- Обмен липидов. Регуляция обмена липидов. Регуляция окисления и синтеза ЖК. (Тема 4)

Содержание
- 2. ПЛАН ЛЕКЦИИ 1. Регуляция обмена липидов. Регуляция окисления и синтеза ЖК. 2. Метаболизм кетоновых тел в
- 3. Регуляция обмена липидов Интенсивность обмена липидов в тканях зависит от: а) поступления липидов с пищей; б)
- 4. . Вместе с тем, растительные масла, содержащие ненасыщенные ЖК напротив, способствуют мобилизации липидов. Наличие липотропных факторов
- 5. Нервно-гормональная регуляция липидного обмена Стимуляторы тканевой липазы - адреналин, норадреналин, глюкагон, тироксин, адренокортикотропный гормон (активируют аденилатциклазу)
- 6. . В такой форме данный рецептор способен активировать аденилатциклазу, что стимулирует образование ц-АМФ и АТФ. Далее
- 8. Окисление и синтез ЖК регулируется: а) разделением этих противоположных процессов в пространстве (окисление ЖК в митохондриях,
- 9. БИОСИНТЕЗ ТРИГЛИЦЕРИДОВ Синтез триглицеридов происходит из глицерина и жирных кислот (главным образом стеариновой, пальмитиновой и олеиновой).
- 10. В почках, а также в стенке кишечника, где активность фермента глицеролкиназы высока, глицерин фосфорилируется за счет
- 11. В жировой ткани и мышцах вследствие очень низкой активности глицеролкиназы образование глицерол-3-фосфата в основном связано с
- 12. Последний в присутствии цитоплазматической глицерол-3-фосфатдегидрогеназы способен превращаться в глицерол-3-фосфат:
- 13. Если содержание глюкозы в жировой ткани понижено (например, при голодании), то образуется лишь незначительное количество глицерол-3-фосфата
- 14. Глицерол-3-фосфат последовательно ацилируется двумя молекулами КоА-производного жирной кислоты. В результате образуется фосфатидная кислота (фосфатидат):
- 15. Далее фосфатидная кислота гидролизуется фосфатидат-фосфогидролазой до 1,2-диглицерида (1,2-диацилглицерола):
- 16. Затем 1,2-диглицерид ацилируется третьей молекулой ацил-КоА и превращается в триглицерид (триацилглицерол). Эта реакция катализируется диацилглицерол-ацилтрансферазой:
