- Обмен углеводов

Содержание
- 2. Понятие об углеводах, распространение в природе и значение для организма животных. Переваривание и всасывание углеводов. Катаболизм
- 3. Углеводы – это альдегидо- и кетонопроизводные многоатомных спиртов, их циклические формы и продукты их конденсации. Энергетическая
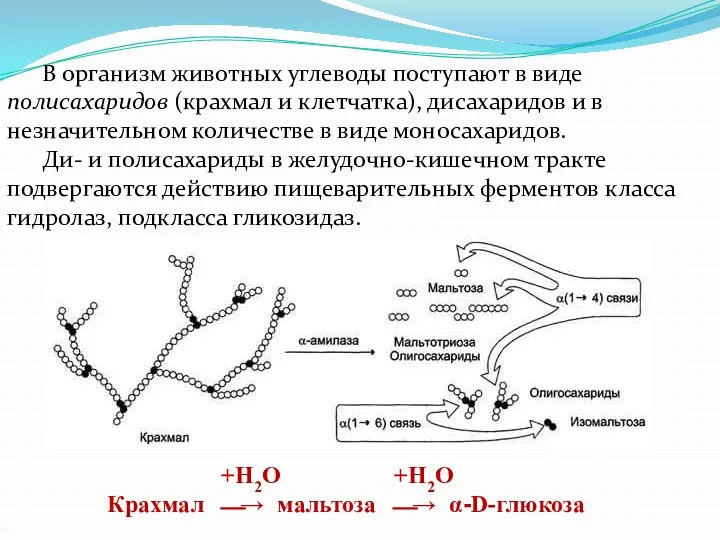
- 4. В организм животных углеводы поступают в виде полисахаридов (крахмал и клетчатка), дисахаридов и в незначительном количестве
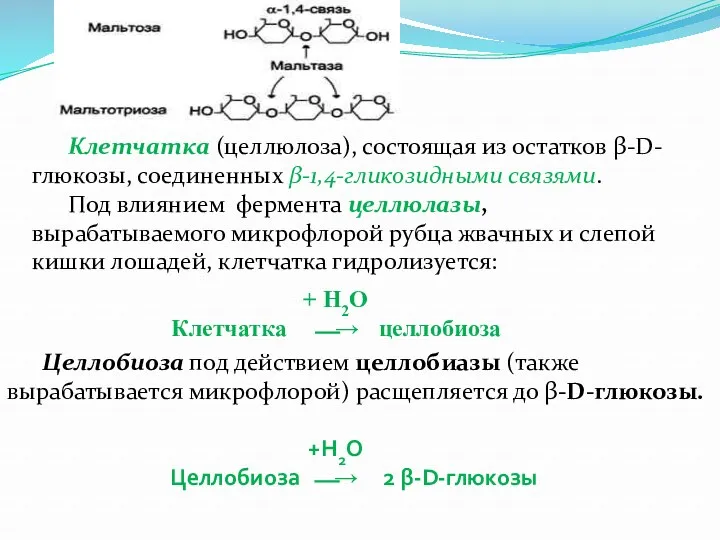
- 5. Клетчатка (целлюлоза), состоящая из остатков β-D-глюкозы, соединенных β-1,4-гликозидными связями. Под влиянием фермента целлюлазы, вырабатываемого микрофлорой рубца
- 6. В дальнейшем часть β-D-глюкозы под действием ферментов микроорганизмов подвергается различным видам брожения. При этом образуются летучие
- 7. Молочный сахар лактоза расщепляется под действием лактазы: + Н2О Лактоза ⎯→ β-D-галактоза + α-D-глюкоза Сахароза подвергается
- 8. Переваривание углеводов
- 9. Всасывание углеводов Всасывание моносахаридов представляет собой сложный биохимический процесс их транспорта через мембраны клеток тонкого кишечника.
- 11. Поступление углеводов в энтероциты.
- 12. Метаболизм глюкозо-6-фосфата
- 13. Катаболизм глюкозы Анаэробный этап окисления пирувата (образование лактата) Аэробный этап окисления пирувата до СО2 и Н2О
- 14. Гликолиз - анаэробное превращение глюкозы, локализующееся в цитозоле и включающее два этапа из 10 ферментативных реакций.
- 15. В четвертой реакции фруктозо-1,6-дифосфат разрезается пополам фруктозо-1,6-дифосфат-альдолазой с образованием двух фосфорилированных триоз-изомеров – альдозы глицеральдегида (ГАФ)
- 16. Второй этап гликолиза – это освобождение энергии, содержащейся в глицеральдегидфосфате, и запасание ее в форме АТФ.
- 17. Восьмая реакция – синтезированный в предыдущей реакции 3-фосфоглицерат под влиянием фосфоглицератмутазы изомеризуется в 2-фосфоглицерат. Девятая реакция
- 18. Метаболизм пирувата в анаэорбных условиях С6Н1206 + 2 Н3Р04 + 2 АДФ = 2 С3Н6О3 +
- 19. Для стабильного протекания гликолиза в бескислородных условиях необходимы ряд простых условий: 1) поступление глюкозы, 2) наличие
- 20. Баланс АТФ при анаэробном распаде глюкозы Анаэробный гликолиз по сравнению с аэробным менее эффективен. В этом
- 21. Биологическая значение анаэробного превращения глюкозы Энергетическое. Анаэробное превращение глюкозы – источник АТФ для биосинтетических процессов (идущих
- 22. Это снижение потребления глюкозы и прекращение продукции молочной кислоты клеткой в присутствии кислорода. Биохимический механизм эффекта
- 23. Глицеролфосфатный челночный механизм. Челночные системы. Ключевыми ферментами глицеролфосфатного челнока являются изоферменты глицерол-3-фосфат-дегидрогеназы – цитоплазматический и митохондриальный.
- 24. . Ключевыми ферментами этого челнока являются изоферменты малатдегидрогеназы – цитоплазматический и митохондриальный. Он является распространенным по
- 25. Из цитозоля пировиноградная кислота симпортом с ионами Н+ движется по протонному градиент в митохондрии. В митохондриях
- 26. Баланс АТФ при аэробном распаде до СО2 и Н2О
- 27. Физиологический смысл гликолиза 1. У аэробных организмов гликолиз является первым, подготовительным этапом дыхания. В процессе гликолиза
- 28. Это синтез глюкозы из неуглеводных компонентов: лактата, пирувата, глицерола, кетокислот цикла Кребса и других кетокислот, из
- 29. На этом этапе глюконеогенеза работают два ключевых фермента – в митохондриях пируваткарбоксилаза и в цитозоле фосфоенолпируват-карбоксикиназа.
- 30. Из цитозоля пировиноградная кислота симпортом с ионами Н+ движется по протонному градиент в митохондрии. 2. Далее
- 31. Обход третьей реакции гликолиза Второе препятствие на пути синтеза глюкозы – фосфофруктокиназная реакция – преодолевается с
- 32. Обход первой реакции гликолиза Последняя реакция катализируется глюкозо-6-фосфатазой. Она имеется только в печени и почках, следовательно,
- 33. Гликолиз и глюконеогенез дополняют друг друга Поскольку гликолитическое окисление глюкозы является способом получения энергии и в
- 34. Регуляция гликолиза и глюконеогенеза
- 35. Гликоген - разветвлённый гомополимер глюкозы, в котором остатки глюкозы соединены в линейных участках α-1,4-гликозидной связью. В
- 36. Синтез гликогена (гликогеногенез)
- 37. Глюкоза, поступающая в клетку, фосфорилируется при участии АТФ (реакция 1). Затем глюкозо-6-фосфат в ходе обратимой реакции
- 38. Гликогенолиз
- 39. Распад гликогена или его мобилизация происходят в ответ на повышение потребности организма в глюкозе. Гликоген печени
- 40. Считают, что перенос трёх остатков глюкозы и удаление мономера из точки ветвления (реакции 2 и 3)
- 41. Сравнение этих процессов позволяет сделать следующие выводы: а) синтез и распад гликогена протекают по разным метаболическими
- 43. Скачать презентацию

