- Общая вирусология

Содержание
- 2. История открытия первых вирусов 1.Вирус табачной мозаики - Д.И.Ивановский – 1892 г. 2.Бактериофаг - д’Эррель –
- 3. Д.И.Ивановский (1864 – 1920)
- 4. Стэнли Прузинер (1942)
- 5. Основные отличия вирусов от других форм жизни один тип нуклеиновой кислоты отсутствие клеточного строения белоксинтезирующих систем
- 6. Основные признаки, используемые для классификации вирусов тип нуклеиновой кислоты (ДНК/РНК) структура генома – количество нитей (цепочек)
- 7. Иерархическая система таксонов, применяемых в вирусологии Царство: Vira Подцарства: ДНК-геномные вирусы РНК-геномные вирусы Семейство Название таксона
- 8. КЛАССИФИКАЦИЯ ВИРУСОВ
- 9. КЛАССИФИКАЦИЯ ВИРУСОВ
- 10. Формы существования вирусов внеклеточная = вирион (структура) : НК капсид [суперкапсид] . Н-р, вирион имеет форму…
- 11. Принцип строения вириона Простой: НК+ капсид = нуклеокапсид Сложный: нуклеокапсид + суперкапсид
- 12. Типы симметрии капсида спиральная кубическая
- 13. Принцип строения суперкапсида билипидный слой матричный белок гликопротеины (шипы, ворсинки)
- 15. Форма вирионов
- 16. Общая характеристика ДНК вирионов форма: линейная кольцевая на концах – идентичные повторы: маркеры вирусной (не клеточной)
- 17. Общая характеристика РНК вирусов форма: линейная кольцевая структура: цельная фрагментированная информационная функция: +нить (позитивный геном) =
- 18. Общая характеристика белков вирусов Структурные капсидные «внутренние», гистоноподобные (НК ⇒ рибо/дезоксирибонуклеопротеин) Функциональные (ферменты) вирионные вирусиндуцированные вирус
- 19. Схема строения просто-устроенного вириона = паповавируса (вирус имеет двунитевую кольцевую ДНК)
- 20. Схема строения вируса гепатита А (вирус имеет однонитевую +РНК)
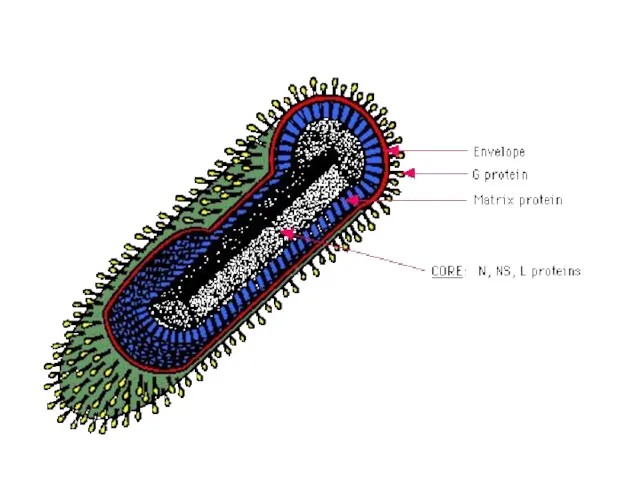
- 21. Схема строения сложно-устроенного вириона = вируса герпеса (вирус с линейной двухнитевой ДНК)
- 22. Схема строения вируса гриппа (вирус с однонитевой из 8 фрагментов минус РНК )
- 23. Строгий цитотропизм вирусов = Способность вирусов к репликации только в строго определённых клетках и органах поражаемая
- 24. Патологические процессы, вызываемые вирусами инфекционные (микробные) болезни = вирусные инфекции опухоли
- 25. Исходы вирусной инфекции клетки
- 26. Репродукция вирусов Различают три типа взаимодействия вируса с клеткой: 1. продуктивный тип, при котором образуются новые
- 27. Продуктивный тип взаимодействия вируса с клеткой Этапы размножения вирусов в чувствительной клетке: адсорбция вирионов на клетке
- 28. 1. Адсорбция вирионов на клетке = прикрепление вириона к поверхности клетки: 2 фазы: - неспецифическая –
- 29. Адсорбция вирионов на клетке Белки на поверхности вирусов наз-ся прикрепительными, чаще всего это гликопротеины. У просто
- 30. Адсорбция вирионов на клетке Рецепторы клеток: - белки, липиды, глигопротеины, гликолипиды и др. Н-р, сиаловая кислота
- 31. Адсорбция вирионов на клетке Избирательность поражения вирусами определенных клеток называется тропизм: клетки печени – гепатотропные, нервные
- 32. 2. Проникновение вируса в клетку 3 пути: Рецептор-зависимый эндоцитоз, слияние оболочки вириона с клеточной мембраной, смешанный.
- 33. Проникновение вируса в клетку 1.Рецептор-зависимый эндоцитоз – захватывание и поглощение вириона клеткой: Клеточная мембрана с вирионом
- 34. Проникновение вируса в клетку 2. Слияние оболочки вириона с клеточной мембраной = виропексис – характерно для
- 35. 2а.Депротеинизация вирусов = освобождение нуклеиновой кислоты путём сброса вирусом белковой (-ых) оболочки (-чек) При виропексисе –
- 36. «Раздевание» = депротеинизация вирусов начинается сразу после прикрепления к рецепторам и проникновения в клетку, продолжается в
- 37. «Раздевание» = депротеинизация вирусов Конечными продуктами раздевания являются: - нуклеиновая кислота - пикорнавирусы, - нуклеокапсид –
