- Общая вирусология

Содержание
- 2. История открытия первых вирусов 1.Вирус табачной мозаики - Д.И.Ивановский – 1892 г. 2.Бактериофаг - д’Эррель –
- 3. Д.И.Ивановский (1864 – 1920)
- 4. Стэнли Прузинер (1942)
- 5. Основные отличия вирусов от других форм жизни один тип нуклеиновой кислоты отсутствие клеточного строения белоксинтезирующих систем
- 6. Основные признаки, используемые для классификации вирусов тип нуклеиновой кислоты (ДНК/РНК) структура генома – количество нитей (цепочек)
- 7. Иерархическая система таксонов, применяемых в вирусологии Царство: Vira Подцарства: ДНК-геномные вирусы РНК-геномные вирусы Семейство Название таксона
- 8. КЛАССИФИКАЦИЯ ВИРУСОВ
- 9. КЛАССИФИКАЦИЯ ВИРУСОВ
- 10. Формы существования вирусов внеклеточная = вирион (структура) : НК капсид [суперкапсид] . Н-р, вирион имеет форму…
- 11. Принцип строения вириона Простой: НК+ капсид = нуклеокапсид Сложный: нуклеокапсид + суперкапсид
- 12. Типы симметрии капсида спиральная кубическая
- 13. Принцип строения суперкапсида билипидный слой матричный белок гликопротеины (шипы, ворсинки)
- 15. Форма вирионов
- 16. Общая характеристика ДНК вирионов форма: линейная кольцевая на концах – идентичные повторы: маркеры вирусной (не клеточной)
- 17. Общая характеристика РНК вирусов форма: линейная кольцевая структура: цельная фрагментированная информационная функция: +нить (позитивный геном) =
- 18. Общая характеристика белков вирусов Структурные капсидные «внутренние», гистоноподобные (НК ⇒ рибо/дезоксирибонуклеопротеин) Функциональные (ферменты) вирионные вирусиндуцированные вирус
- 19. Схема строения паповавируса (вирус имеет двунитевую кольцевую ДНК) просто-устроенный вирион
- 20. Схема строения вируса герпеса (вирус с линейной двухнитевой ДНК) сложно-устроенный вирион
- 21. Схема строения вируса гепатита А (вирус имеет однонитевую +РНК)
- 22. Схема строения вируса гриппа (вирус с однонитевой из 8 фрагментов минус РНК )
- 23. Строгий цитотропизм вирусов = Способность вирусов к репликации только в строго определённых клетках и органах поражаемая
- 24. Патологические процессы, вызываемые вирусами инфекционные (микробные) болезни = вирусные инфекции опухоли
- 25. Репродукция вирусов Различают три типа взаимодействия вируса с клеткой: 1. продуктивный тип, при котором образуются новые
- 26. Исходы вирусной инфекции клетки
- 27. Продуктивный тип взаимодействия вируса с клеткой Этапы размножения вирусов в чувствительной клетке: адсорбция вирионов на клетке
- 28. Адсорбция вирионов на клетке = прикрепление вириона к поверхности клетки: 2 фазы: - неспецифическая – ионное
- 29. Адсорбция вирионов на клетке Рецепторами клеток м.б. белки, липиды, глигопротеины, гликолипиды и др. Н-р, сиаловая кислота
- 30. Проникновение вируса в клетку 3 пути: Рецептор-зависимый эндоцитоз, слияние оболочки вириона с клеточной мембраной, смешанный.
- 31. Проникновение вируса в клетку 1.Рецептор-зависимый эндоцитоз – захватывание и поглощение вириона клеткой: Клеточная мембрана с вирионом
- 32. Проникновение вируса в клетку 2. Слияние оболочки вириона с клеточной мембраной = виропексис – характерно для
- 33. Депротеинизация вирусов = Освобождение нуклеиновой кислоты путём сброса вирусом белковой (-ых) оболочки (-чек) При виропексисе –
- 34. «Раздевание» = депротеинизация вирусов начинается сразу после прикрепления к рецепторам и проникновения в клетку, продолжается в
- 35. Синтез вирусных компонентов = дизъюнктивная репродукция = синтез вирусных белков и нуклеиновых кислот, = происходит в
- 36. Синтез вирусных компонентов = дизъюнктивная репродукция 1. Синтез вирусных белков В зараженной клетке вирусный геном кодирует
- 37. Синтез вирусных компонентов = дизъюнктивная репродукция 1. Синтез вирусных белков 2 процесса составляют синтез белков: Транскрипция
- 38. Синтез вирусных компонентов = дизъюнктивная репродукция 1.Синтез вирусных белков – варианты: ДНК-содержащие вирусы: Геномная ДНК вируса
- 39. Синтез вирусных компонентов = дизъюнктивная репродукция 1. Синтез вирусных белков -варианты: Плюс-нитевые РНК-содержащие вирусы = вирусный
- 40. Синтез вирусных компонентов = дизъюнктивная репродукция Синтез вирусных белков -варианты: Ретровирусы: – геном состоит из 2-х
- 41. Синтез вирусных компонентов = дизъюнктивная репродукция 2. Репликация вирусных геномов – зависит от типа нуклеиновой кислоты,
- 42. Синтез вирусных компонентов = дизъюнктивная репродукция 2. Репликация вирусных геномов 1.Двунитевые ДНК-вирусы (аденовирусы, герпесвирусы, поксвирусы) =
- 43. Синтез вирусных компонентов = дизъюнктивная репродукция 2.Репликация вирусных геномов 2.2. Однонитевые ДНК-вирусы(парвовирусы) – используют клеточные ДНК-полимеразы:
- 44. Синтез вирусных компонентов = дизъюнктивная репродукция 2.Репликация вирусных геномов 2.3. Плюс-однонитевые РНК-вирусы (пикорнавирусы, флавивирусы, тогавирусы, полиовирусы)