Понятие об углеводах, распространение в природе и значение для организма животных.
Переваривание
Понятие об углеводах, распространение в природе и значение для организма животных.
Переваривание

Углеводы – это альдегидо- и кетонопроизводные многоатомных спиртов, их циклические формы
Углеводы – это альдегидо- и кетонопроизводные многоатомных спиртов, их циклические формы
В организм животных углеводы поступают в виде полисахаридов (крахмал и клетчатка),
В организм животных углеводы поступают в виде полисахаридов (крахмал и клетчатка),
Клетчатка (целлюлоза), состоящая из остатков β-D-глюкозы, соединенных β-1,4-гликозидными связями.
Под влиянием фермента
Клетчатка (целлюлоза), состоящая из остатков β-D-глюкозы, соединенных β-1,4-гликозидными связями.
Под влиянием фермента

В дальнейшем часть β-D-глюкозы под действием ферментов микроорганизмов подвергается различным видам
В дальнейшем часть β-D-глюкозы под действием ферментов микроорганизмов подвергается различным видам

Молочный сахар лактоза расщепляется под действием лактазы:
+ Н2О
Лактоза ⎯→ β-D-галактоза
Молочный сахар лактоза расщепляется под действием лактазы:
+ Н2О
Лактоза ⎯→ β-D-галактоза
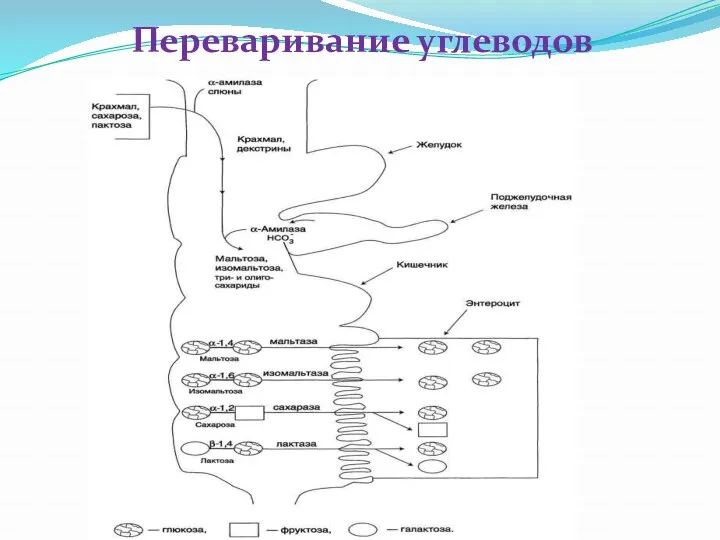
Переваривание углеводов
Переваривание углеводов