- 38. 3. Синтез вирусных компонентов = дизъюнктивная репродукция = синтез вирусных белков и нуклеиновых кислот, = происходит
- 39. 1. Синтез вирусных белков В зараженной клетке вирусный геном кодирует синтез 2-х групп белков: Структурные =
- 40. 1. Синтез вирусных белков 2 процесса составляют синтез белков: Транскрипция – переписывание генетической информации с нуклеиновой
- 41. 3.1.Синтез вирусных белков –варианты: ДНК-содержащие вирусы: Геномная ДНК вируса → транскрипция иРНК→трансляция белка вируса. Ферменты: -
- 42. 3.1. Синтез вирусных белков -варианты: Плюс-нитевые РНК-содержащие вирусы = вирусный геном выполняет функцию иРНК (пикорнавирусы, флавивирусы,
- 43. 3.1. Синтез вирусных белков -варианты: Минус-нитевые РНК-содержащие вирусы (ортомиксовирусы, парамиксовирусы, рабдовирусы) и двунитевые (реовирусы): Геномная РНК
- 44. 3.1.Синтез вирусных белков -варианты: Ретровирусы: – геном состоит из 2-х одинаковых молекул РНК = диплоидный, имеют
- 45. 3.2. Репликация вирусных геномов – зависит от типа нуклеиновой кислоты, - наличия вирусоспецифических или клеточных полимераз,
- 46. 3.2. Репликация вирусных геномов Двунитевые ДНК-вирусы (аденовирусы, герпесвирусы, поксвирусы) = полуконсервативный механизм, происходит в ядре (исключение
- 47. 3.2. Репликация вирусных геномов Двунитевые Особенность: гепаднавирусы→геном (кольцо) состоит из 2-х нитей: одна (неполная плюс-нить) короче
- 48. 3.2.Репликация вирусных геномов Однонитевые ДНК-вирусы(парвовирусы) – используют клеточные ДНК-полимеразы: - на исходной вирусной ДНК (+нить) синтезируется
- 49. 3.2.Репликация вирусных геномов Плюс-однонитевые РНК-вирусы (пикорнавирусы, флавивирусы, тогавирусы, полиовирусы) = геномная нить РНК выполняет функцию иРНК:
- 50. 3.2.Репликация вирусных геномов Минус-однонитевые РНК-вирусы (Рабдовирусы, парамиксовирусы, ортомиксовирусы) – имеют РНК-зависимую РНК-полимеразу: Минус-нитевая РНК + РНК-полимераза
- 51. 3.2.Репликация вирусных геномов Двунитевые РНК-вирусы (реовирусы, ротавирусы) – как у минус нитевых, но в цитоплазме клеток.
- 52. 3.2.Репликация вирусных геномов Ретровирусы = плюс-нитевые диплоидные РНК-содержащие вирусы, имеют обратную транскриптазу: обратная транскриптаза на матрице
- 53. 4.Формирование вирусов Происходит путем самосборки = составные части вируса транспортируются в определенный участок цитоплазмы или ядра
- 54. 4.Формирование вирусов сборка сложноустроенных вирусов: = сначала формируется нуклеокапсид, который взаимодействует с мембранами клетки: = вирусы,
- 55. 5.Выход вирусов из клетки 1. взрывной путь: клетка погибает и вирусы выходят наружу = простоустроенные вирусы,
- 56. Абортивный тип взаимодействия вируса с клеткой = прерывание инфекционного процесса в клетке на одном из этапов,
- 57. Абортивный тип взаимодействия вируса с клеткой Происходит когда: 1. чувствительные клетки заражаются дефектными вирусами или дефектными
- 58. Абортивный тип взаимодействия вируса с клеткой 2. стандартным вирусом заражаются генетически резистентные к нему клетки: Механизм
- 59. Абортивный тип взаимодействия вируса с клеткой 3. стандартным вирусом заражаются чувствительные клетки в неразрешающих (непермиссивных) условиях:
- 60. Интегративный тип взаимодействия вируса с клеткой = вирогения = нуклеиновая кислота вируса встраивается в хромосому клетки
- 61. Интегративный тип взаимодействия вируса с клеткой = вирогения ДНК-содержащие вирусы: Вирусная ДНК в кольцевой форме прикрепляется
- 62. Интегративный тип взаимодействия вируса с клеткой = вирогения РНК-содержащие вирусы: - синтез комплементарной нити ДНК на
- 63. Значение вирогении 1. Сохранение вирусной информации в составе клеточного генома = персистенция→ клетка при этом получает
- 64. Исходы интегративной вирусной инфекции
- 66. Скачать презентацию