- 45. Синтез вирусных компонентов = дизъюнктивная репродукция 2.Репликация вирусных геномов 2.4. Минус-однонитевые РНК-вирусы (Рабдовирусы, парамиксовирусы, ортомиксовирусы) –
- 46. Синтез вирусных компонентов = дизъюнктивная репродукция 2.Репликация вирусных геномов 2.5. Двунитевые РНК-вирусы (реовирусы, ротавирусы) – как
- 47. Синтез вирусных компонентов = дизъюнктивная репродукция 2.Репликация вирусных геномов 2.6. Ретровирусы = плюс-нитевые диплоидные РНК-содержащие вирусы,
- 48. Формирование вирусов Происходит путем самосборки = составные части вируса транспортируются в определенный участок цитоплазмы или ядра
- 49. Выход вирусов из клетки 1. взрывной путь: клетка погибает и вирусы выходят наружу = простоустроенные вирусы,
- 50. Абортивный тип взаимодействия вируса с клеткой = прерывание инфекционного процесса в клетке на одном из этапов,
- 51. Абортивный тип взаимодействия вируса с клеткой 2. стандартным вирусом заражаются генетически резистентные к нему клетки: Механизм
- 52. Интегративный тип взаимодействия вируса с клеткой = вирогения = нуклеиновая кислота вируса встраивается в хромосому клетки
- 53. Значение вирогении 1. Сохранение вирусной информации в составе клеточного генома = персистенция→ клетка при этом получает
- 54. Исходы интегративной вирусной инфекции
- 55. Исходы активации персистирующего вируса рецидив того же заболевания развитие другого заболевания, вызываемого тем же самым вирусом
- 56. Способы культивирования вирусов куриный эмбрион культура клеток организм лабораторного животного ⇓ обнаружение наличия вируса (индикация) ⇓
- 57. Использование для вирусологического метода куриного эмбриона 5-7-дневные, реже – 10-11-дневные
- 58. Основные способы заражения куриных эмбрионов на хорион-аллантоисную оболочку в хорион-аллантоисную полость в полость желточного мешка в
- 59. Обнаружение вирусов в курином эмбрионе индикация: гибель эмбриона морфологические изменения эмбриона/оболочек РГА с жидкостью из полостей
- 60. Использование культур клеток Культуры клеток = соматические или эмбриональные клетки человека или животных, культивируемые в лабораторных
- 61. Использование культур клеток Чаще – перевиваемые монослойные индикация: ЦПД (цитопатическое действие вирусов – любое изменение клеток
- 62. Первичные культуры клеток получают из тканей (эмбриональных или нормальных) многоклеточных организмов. Такие клетки не способны к
- 63. Перевиваемые культуры клеток Перевиваемые = стабильные = готовят из опухолевых клеток, способных длительно размножаться in vitro
- 64. Преимущества перевиваемых культур клеток перед первичными: продолжительность культивирования – десятки лет, высокая скорость размножения, меньшая трудоемкость,
- 65. Полуперевиваемые культуры клеток – диплоидные клетки различных тканей и органов, способные к ограниченному размножению in vitro.
- 66. Условия культивирования клеток: Питательные среды сложного состава (среда 199, Игла), сод-т источники энергии (глюкозу), минеральные вещества,
- 67. Обнаружение = индикация вирусов в культуре клеток проводят на основе следующих феноменов: - цитопатогенного действия (ЦПД)
- 68. ЦПД - видимые под микроскопом морфологические изменения клеток (вплоть до их отторжения от стекла), возникающие в
- 69. Виды ЦПД округление и сморщивание клеток – пикорнавирусы, нарастающая деструкция – герпесвирусы, пролиферация (образование дырок) –
- 70. ЦПД вирусов
- 71. Включения = скопление вирионов или отдельных их компонентов в цитоплазме или ядре клеток, выявляемые под микроскопом
- 72. Тельца Бабеша-Негри
- 73. Бляшки, или “негативные” колонии = ограниченные участки разрушенных вирусами клеток, культивируемых на питательной среде под агаровым
- 74. Реакция гемагглютинации (РГА) основана на способности некоторых вирусов вызывать агглютинацию (склеивание) эритроцитов за счет вирусных гликопротеиновых
- 75. Реакция гемадсорбции =РГАдс = способность культур клеток, инфицированных вирусами, адсорбировать на своей поверхности эритроциты.
- 76. Реакция иммунофлюоресценции (РИФ)
- 77. Использование лабораторных животных взрослые или новорожденные белые мыши, хомяки, кролики, обезьяны применяется для выделения тех вирусов,
- 78. Способы заражения лабораторных животных интраназально, подкожно, внутримышечно, внутрибрюшинно, интрацеребрально,
- 79. Обнаружение вируса при заражении лабораторных животных обнаруживают вирус по: - развитию видимых клинических проявлений – параличи
- 80. Прионы – белковые молекулы, способные вызывать разрушение клеток организма человека и животных. Они характеризуются устойчивостью: к
- 81. Прионы Прионный белок может существовать в двух формах: нормальная клеточная форма(РrPc) - обнаруживается в организме всех
- 82. Прионы инфекционная форма (PrPs) – характеризуется: измененной вторичной и третичной структурой молекулы, высокой устойчивостью к нагреванию,
- 88. Скачать презентацию