Всасывание углеводов
Всасывание моносахаридов представляет собой сложный биохимический процесс их транспорта через
Всасывание углеводов
Всасывание моносахаридов представляет собой сложный биохимический процесс их транспорта через
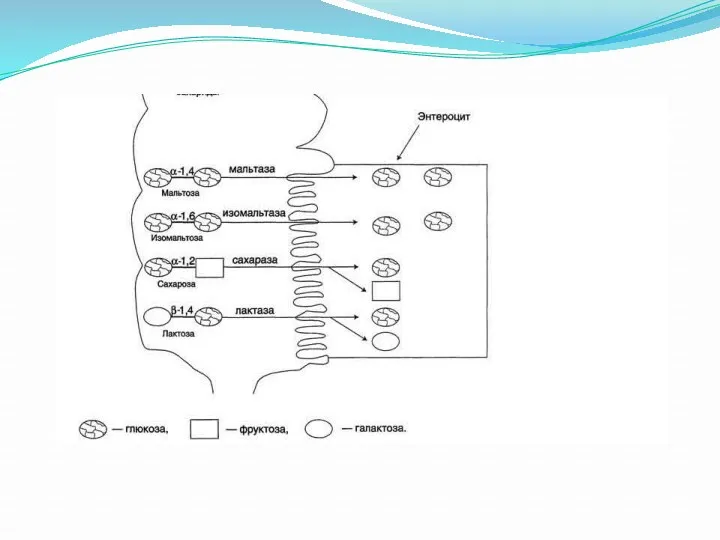
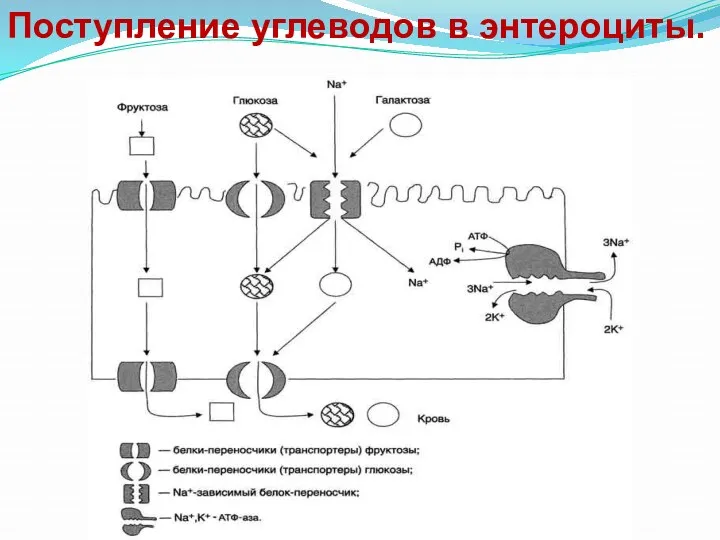
Поступление углеводов в энтероциты.
Поступление углеводов в энтероциты.
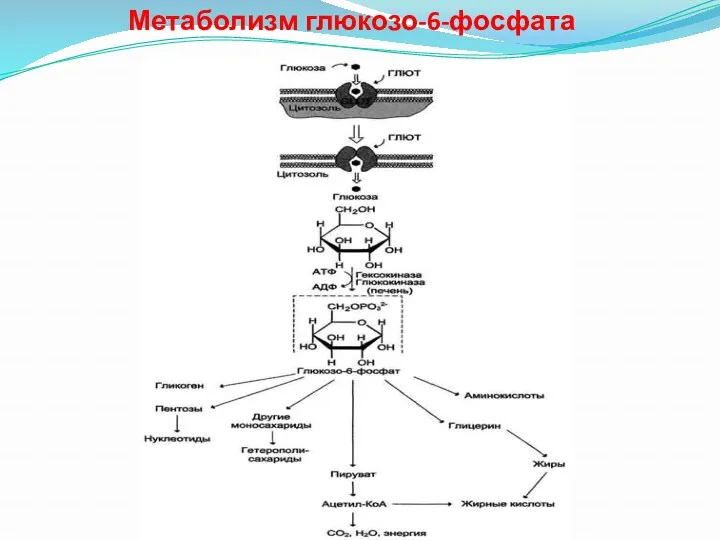
Метаболизм глюкозо-6-фосфата
Метаболизм глюкозо-6-фосфата
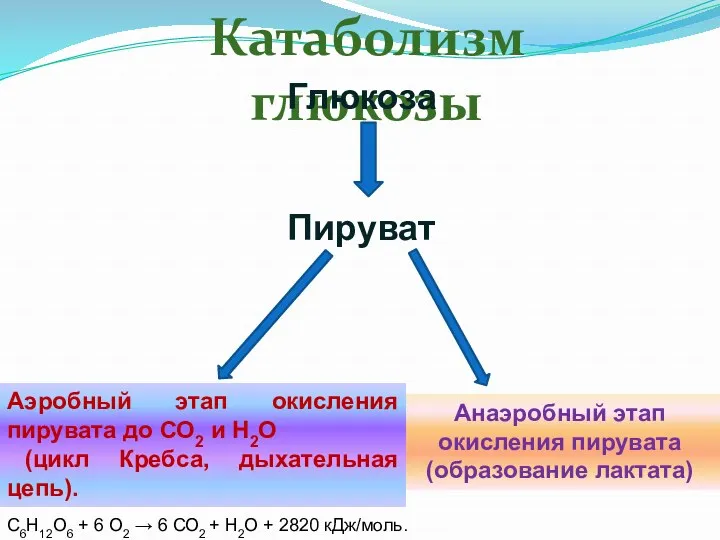
Катаболизм глюкозы
Анаэробный этап окисления пирувата
(образование лактата)
Аэробный этап окисления пирувата до СО2
Катаболизм глюкозы
Анаэробный этап окисления пирувата
(образование лактата)
Аэробный этап окисления пирувата до СО2
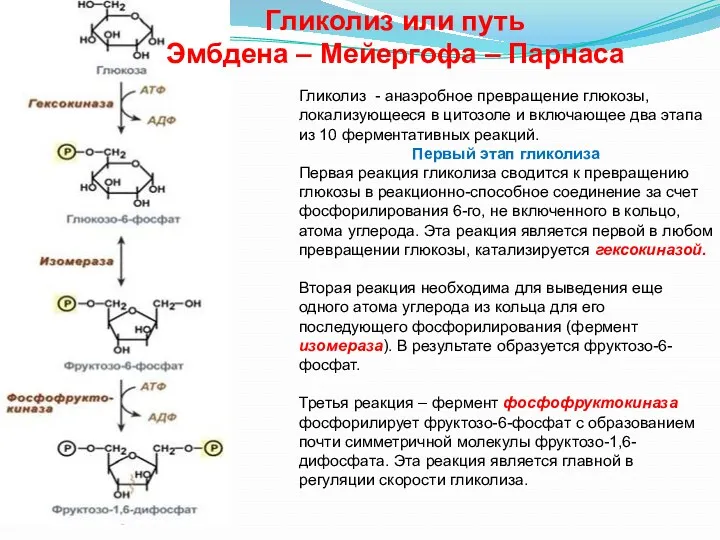
Гликолиз - анаэробное превращение глюкозы, локализующееся в цитозоле и включающее два
Гликолиз - анаэробное превращение глюкозы, локализующееся в цитозоле и включающее два
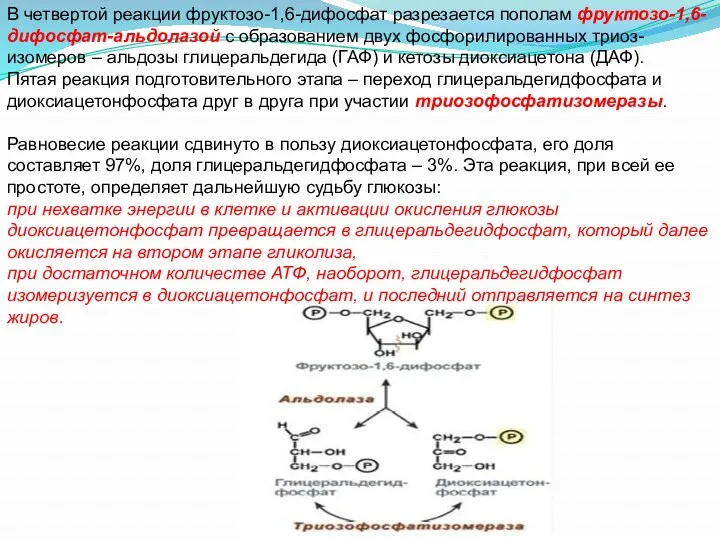
В четвертой реакции фруктозо-1,6-дифосфат разрезается пополам фруктозо-1,6-дифосфат-альдолазой с образованием двух фосфорилированных