История открытия первых вирусов
1.Вирус табачной мозаики -
Д.И.Ивановский – 1892 г.
2.Бактериофаг
История открытия первых вирусов
1.Вирус табачной мозаики -
Д.И.Ивановский – 1892 г.
2.Бактериофаг

Д.И.Ивановский (1864 – 1920)
Д.И.Ивановский (1864 – 1920)

Стэнли Прузинер (1942)
Стэнли Прузинер (1942)

Основные отличия вирусов от других форм жизни
один тип нуклеиновой кислоты
отсутствие
клеточного строения
белоксинтезирующих
Основные отличия вирусов от других форм жизни
один тип нуклеиновой кислоты
отсутствие
клеточного строения
белоксинтезирующих

Основные признаки, используемые для классификации вирусов
тип нуклеиновой кислоты (ДНК/РНК)
структура генома –
Основные признаки, используемые для классификации вирусов
тип нуклеиновой кислоты (ДНК/РНК)
структура генома –

Иерархическая система таксонов, применяемых в вирусологии
Царство: Vira
Подцарства: ДНК-геномные вирусы
РНК-геномные вирусы
Иерархическая система таксонов, применяемых в вирусологии
Царство: Vira
Подцарства: ДНК-геномные вирусы
РНК-геномные вирусы
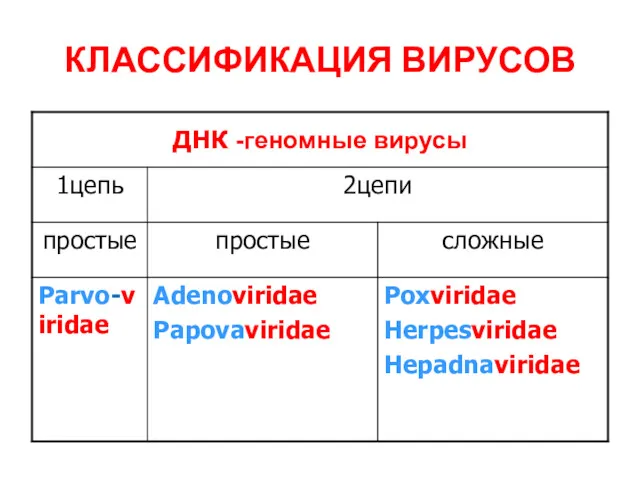
КЛАССИФИКАЦИЯ ВИРУСОВ
КЛАССИФИКАЦИЯ ВИРУСОВ