- 17. Синтез триглицеридов (триацилглицеролов) в тканях происходит с учетом двух путей образования глицерол-3-фосфата и возможности синтеза триглицеридов
- 18. БИОСИНТЕЗ ХОЛЕСТЕРИНА В синтезе холестерина можно выделить три основные стадии: I – превращение активного ацетата в
- 19. Превращение активного ацетата в мевалоновую кислоту Начальным этапом синтеза мевалоновой кислоты из ацетил-КоА является образование ацетоацетил-КоА
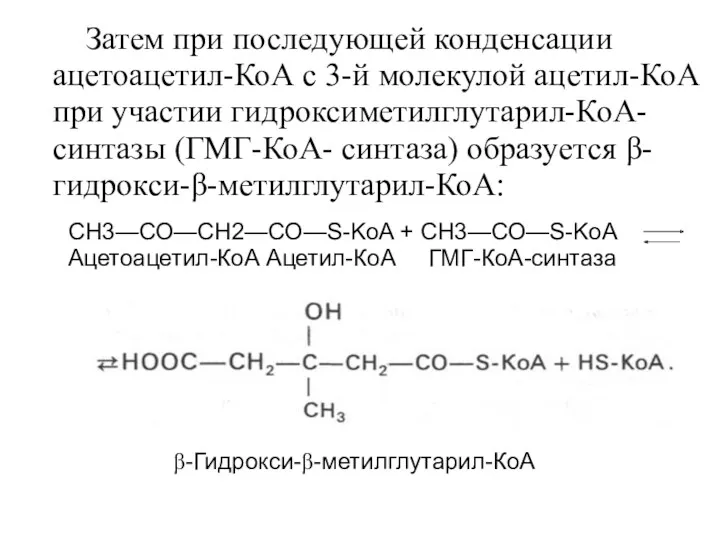
- 20. Затем при последующей конденсации ацетоацетил-КоА с 3-й молекулой ацетил-КоА при участии гидроксиметилглутарил-КоА-синтазы (ГМГ-КоА- синтаза) образуется β-гидрокси-β-метилглутарил-КоА:
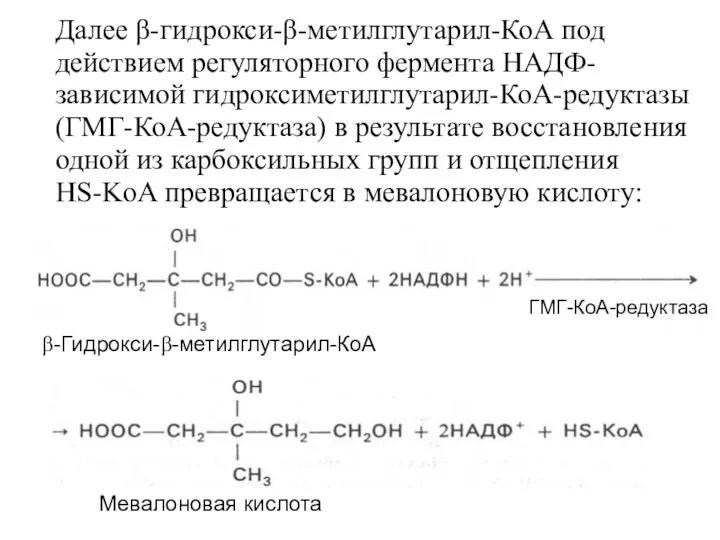
- 21. Далее β-гидрокси-β-метилглутарил-КоА под действием регуляторного фермента НАДФ-зависимой гидроксиметилглутарил-КоА-редуктазы (ГМГ-КоА-редуктаза) в результате восстановления одной из карбоксильных групп
- 22. На II стадии синтеза холестерина мевалоновая кислота превращается в сквален. Реакции II стадии начинаются с фосфорилирования
- 23. 5-пирофосфомевалоновая кислота в результате последующего фосфорилирования третичной гидроксильной группы образует нестабильный промежуточный продукт – 3-фосфо-5-пирофосфомевалоновую кислоту,
- 24. Диметилаллилпирофосфат Изопентенилпирофосфат
- 25. Затем оба изомерных изопентенилпирофосфата (диметилаллилпирофосфат и изопентенилпирофосфат) конденсируются с высвобождением пирофосфата и образованием геранилпирофосфата: Геранилпирофосфат
- 26. К геранилпирофосфату вновь присоединяется изопентенилпирофосфат. В результате этой реакции образуется фарнезилпирофосфат: Фарнезилпирофосфат (С15)
- 27. В заключительной реакции данной стадии в результате НАДФН-зависимой восстановительной конденсации 2 молекул фарнезилпирофосфата образуется сквален: Сквален
- 28. На III стадии биосинтеза холестерина сквален под влиянием сквален-оксидоциклазы циклизируется с образованием ланостерина. Дальнейший процесс превращения
- 29. Ланостерин(С30) Холестерин (С27)
- 30. Начиная со сквалена, все промежуточные продукты биосинтеза холестерина (включая и холестерин) нерастворимы в водной среде. Поэтому
- 32. Скачать презентацию