В четвертой реакции фруктозо-1,6-дифосфат разрезается пополам фруктозо-1,6-дифосфат-альдолазой с образованием двух фосфорилированных
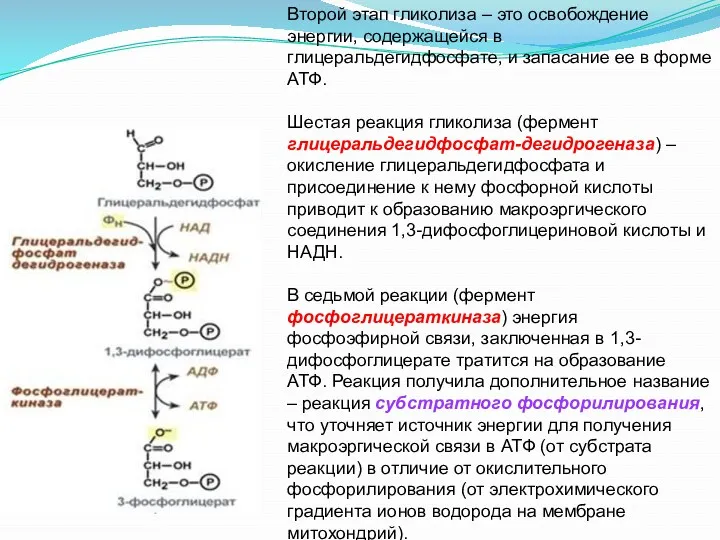
Второй этап гликолиза – это освобождение энергии, содержащейся в глицеральдегидфосфате, и
Второй этап гликолиза – это освобождение энергии, содержащейся в глицеральдегидфосфате, и
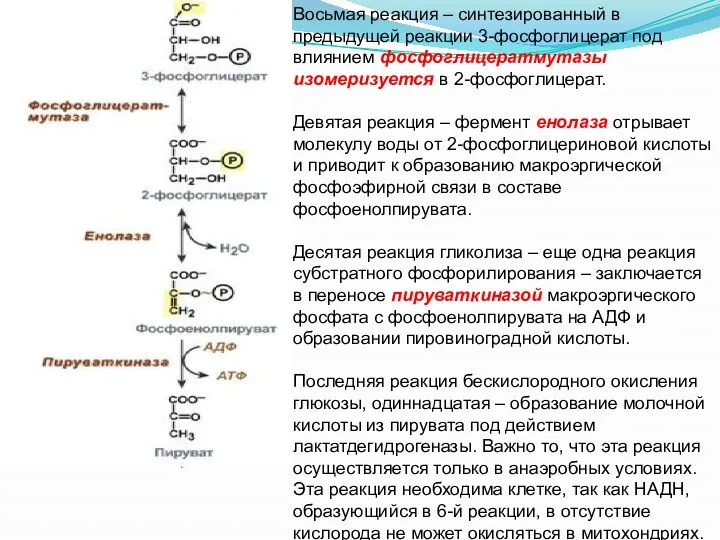
Восьмая реакция – синтезированный в предыдущей реакции 3-фосфоглицерат под влиянием фосфоглицератмутазы
Восьмая реакция – синтезированный в предыдущей реакции 3-фосфоглицерат под влиянием фосфоглицератмутазы
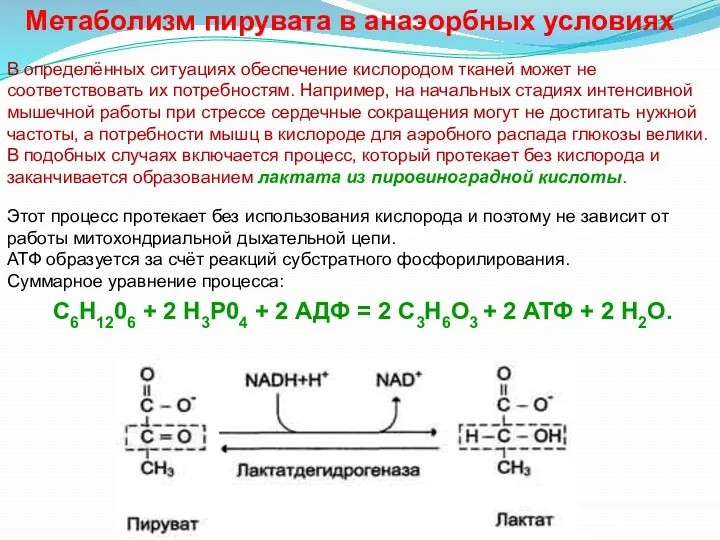
Метаболизм пирувата в анаэорбных условиях
С6Н1206 + 2 Н3Р04 + 2 АДФ
Метаболизм пирувата в анаэорбных условиях
С6Н1206 + 2 Н3Р04 + 2 АДФ
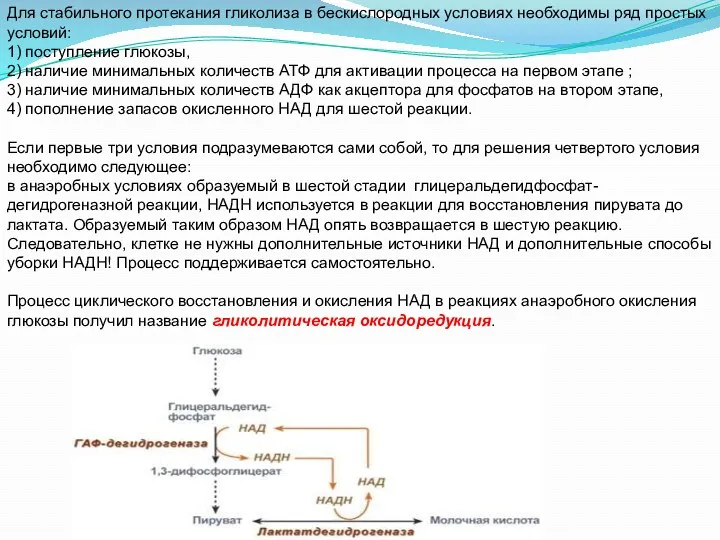
Для стабильного протекания гликолиза в бескислородных условиях необходимы ряд простых условий:
1)
Для стабильного протекания гликолиза в бескислородных условиях необходимы ряд простых условий:
1)