КЛАССИФИКАЦИЯ ВИРУСОВ
КЛАССИФИКАЦИЯ ВИРУСОВ

Формы существования вирусов
внеклеточная = вирион (структура) :
НК
капсид
[суперкапсид]
. Н-р, вирион имеет форму…
внутриклеточная
Формы существования вирусов
внеклеточная = вирион (структура) :
НК
капсид
[суперкапсид]
. Н-р, вирион имеет форму…
внутриклеточная
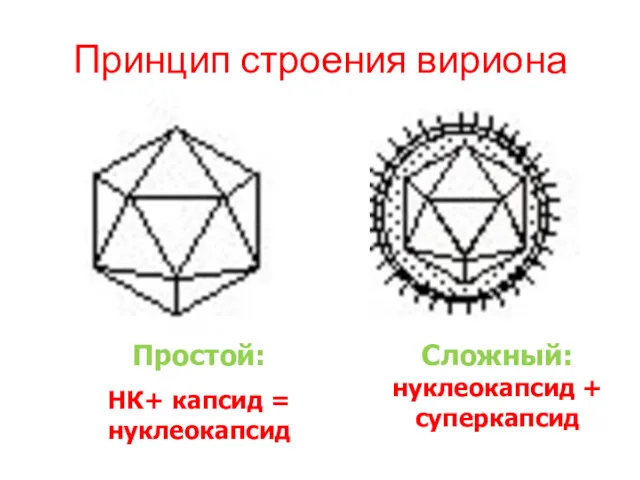
Принцип строения вириона
Простой:
НК+ капсид = нуклеокапсид
Сложный: нуклеокапсид + суперкапсид
Принцип строения вириона
Простой:
НК+ капсид = нуклеокапсид
Сложный: нуклеокапсид + суперкапсид
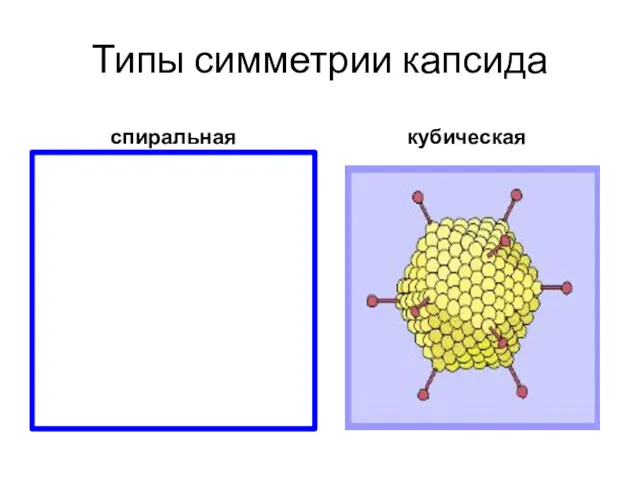
Типы симметрии капсида
спиральная
кубическая
Типы симметрии капсида
спиральная
кубическая
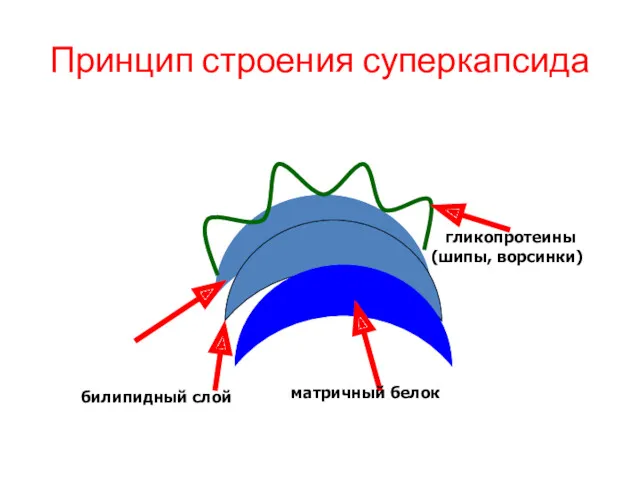
Принцип строения суперкапсида
билипидный слой
матричный белок
гликопротеины (шипы, ворсинки)
Принцип строения суперкапсида
билипидный слой
матричный белок
гликопротеины (шипы, ворсинки)
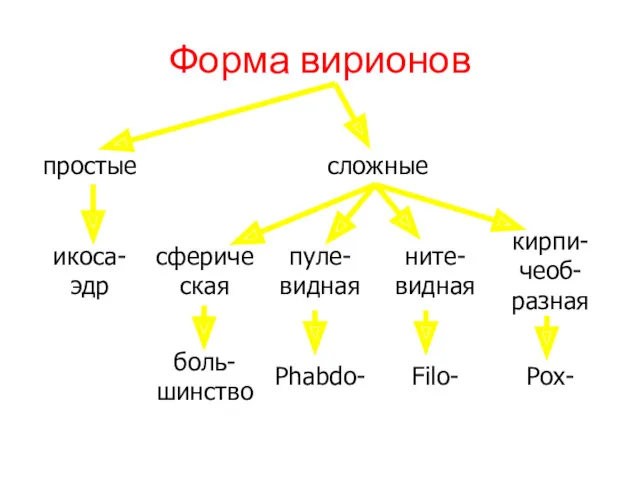
Форма вирионов
Форма вирионов

Общая характеристика ДНК вирионов
форма:
линейная
кольцевая
на концах – идентичные повторы:
маркеры вирусной (не клеточной)
Общая характеристика ДНК вирионов
форма:
линейная
кольцевая
на концах – идентичные повторы:
маркеры вирусной (не клеточной)

Общая характеристика РНК вирусов
форма:
линейная
кольцевая
структура:
цельная
фрагментированная
информационная функция:
+нить (позитивный геном) = иРНК
-нить (негативный геном)
Общая характеристика РНК вирусов
форма:
линейная
кольцевая
структура:
цельная
фрагментированная
информационная функция:
+нить (позитивный геном) = иРНК
-нить (негативный геном)