ПЛАН ЛЕКЦИИ
1. Регуляция обмена липидов. Регуляция окисления и синтеза ЖК.
2. Метаболизм
ПЛАН ЛЕКЦИИ
1. Регуляция обмена липидов. Регуляция окисления и синтеза ЖК.
2. Метаболизм

Регуляция обмена липидов
Интенсивность обмена липидов в тканях зависит от:
а) поступления липидов
Регуляция обмена липидов
Интенсивность обмена липидов в тканях зависит от:
а) поступления липидов

.
Вместе с тем, растительные масла, содержащие ненасыщенные ЖК напротив, способствуют мобилизации
.
Вместе с тем, растительные масла, содержащие ненасыщенные ЖК напротив, способствуют мобилизации

Нервно-гормональная регуляция липидного обмена
Стимуляторы тканевой липазы - адреналин, норадреналин, глюкагон, тироксин,
Нервно-гормональная регуляция липидного обмена
Стимуляторы тканевой липазы - адреналин, норадреналин, глюкагон, тироксин,

.
В такой форме данный рецептор способен активировать аденилатциклазу, что стимулирует образование
.
В такой форме данный рецептор способен активировать аденилатциклазу, что стимулирует образование
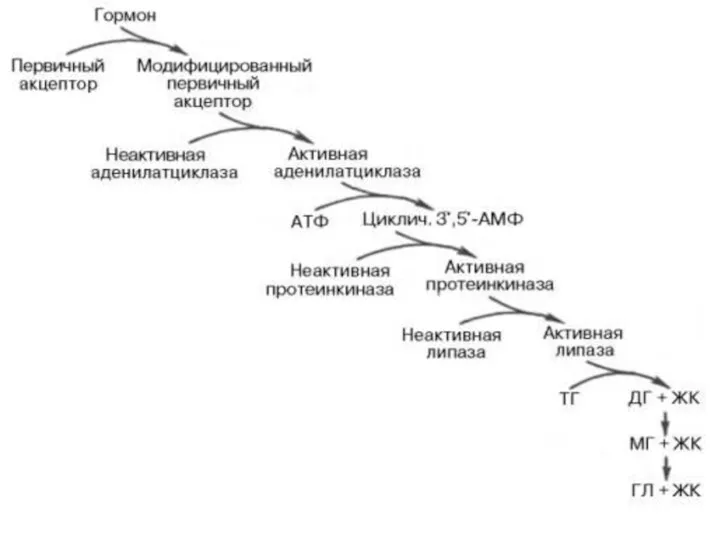

Окисление и синтез ЖК регулируется:
а) разделением этих противоположных процессов в пространстве
Окисление и синтез ЖК регулируется:
а) разделением этих противоположных процессов в пространстве

БИОСИНТЕЗ ТРИГЛИЦЕРИДОВ
Синтез триглицеридов происходит из глицерина и жирных кислот
(главным образом стеариновой,
БИОСИНТЕЗ ТРИГЛИЦЕРИДОВ
Синтез триглицеридов происходит из глицерина и жирных кислот
(главным образом стеариновой,

В почках, а также в стенке кишечника, где активность фермента
глицеролкиназы высока,
В почках, а также в стенке кишечника, где активность фермента
глицеролкиназы высока,

В жировой ткани и мышцах вследствие очень низкой активности
глицеролкиназы образование глицерол-3-фосфата
В жировой ткани и мышцах вследствие очень низкой активности
глицеролкиназы образование глицерол-3-фосфата
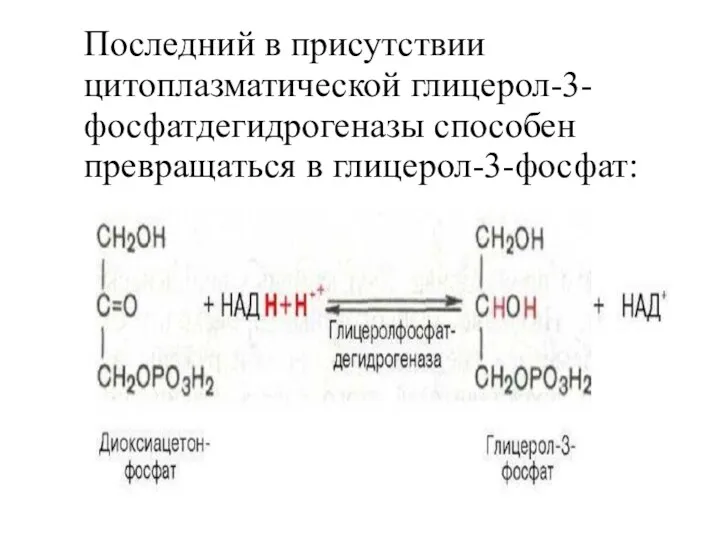
Последний в присутствии цитоплазматической глицерол-3-фосфатдегидрогеназы способен превращаться в глицерол-3-фосфат:
Последний в присутствии цитоплазматической глицерол-3-фосфатдегидрогеназы способен превращаться в глицерол-3-фосфат:

Если содержание глюкозы в жировой ткани понижено (например, при голодании), то
Если содержание глюкозы в жировой ткани понижено (например, при голодании), то
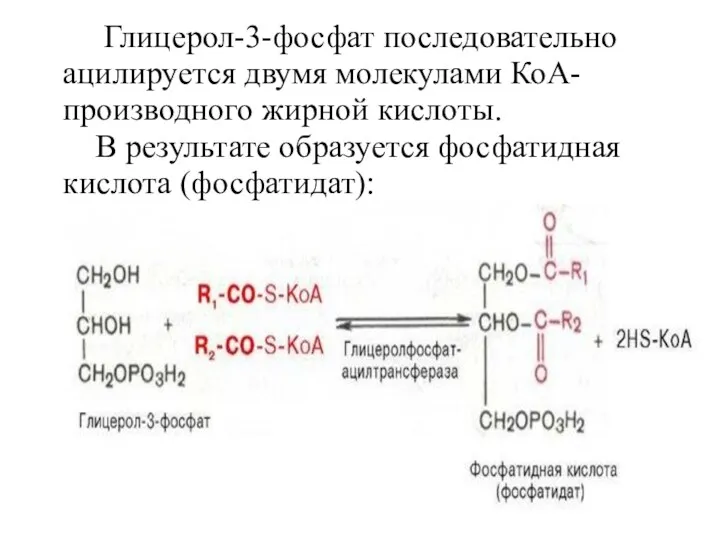
Глицерол-3-фосфат последовательно ацилируется двумя молекулами КоА-производного жирной кислоты.
В результате
Глицерол-3-фосфат последовательно ацилируется двумя молекулами КоА-производного жирной кислоты.
В результате
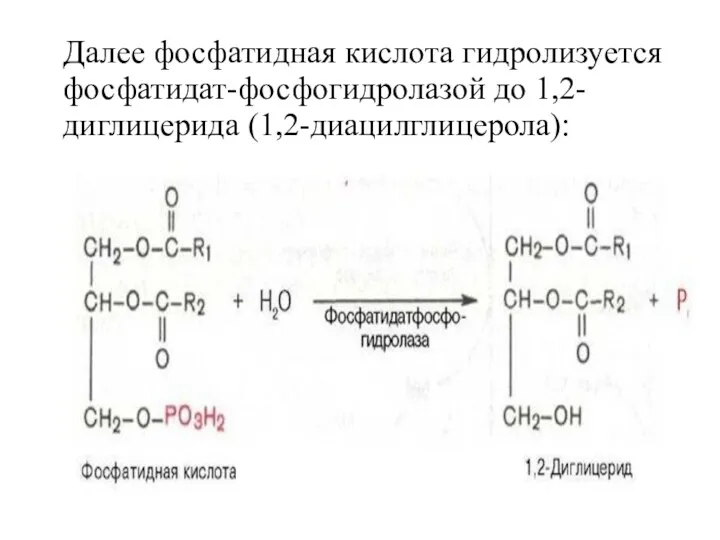
Далее фосфатидная кислота гидролизуется фосфатидат-фосфогидролазой до 1,2-диглицерида (1,2-диацилглицерола):
Далее фосфатидная кислота гидролизуется фосфатидат-фосфогидролазой до 1,2-диглицерида (1,2-диацилглицерола):

Затем 1,2-диглицерид ацилируется третьей молекулой ацил-КоА и превращается в триглицерид (триацилглицерол).
Затем 1,2-диглицерид ацилируется третьей молекулой ацил-КоА и превращается в триглицерид (триацилглицерол).