Баланс АТФ при анаэробном распаде глюкозы
Анаэробный гликолиз по сравнению с аэробным
Баланс АТФ при анаэробном распаде глюкозы
Анаэробный гликолиз по сравнению с аэробным

Биологическая значение анаэробного превращения глюкозы
Энергетическое. Анаэробное превращение глюкозы – источник АТФ
Биологическая значение анаэробного превращения глюкозы
Энергетическое. Анаэробное превращение глюкозы – источник АТФ

Это снижение потребления глюкозы и прекращение продукции молочной кислоты клеткой в
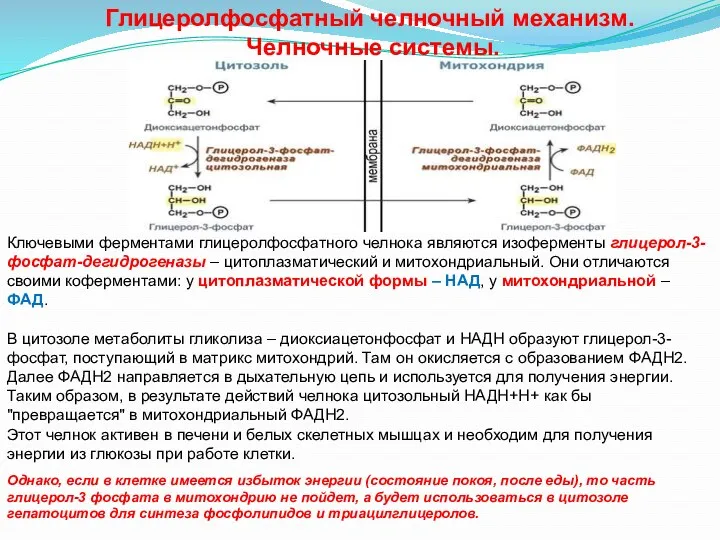
Глицеролфосфатный челночный механизм.
Челночные системы.
Ключевыми ферментами глицеролфосфатного челнока являются изоферменты глицерол-3-фосфат-дегидрогеназы
Глицеролфосфатный челночный механизм.
Челночные системы.
Ключевыми ферментами глицеролфосфатного челнока являются изоферменты глицерол-3-фосфат-дегидрогеназы
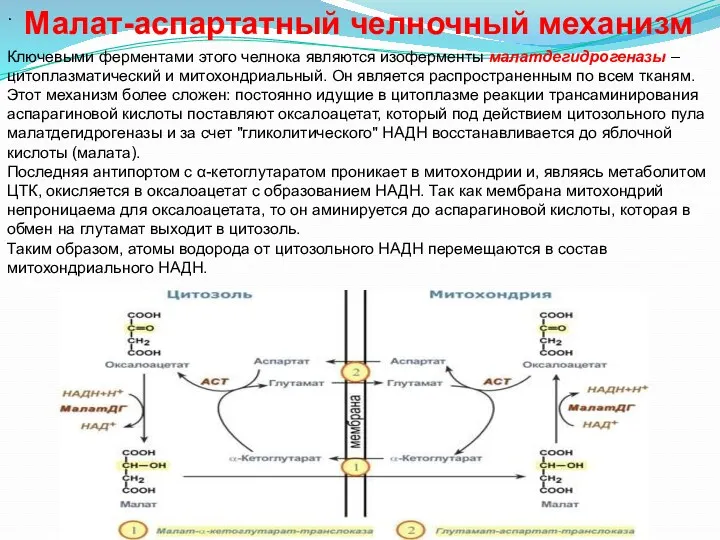
.
Ключевыми ферментами этого челнока являются изоферменты малатдегидрогеназы – цитоплазматический и
.
Ключевыми ферментами этого челнока являются изоферменты малатдегидрогеназы – цитоплазматический и