Общая характеристика белков вирусов
Структурные
капсидные
«внутренние», гистоноподобные (НК ⇒ рибо/дезоксирибонуклеопротеин)
Функциональные (ферменты)
вирионные
вирусиндуцированные
вирус может модифицировать
Общая характеристика белков вирусов
Структурные
капсидные
«внутренние», гистоноподобные (НК ⇒ рибо/дезоксирибонуклеопротеин)
Функциональные (ферменты)
вирионные
вирусиндуцированные
вирус может модифицировать

Схема строения
просто-устроенного вириона = паповавируса (вирус имеет двунитевую кольцевую ДНК)
Схема строения
просто-устроенного вириона = паповавируса (вирус имеет двунитевую кольцевую ДНК)
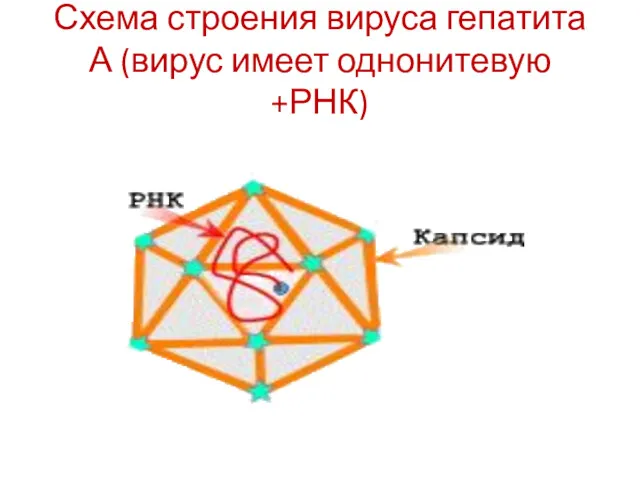
Схема строения вируса гепатита А (вирус имеет однонитевую +РНК)
Схема строения вируса гепатита А (вирус имеет однонитевую +РНК)
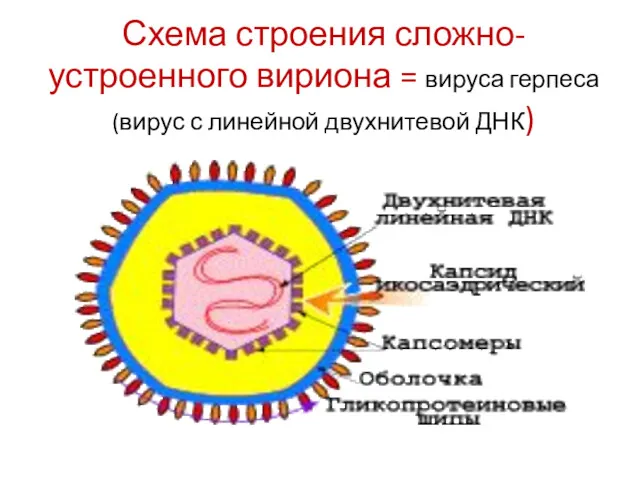
Схема строения сложно-устроенного вириона = вируса герпеса (вирус с линейной двухнитевой
Схема строения сложно-устроенного вириона = вируса герпеса (вирус с линейной двухнитевой
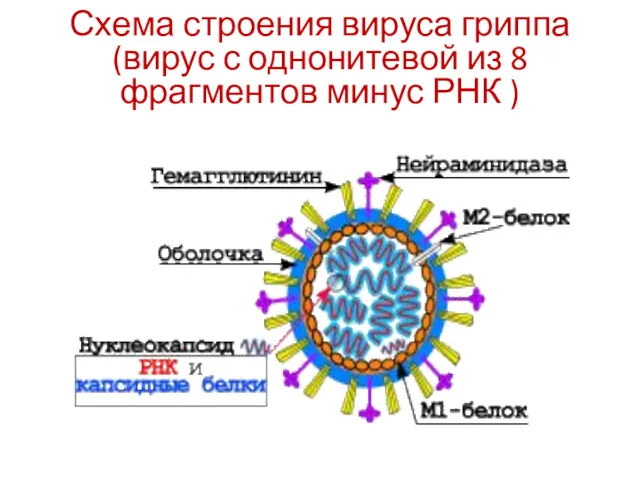
Схема строения вируса гриппа (вирус с однонитевой из 8 фрагментов минус
Схема строения вируса гриппа (вирус с однонитевой из 8 фрагментов минус