Синтез триглицеридов (триацилглицеролов) в тканях происходит с учетом двух путей
Синтез триглицеридов (триацилглицеролов) в тканях происходит с учетом двух путей

БИОСИНТЕЗ ХОЛЕСТЕРИНА
В синтезе холестерина можно выделить три основные стадии: I –
БИОСИНТЕЗ ХОЛЕСТЕРИНА
В синтезе холестерина можно выделить три основные стадии: I –

Превращение активного ацетата в мевалоновую кислоту
Начальным этапом синтеза мевалоновой кислоты
Превращение активного ацетата в мевалоновую кислоту
Начальным этапом синтеза мевалоновой кислоты
Затем при последующей конденсации ацетоацетил-КоА с 3-й молекулой ацетил-КоА при
Затем при последующей конденсации ацетоацетил-КоА с 3-й молекулой ацетил-КоА при
Далее β-гидрокси-β-метилглутарил-КоА под действием регуляторного фермента НАДФ-зависимой гидроксиметилглутарил-КоА-редуктазы (ГМГ-КоА-редуктаза) в результате
Далее β-гидрокси-β-метилглутарил-КоА под действием регуляторного фермента НАДФ-зависимой гидроксиметилглутарил-КоА-редуктазы (ГМГ-КоА-редуктаза) в результате

На II стадии синтеза холестерина мевалоновая кислота превращается в сквален.
На II стадии синтеза холестерина мевалоновая кислота превращается в сквален.

