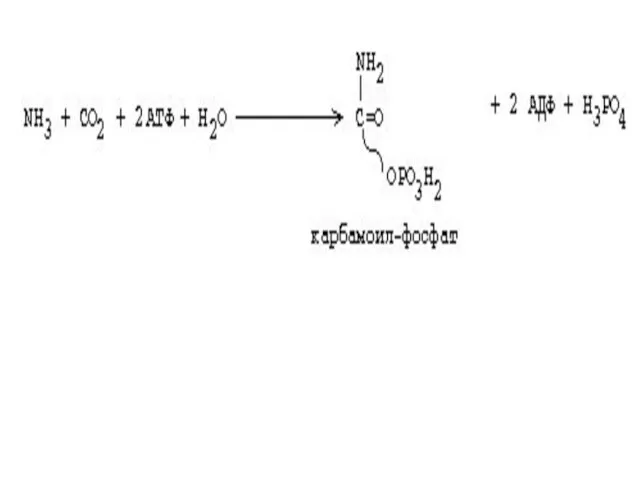
Первая реакция орнитинового цикла
образование карбамоилфосфата путем конденсации NH3, CO2 и АТФ,
Первая реакция орнитинового цикла
образование карбамоилфосфата путем конденсации NH3, CO2 и АТФ,
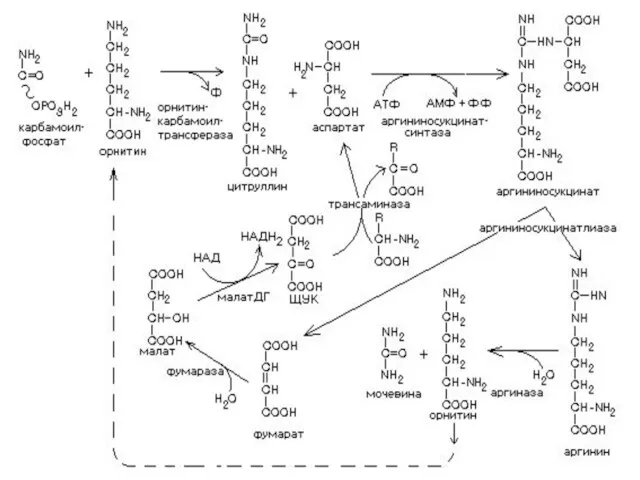

АРГИНАЗА обладает абсолютной специфичностью и содержится только в печени.
В составе
АРГИНАЗА обладает абсолютной специфичностью и содержится только в печени.
В составе

Образование креатинина
КРЕАТИНИН образуется из креатина, который в свою очередь синтезируется в
Образование креатинина
КРЕАТИНИН образуется из креатина, который в свою очередь синтезируется в

Креатинфосфат активно синтезируется в покое и распадается при мышечной работе. Это
Креатинфосфат активно синтезируется в покое и распадается при мышечной работе. Это

Креатинфосфат распадается до креатинина, который является конечным продуктом и выводится с
Креатинфосфат распадается до креатинина, который является конечным продуктом и выводится с
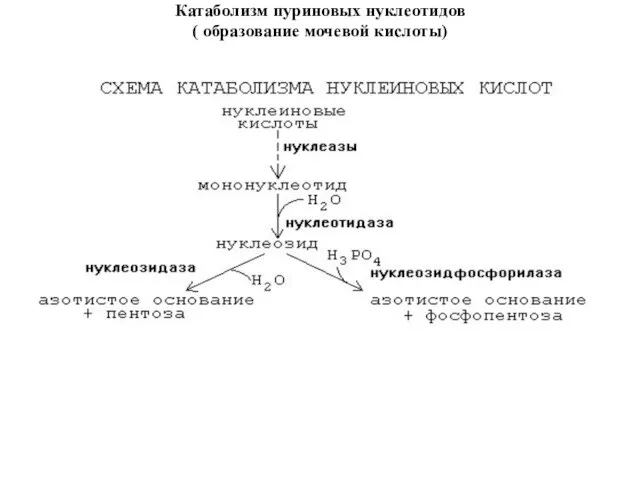
Катаболизм пуриновых нуклеотидов
( образование мочевой кислоты)
Катаболизм пуриновых нуклеотидов
( образование мочевой кислоты)
 Человек – предмет изучения анатомии и физиологии
Человек – предмет изучения анатомии и физиологии Обмен веществ. Метаболизм Катаболизм (энергетический обмен). Анаболизм (пластический обмен)
Обмен веществ. Метаболизм Катаболизм (энергетический обмен). Анаболизм (пластический обмен) Пищеварительная система человека
Пищеварительная система человека Фитотерапия - лечение лекарственными травами. 1 класс
Фитотерапия - лечение лекарственными травами. 1 класс Тип Иглокожие
Тип Иглокожие Азотное питание растений
Азотное питание растений Изменчивость и патология
Изменчивость и патология Основы молекулярной биологии. Поток генетической информации: клеточный уровень. Нуклеиновые кислоты: строение, свойства, функции
Основы молекулярной биологии. Поток генетической информации: клеточный уровень. Нуклеиновые кислоты: строение, свойства, функции Викторина на тему Витамины
Викторина на тему Витамины Animals zoo
Animals zoo Обонятельный и вкусовой анализаторы
Обонятельный и вкусовой анализаторы Характеристика класса птицы. Внешнее строение птиц
Характеристика класса птицы. Внешнее строение птиц Метаботропные рецепторы
Метаботропные рецепторы Плоды. Значение и разнообразие плодов
Плоды. Значение и разнообразие плодов Секвенирование. Нуклеотиды
Секвенирование. Нуклеотиды Прокариотическая клетка
Прокариотическая клетка Пищеварение 1.1
Пищеварение 1.1 Размножение, гаметогенез
Размножение, гаметогенез Витамины и их роль в обмене веществ
Витамины и их роль в обмене веществ Амурский тигр
Амурский тигр Многообразие живых организмов
Многообразие живых организмов Система CRISPR/Cas9
Система CRISPR/Cas9 Жердегі биологиялық эволюцияның кезеңдері мен негіздері
Жердегі биологиялық эволюцияның кезеңдері мен негіздері Филогенетическое разнообразие грибоподобных организмов. Разнообразие. Систематика. Жизненные циклы
Филогенетическое разнообразие грибоподобных организмов. Разнообразие. Систематика. Жизненные циклы Презентация к уроку биологии Тип инфузории.Происхождение простейших 7 класс
Презентация к уроку биологии Тип инфузории.Происхождение простейших 7 класс Птицы Алтайского края
Птицы Алтайского края Артерии большого круга кровообращения
Артерии большого круга кровообращения Наследственная изменчивость. Мутации
Наследственная изменчивость. Мутации