Строгий цитотропизм вирусов
= Способность вирусов к репликации только в строго определённых
Строгий цитотропизм вирусов
= Способность вирусов к репликации только в строго определённых

Патологические процессы, вызываемые вирусами
инфекционные (микробные) болезни = вирусные инфекции
опухоли
Патологические процессы, вызываемые вирусами
инфекционные (микробные) болезни = вирусные инфекции
опухоли
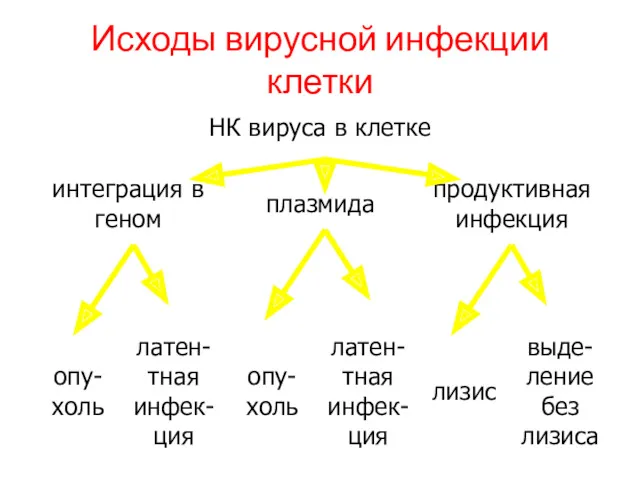
Исходы вирусной инфекции клетки
Исходы вирусной инфекции клетки

Репродукция вирусов
Различают три типа взаимодействия вируса с клеткой:
1. продуктивный тип,
Репродукция вирусов
Различают три типа взаимодействия вируса с клеткой:
1. продуктивный тип,

Продуктивный тип взаимодействия вируса с клеткой
Этапы размножения вирусов в чувствительной клетке:
адсорбция
Продуктивный тип взаимодействия вируса с клеткой
Этапы размножения вирусов в чувствительной клетке:
адсорбция

1. Адсорбция вирионов на клетке
= прикрепление вириона к поверхности клетки:
2 фазы:
1. Адсорбция вирионов на клетке
= прикрепление вириона к поверхности клетки:
2 фазы:

Адсорбция вирионов на клетке
Белки на поверхности вирусов наз-ся прикрепительными, чаще всего
Адсорбция вирионов на клетке
Белки на поверхности вирусов наз-ся прикрепительными, чаще всего

Адсорбция вирионов на клетке
Рецепторы клеток:
- белки,
липиды,
глигопротеины,
гликолипиды и
Адсорбция вирионов на клетке
Рецепторы клеток:
- белки,
липиды,
глигопротеины,
гликолипиды и

Адсорбция вирионов на клетке
Избирательность поражения вирусами определенных клеток называется тропизм:
клетки
Адсорбция вирионов на клетке
Избирательность поражения вирусами определенных клеток называется тропизм:
клетки

2. Проникновение вируса в клетку
3 пути:
Рецептор-зависимый эндоцитоз,
слияние оболочки вириона с клеточной
2. Проникновение вируса в клетку
3 пути:
Рецептор-зависимый эндоцитоз,
слияние оболочки вириона с клеточной

Проникновение вируса в клетку
1.Рецептор-зависимый эндоцитоз – захватывание и поглощение вириона клеткой:
Клеточная
Проникновение вируса в клетку
1.Рецептор-зависимый эндоцитоз – захватывание и поглощение вириона клеткой:
Клеточная

Проникновение вируса в клетку
2. Слияние оболочки вириона с клеточной мембраной =
Проникновение вируса в клетку
2. Слияние оболочки вириона с клеточной мембраной =

2а.Депротеинизация вирусов
= освобождение нуклеиновой кислоты путём сброса вирусом белковой (-ых) оболочки
2а.Депротеинизация вирусов
= освобождение нуклеиновой кислоты путём сброса вирусом белковой (-ых) оболочки

«Раздевание» = депротеинизация вирусов
начинается сразу после прикрепления к рецепторам и
«Раздевание» = депротеинизация вирусов
начинается сразу после прикрепления к рецепторам и

«Раздевание» = депротеинизация вирусов
Конечными продуктами раздевания являются:
- нуклеиновая кислота
«Раздевание» = депротеинизация вирусов
Конечными продуктами раздевания являются:
- нуклеиновая кислота

3. Синтез вирусных компонентов = дизъюнктивная репродукция
= синтез вирусных белков и
3. Синтез вирусных компонентов = дизъюнктивная репродукция
= синтез вирусных белков и

1. Синтез вирусных белков
В зараженной клетке вирусный геном кодирует синтез 2-х
1. Синтез вирусных белков
В зараженной клетке вирусный геном кодирует синтез 2-х