5-пирофосфомевалоновая кислота в результате последующего фосфорилирования третичной гидроксильной группы образует нестабильный
промежуточный
5-пирофосфомевалоновая кислота в результате последующего фосфорилирования третичной гидроксильной группы образует нестабильный
промежуточный
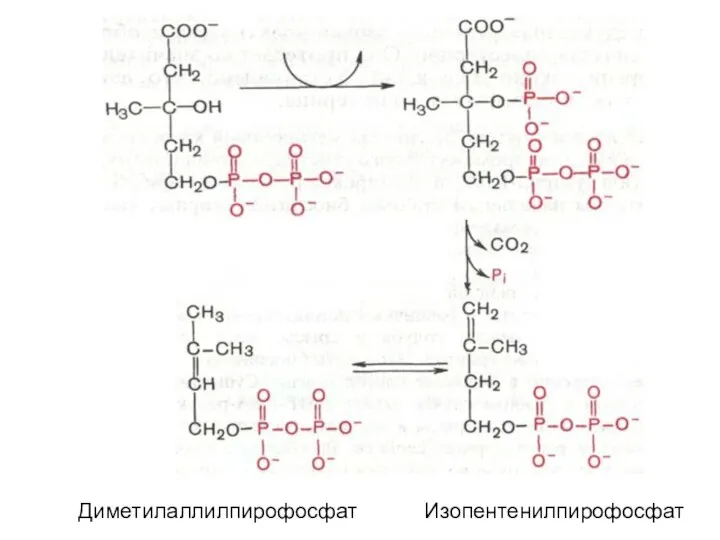
Диметилаллилпирофосфат Изопентенилпирофосфат
Диметилаллилпирофосфат Изопентенилпирофосфат
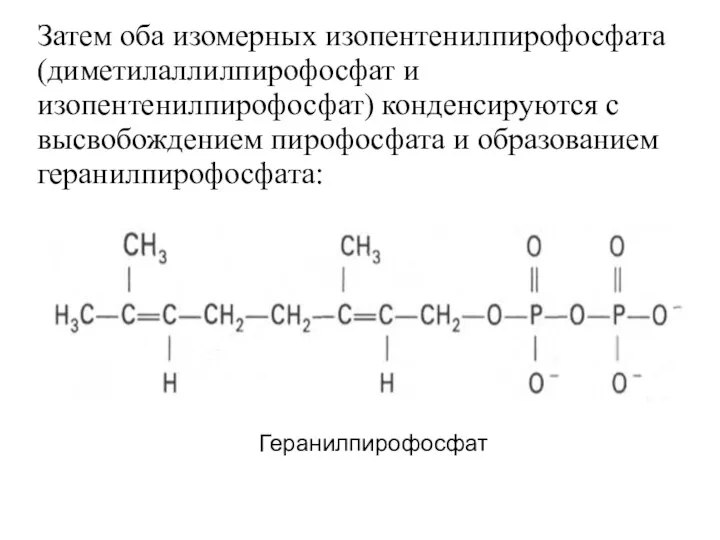
Затем оба изомерных изопентенилпирофосфата (диметилаллилпирофосфат и изопентенилпирофосфат) конденсируются с высвобождением пирофосфата
Затем оба изомерных изопентенилпирофосфата (диметилаллилпирофосфат и изопентенилпирофосфат) конденсируются с высвобождением пирофосфата
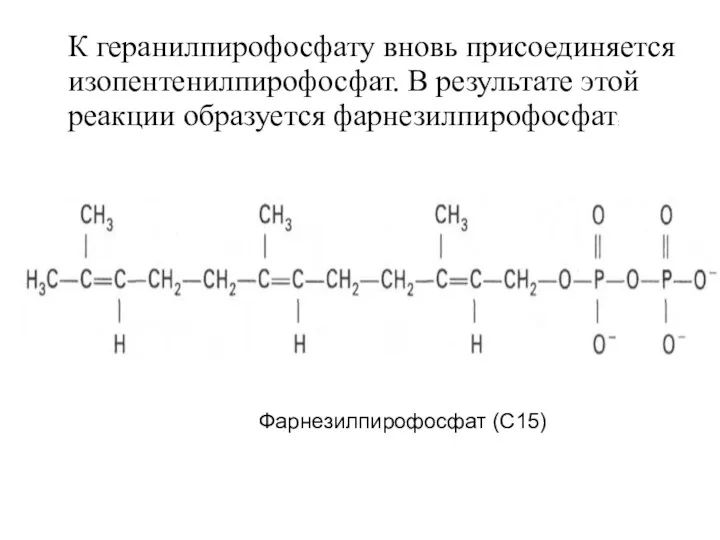
К геранилпирофосфату вновь присоединяется изопентенилпирофосфат. В результате этой реакции образуется фарнезилпирофосфат:
Фарнезилпирофосфат
К геранилпирофосфату вновь присоединяется изопентенилпирофосфат. В результате этой реакции образуется фарнезилпирофосфат:
Фарнезилпирофосфат
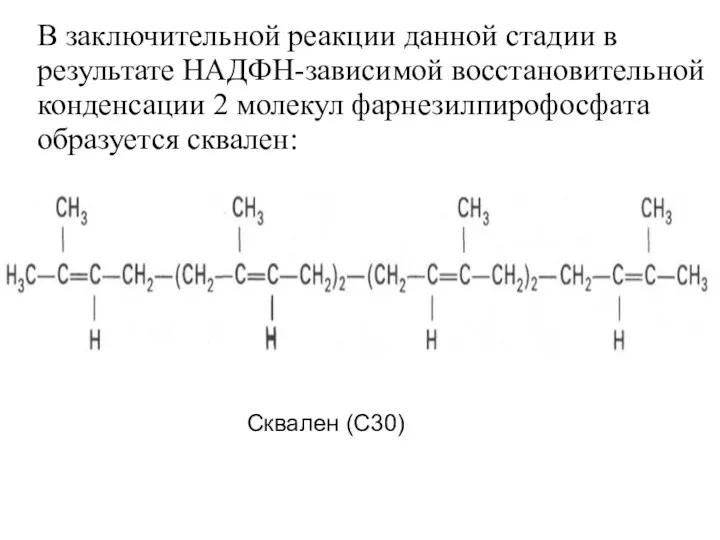
В заключительной реакции данной стадии в результате НАДФН-зависимой восстановительной конденсации 2
В заключительной реакции данной стадии в результате НАДФН-зависимой восстановительной конденсации 2