Из цитозоля пировиноградная кислота симпортом с ионами Н+ движется по протонному
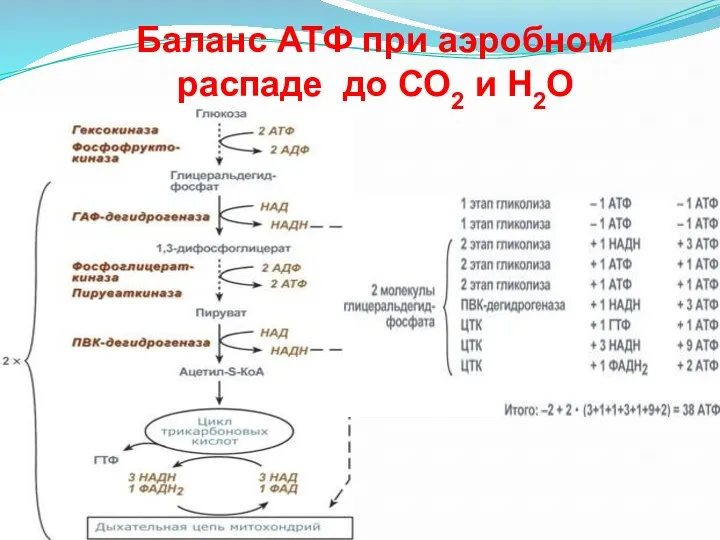
Баланс АТФ при аэробном
распаде до СО2 и Н2О
Баланс АТФ при аэробном
распаде до СО2 и Н2О

Физиологический смысл гликолиза
1. У аэробных организмов гликолиз является первым, подготовительным этапом
Физиологический смысл гликолиза
1. У аэробных организмов гликолиз является первым, подготовительным этапом

Это синтез глюкозы из неуглеводных компонентов: лактата, пирувата, глицерола, кетокислот цикла
Это синтез глюкозы из неуглеводных компонентов: лактата, пирувата, глицерола, кетокислот цикла
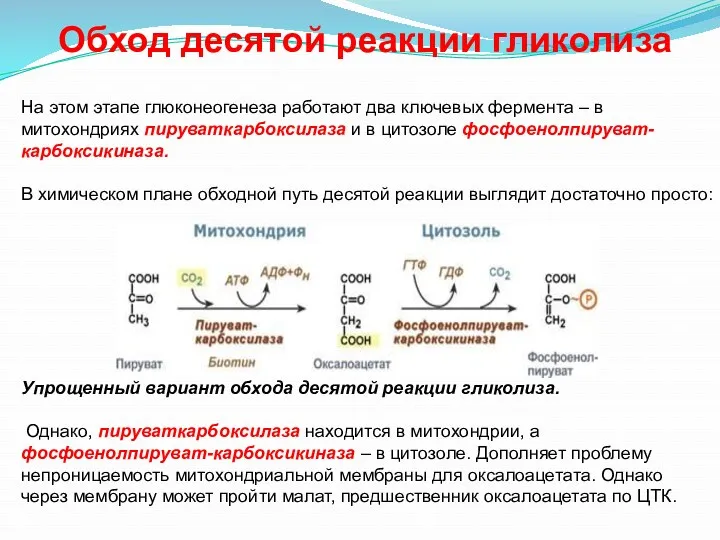
На этом этапе глюконеогенеза работают два ключевых фермента – в митохондриях
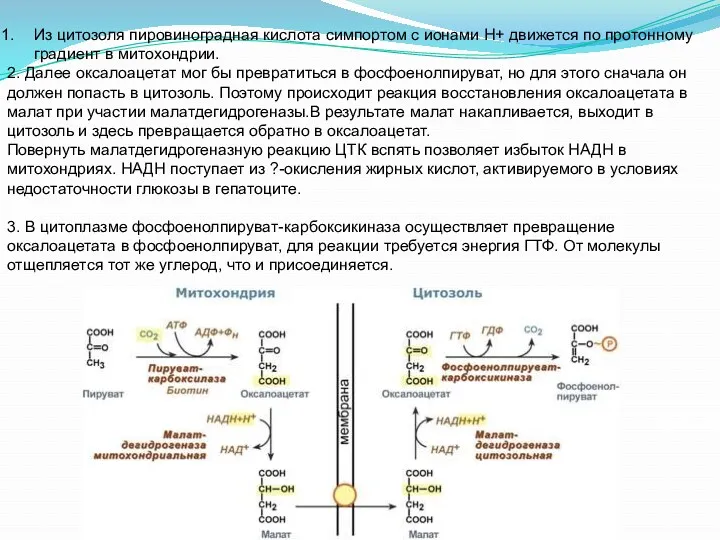
Из цитозоля пировиноградная кислота симпортом с ионами Н+ движется по
Из цитозоля пировиноградная кислота симпортом с ионами Н+ движется по
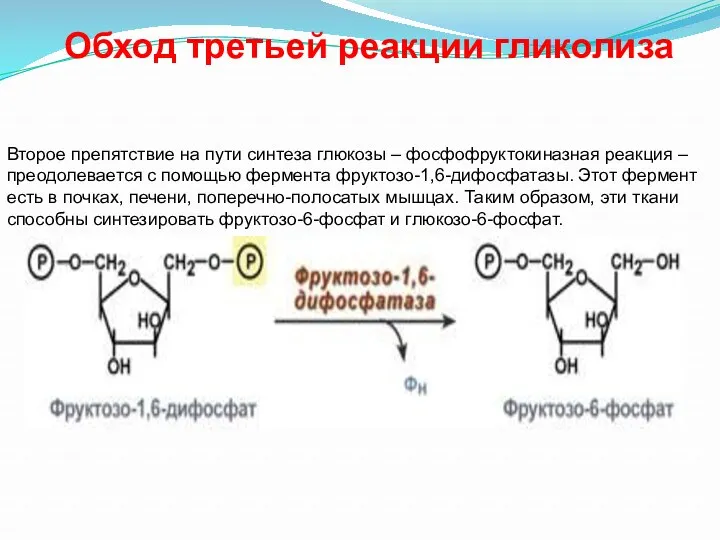
Обход третьей реакции гликолиза
Второе препятствие на пути синтеза глюкозы – фосфофруктокиназная
Обход третьей реакции гликолиза
Второе препятствие на пути синтеза глюкозы – фосфофруктокиназная
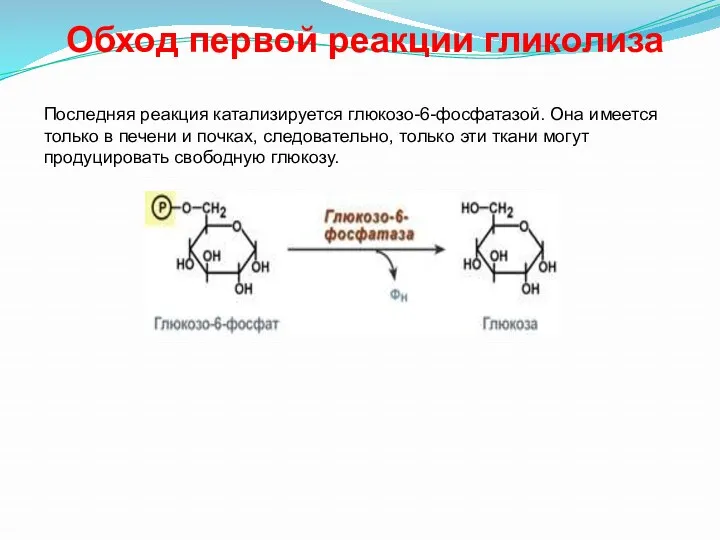
Обход первой реакции гликолиза
Последняя реакция катализируется глюкозо-6-фосфатазой. Она имеется только в
Обход первой реакции гликолиза
Последняя реакция катализируется глюкозо-6-фосфатазой. Она имеется только в