1. Синтез вирусных белков
2 процесса составляют синтез белков:
Транскрипция – переписывание генетической
1. Синтез вирусных белков
2 процесса составляют синтез белков:
Транскрипция – переписывание генетической

3.1.Синтез вирусных белков –варианты:
ДНК-содержащие вирусы:
Геномная ДНК вируса → транскрипция иРНК→трансляция белка
3.1.Синтез вирусных белков –варианты:
ДНК-содержащие вирусы:
Геномная ДНК вируса → транскрипция иРНК→трансляция белка

3.1. Синтез вирусных белков -варианты:
Плюс-нитевые РНК-содержащие вирусы = вирусный геном выполняет
3.1. Синтез вирусных белков -варианты:
Плюс-нитевые РНК-содержащие вирусы = вирусный геном выполняет

3.1. Синтез вирусных белков -варианты:
Минус-нитевые РНК-содержащие вирусы (ортомиксовирусы, парамиксовирусы, рабдовирусы) и
3.1. Синтез вирусных белков -варианты:
Минус-нитевые РНК-содержащие вирусы (ортомиксовирусы, парамиксовирусы, рабдовирусы) и

3.1.Синтез вирусных белков -варианты:
Ретровирусы:
– геном состоит из 2-х одинаковых молекул РНК
3.1.Синтез вирусных белков -варианты:
Ретровирусы:
– геном состоит из 2-х одинаковых молекул РНК

3.2. Репликация вирусных геномов
– зависит от типа нуклеиновой кислоты,
- наличия
3.2. Репликация вирусных геномов
– зависит от типа нуклеиновой кислоты,
- наличия

3.2. Репликация вирусных геномов
Двунитевые ДНК-вирусы (аденовирусы, герпесвирусы, поксвирусы) = полуконсервативный
3.2. Репликация вирусных геномов
Двунитевые ДНК-вирусы (аденовирусы, герпесвирусы, поксвирусы) = полуконсервативный

3.2. Репликация вирусных геномов
Двунитевые
Особенность: гепаднавирусы→геном (кольцо) состоит из 2-х
3.2. Репликация вирусных геномов
Двунитевые
Особенность: гепаднавирусы→геном (кольцо) состоит из 2-х

3.2.Репликация вирусных геномов
Однонитевые ДНК-вирусы(парвовирусы) – используют клеточные ДНК-полимеразы:
-
3.2.Репликация вирусных геномов
Однонитевые ДНК-вирусы(парвовирусы) – используют клеточные ДНК-полимеразы:
-

3.2.Репликация вирусных геномов
Плюс-однонитевые РНК-вирусы (пикорнавирусы, флавивирусы, тогавирусы, полиовирусы)
= геномная
3.2.Репликация вирусных геномов
Плюс-однонитевые РНК-вирусы (пикорнавирусы, флавивирусы, тогавирусы, полиовирусы)
= геномная

3.2.Репликация вирусных геномов
Минус-однонитевые РНК-вирусы (Рабдовирусы, парамиксовирусы, ортомиксовирусы) – имеют
3.2.Репликация вирусных геномов
Минус-однонитевые РНК-вирусы (Рабдовирусы, парамиксовирусы, ортомиксовирусы) – имеют

3.2.Репликация вирусных геномов
Двунитевые РНК-вирусы (реовирусы, ротавирусы) – как у
3.2.Репликация вирусных геномов
Двунитевые РНК-вирусы (реовирусы, ротавирусы) – как у

3.2.Репликация вирусных геномов
Ретровирусы = плюс-нитевые диплоидные РНК-содержащие вирусы, имеют
3.2.Репликация вирусных геномов
Ретровирусы = плюс-нитевые диплоидные РНК-содержащие вирусы, имеют

4.Формирование вирусов
Происходит путем самосборки = составные части вируса транспортируются в определенный
4.Формирование вирусов
Происходит путем самосборки = составные части вируса транспортируются в определенный

4.Формирование вирусов
сборка сложноустроенных вирусов:
= сначала формируется нуклеокапсид, который взаимодействует с мембранами
4.Формирование вирусов
сборка сложноустроенных вирусов:
= сначала формируется нуклеокапсид, который взаимодействует с мембранами

5.Выход вирусов из клетки
1. взрывной путь: клетка погибает и вирусы выходят
5.Выход вирусов из клетки
1. взрывной путь: клетка погибает и вирусы выходят

Абортивный тип взаимодействия вируса с клеткой
= прерывание инфекционного процесса в клетке
Абортивный тип взаимодействия вируса с клеткой
= прерывание инфекционного процесса в клетке