На III стадии биосинтеза холестерина сквален под влиянием сквален-оксидоциклазы циклизируется
На III стадии биосинтеза холестерина сквален под влиянием сквален-оксидоциклазы циклизируется
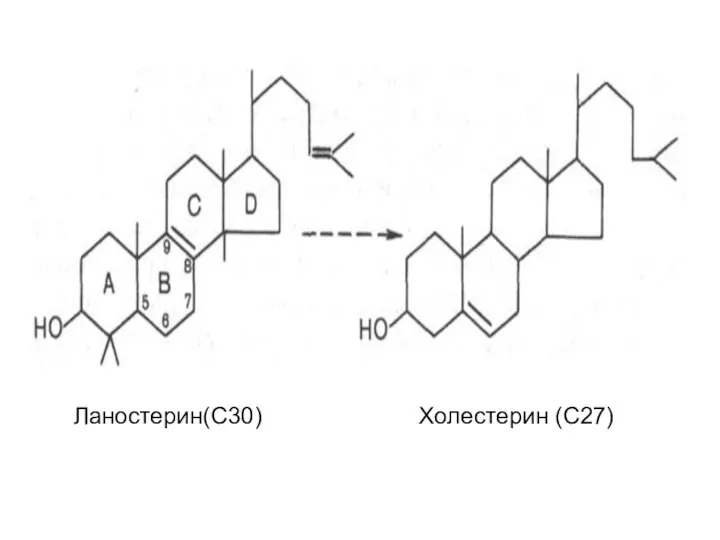
Ланостерин(С30) Холестерин (С27)
Ланостерин(С30) Холестерин (С27)

Начиная со сквалена, все промежуточные продукты биосинтеза холестерина (включая и холестерин)
Начиная со сквалена, все промежуточные продукты биосинтеза холестерина (включая и холестерин)
 Лимфатическая система
Лимфатическая система Красные водоросли
Красные водоросли Органы пищеварения
Органы пищеварения Рациональное питание - залог здоровья
Рациональное питание - залог здоровья Красота природы. Жизнь в заповеднике (фотографии)
Красота природы. Жизнь в заповеднике (фотографии) Цианобактерии. Происхождение
Цианобактерии. Происхождение Ощущение как психологический процесс
Ощущение как психологический процесс Всё о кошках
Всё о кошках Микроорганизмдер, антибиотиктер және биологиялық активті заттарды түзушілер
Микроорганизмдер, антибиотиктер және биологиялық активті заттарды түзушілер Закономерности артериальной системы
Закономерности артериальной системы Мужская половая система
Мужская половая система Лишайники- биоиндикаторы загрязнения воздуха
Лишайники- биоиндикаторы загрязнения воздуха Генетика как наука
Генетика как наука Насекомые полезные и вредные
Насекомые полезные и вредные Хрящевые рыбы
Хрящевые рыбы Презентация по биологии Тип Кишечнополостные
Презентация по биологии Тип Кишечнополостные Грибы, их значение в природе и жизни человека
Грибы, их значение в природе и жизни человека Интегративные функции промежуточного мозга и подкорковых ядер
Интегративные функции промежуточного мозга и подкорковых ядер Мышечная система. Строение и функции мышц
Мышечная система. Строение и функции мышц Змiни в природi взимку. Урок №49. Я досліджую світ
Змiни в природi взимку. Урок №49. Я досліджую світ Функциональная анатомия вегетативной нервной системы. Симпатическая часть ВНС (лекция № 23)
Функциональная анатомия вегетативной нервной системы. Симпатическая часть ВНС (лекция № 23) Общая характеристика типа Членистоногие
Общая характеристика типа Членистоногие Общие закономерности развития. Биогенетический закон
Общие закономерности развития. Биогенетический закон Методы изучения живой природы. Измерение. 5 урок. 5 класс
Методы изучения живой природы. Измерение. 5 урок. 5 класс Вегетативне розмноження рослин
Вегетативне розмноження рослин Загадки о животных и растениях (2 класс)
Загадки о животных и растениях (2 класс) Видоизменение корней у растений
Видоизменение корней у растений Дельфины. Общие сведения о дельфинах
Дельфины. Общие сведения о дельфинах