Гликолиз и глюконеогенез дополняют друг друга
Поскольку гликолитическое окисление глюкозы является способом
Гликолиз и глюконеогенез дополняют друг друга
Поскольку гликолитическое окисление глюкозы является способом
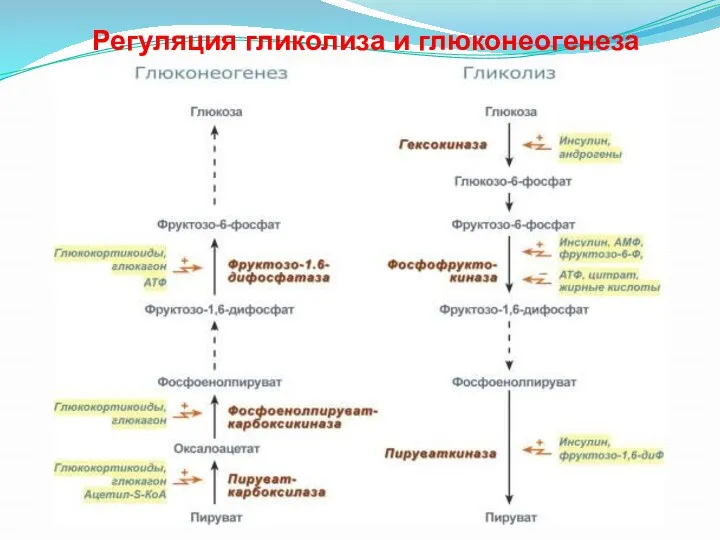
Регуляция гликолиза и глюконеогенеза
Регуляция гликолиза и глюконеогенеза
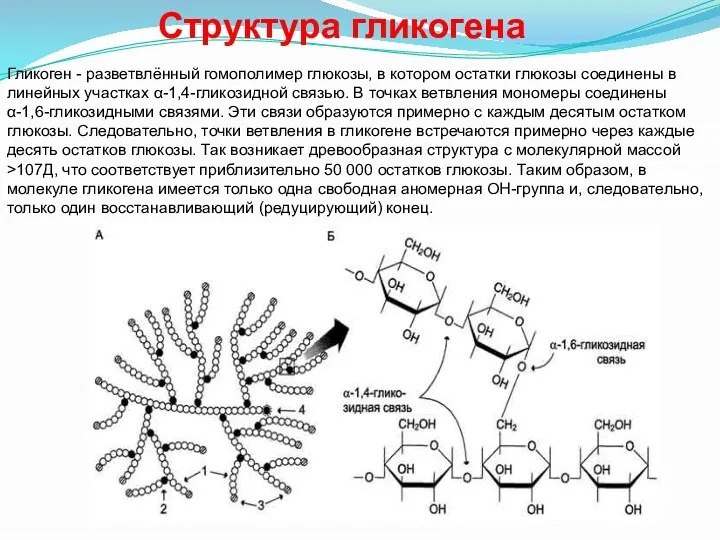
Гликоген - разветвлённый гомополимер глюкозы, в котором остатки глюкозы соединены в
Гликоген - разветвлённый гомополимер глюкозы, в котором остатки глюкозы соединены в
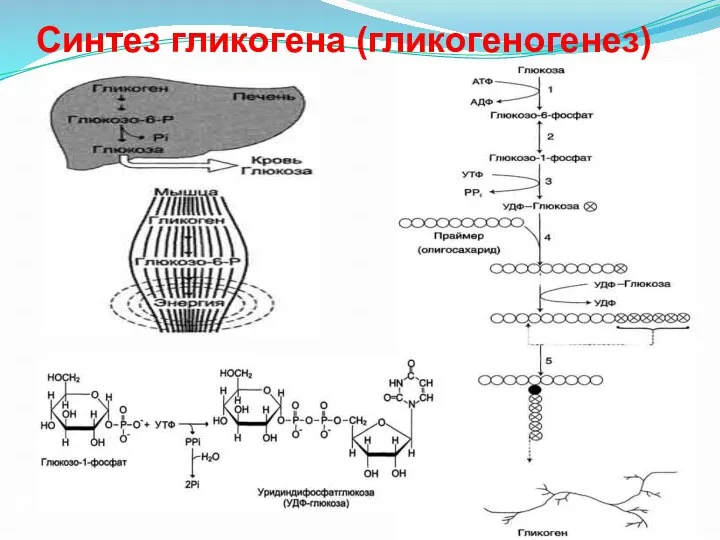
Синтез гликогена (гликогеногенез)
Синтез гликогена (гликогеногенез)