Абортивный тип взаимодействия вируса с клеткой
Происходит когда:
1. чувствительные клетки
Абортивный тип взаимодействия вируса с клеткой
Происходит когда:
1. чувствительные клетки

Абортивный тип взаимодействия вируса с клеткой
2. стандартным вирусом заражаются генетически резистентные
Абортивный тип взаимодействия вируса с клеткой
2. стандартным вирусом заражаются генетически резистентные

Абортивный тип взаимодействия вируса с клеткой
3. стандартным вирусом заражаются чувствительные клетки
Абортивный тип взаимодействия вируса с клеткой
3. стандартным вирусом заражаются чувствительные клетки

Интегративный тип взаимодействия вируса с клеткой = вирогения
= нуклеиновая кислота вируса
Интегративный тип взаимодействия вируса с клеткой = вирогения
= нуклеиновая кислота вируса

Интегративный тип взаимодействия вируса с клеткой = вирогения
ДНК-содержащие вирусы:
Вирусная ДНК в
Интегративный тип взаимодействия вируса с клеткой = вирогения
ДНК-содержащие вирусы:
Вирусная ДНК в

Интегративный тип взаимодействия вируса с клеткой = вирогения
РНК-содержащие вирусы:
- синтез комплементарной
Интегративный тип взаимодействия вируса с клеткой = вирогения
РНК-содержащие вирусы:
- синтез комплементарной

Значение вирогении
1. Сохранение вирусной информации в составе клеточного генома = персистенция→
Значение вирогении
1. Сохранение вирусной информации в составе клеточного генома = персистенция→
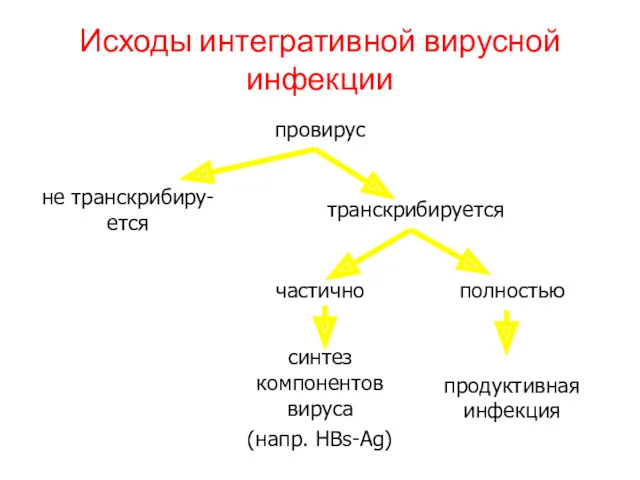
Исходы интегративной вирусной инфекции
Исходы интегративной вирусной инфекции
 Самые опасные насекомые в мире
Самые опасные насекомые в мире Мегагаметофит. Мегаспорангий
Мегагаметофит. Мегаспорангий Клетка.10 класс
Клетка.10 класс Почему мы любим кошек и собак
Почему мы любим кошек и собак Анатомия тройничного нерва
Анатомия тройничного нерва Биология ирбиса, или снежного барса
Биология ирбиса, или снежного барса Царство животные. Тип хордовые. Класс птицы
Царство животные. Тип хордовые. Класс птицы Насекомые леса
Насекомые леса Строение растений
Строение растений Полезные насекомые
Полезные насекомые Стратегия, принципы и цели устойчивого развития
Стратегия, принципы и цели устойчивого развития Комбінативна мінливість
Комбінативна мінливість Тип Иглокожие
Тип Иглокожие Мышцы. Работа мышц
Мышцы. Работа мышц Работа мышц
Работа мышц Презентация Насекомые Красной Книги
Презентация Насекомые Красной Книги Глаз человека как оптическая система. Построение изображения на сетчатке
Глаз человека как оптическая система. Построение изображения на сетчатке Животные Африки, Австралии, Южной Америки и Северной Америки
Животные Африки, Австралии, Южной Америки и Северной Америки Животные. Природоведение. 5 класс
Животные. Природоведение. 5 класс Вегетативное размножение растений
Вегетативное размножение растений Анализирующее скрещивание. Дигибридное скрещивание. Третий закон Менделя
Анализирующее скрещивание. Дигибридное скрещивание. Третий закон Менделя Отдел моховидные
Отдел моховидные Организация и технология промышленного семеноводства
Организация и технология промышленного семеноводства Эволюция животных
Эволюция животных Определение пола
Определение пола Железы
Железы Питьевая вода
Питьевая вода Палеонтология и эволюция
Палеонтология и эволюция