Глюкоза, поступающая в клетку, фосфорилируется при участии АТФ (реакция 1). Затем
Глюкоза, поступающая в клетку, фосфорилируется при участии АТФ (реакция 1). Затем
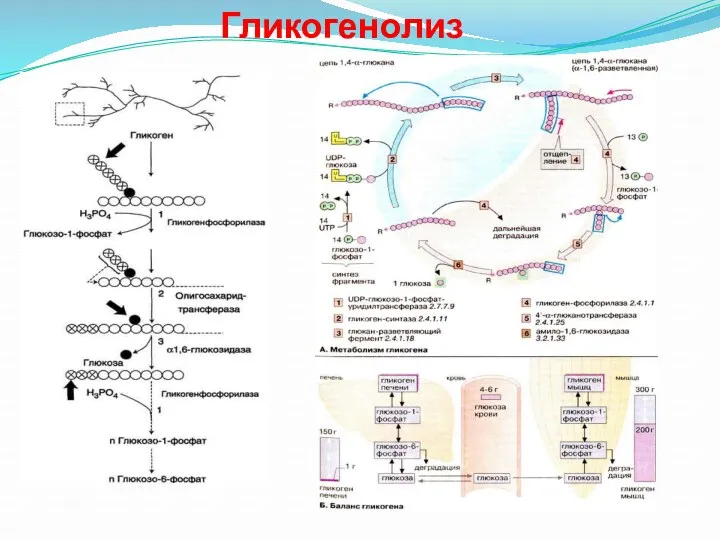
Гликогенолиз
Гликогенолиз

Распад гликогена или его мобилизация происходят в ответ на повышение потребности
Распад гликогена или его мобилизация происходят в ответ на повышение потребности

Считают, что перенос трёх остатков глюкозы и удаление мономера из точки
Считают, что перенос трёх остатков глюкозы и удаление мономера из точки

Сравнение этих процессов позволяет сделать следующие выводы:
а) синтез и
Сравнение этих процессов позволяет сделать следующие выводы:
а) синтез и
 Женская половая система
Женская половая система Бактерии. Грибы. Общая характеристика грибов
Бактерии. Грибы. Общая характеристика грибов Лишайники. Бактерии
Лишайники. Бактерии Презентация Организация исследовательской работы
Презентация Организация исследовательской работы Презентация по экологии
Презентация по экологии Клеточный цикл. Митоз
Клеточный цикл. Митоз Внутренняя среда организма. Кровь
Внутренняя среда организма. Кровь Цианобактерии
Цианобактерии Обмен веществ и энергии. Терморегуляция. Лекция № 20
Обмен веществ и энергии. Терморегуляция. Лекция № 20 Ферментті препараттар өндіру
Ферментті препараттар өндіру Рефлексы: типы, механизм
Рефлексы: типы, механизм Многообразие и значение Пресмыкающихся
Многообразие и значение Пресмыкающихся Раціональне харчування
Раціональне харчування цитология2
цитология2 Біотехнології контролю та очистки повітря
Біотехнології контролю та очистки повітря Биохимия соединительной ткани
Биохимия соединительной ткани Учение о внутренних органах (спланхнология). Лекция 6
Учение о внутренних органах (спланхнология). Лекция 6 Основные генетические термины и понятия
Основные генетические термины и понятия Строение и значение нервной системы
Строение и значение нервной системы Пентозофосфатный путь окисления глюкозы
Пентозофосфатный путь окисления глюкозы Презентация Класс млекопитающиеся для учащихся 7класса
Презентация Класс млекопитающиеся для учащихся 7класса Организм человека. Мифы и реальность
Организм человека. Мифы и реальность Развитие жизни в палеозойскую эру
Развитие жизни в палеозойскую эру Презентация по теме Биоритмы человека
Презентация по теме Биоритмы человека Сосальщики. Жизненный цикл
Сосальщики. Жизненный цикл Насекомые (для дошкольников)
Насекомые (для дошкольников) класс пресмыкающиеся
класс пресмыкающиеся Нуклеиновые кислоты. ДНК, РНК
Нуклеиновые кислоты. ДНК, РНК