История открытия первых вирусов
1.Вирус табачной мозаики -
Д.И.Ивановский – 1892 г.
2.Бактериофаг
История открытия первых вирусов
1.Вирус табачной мозаики -
Д.И.Ивановский – 1892 г.
2.Бактериофаг

Д.И.Ивановский (1864 – 1920)
Д.И.Ивановский (1864 – 1920)

Стэнли Прузинер (1942)
Стэнли Прузинер (1942)

Основные отличия вирусов от других форм жизни
один тип нуклеиновой кислоты
отсутствие
клеточного строения
белоксинтезирующих
Основные отличия вирусов от других форм жизни
один тип нуклеиновой кислоты
отсутствие
клеточного строения
белоксинтезирующих

Основные признаки, используемые для классификации вирусов
тип нуклеиновой кислоты (ДНК/РНК)
структура генома –
Основные признаки, используемые для классификации вирусов
тип нуклеиновой кислоты (ДНК/РНК)
структура генома –

Иерархическая система таксонов, применяемых в вирусологии
Царство: Vira
Подцарства: ДНК-геномные вирусы
РНК-геномные вирусы
Иерархическая система таксонов, применяемых в вирусологии
Царство: Vira
Подцарства: ДНК-геномные вирусы
РНК-геномные вирусы
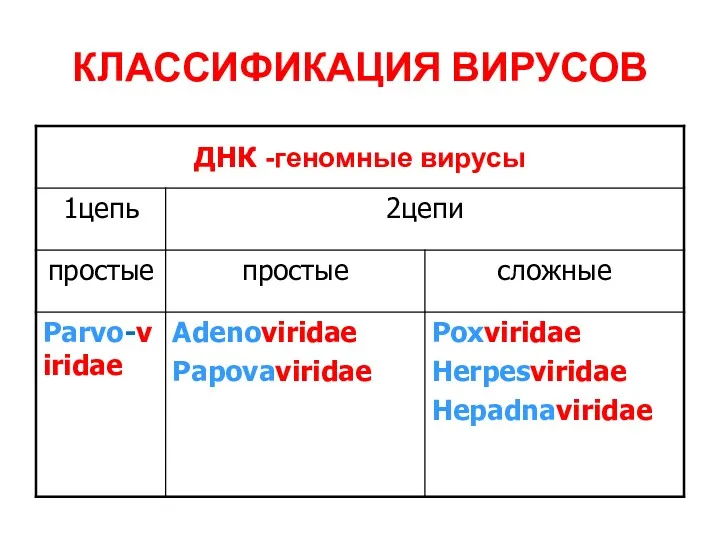
КЛАССИФИКАЦИЯ ВИРУСОВ
КЛАССИФИКАЦИЯ ВИРУСОВ
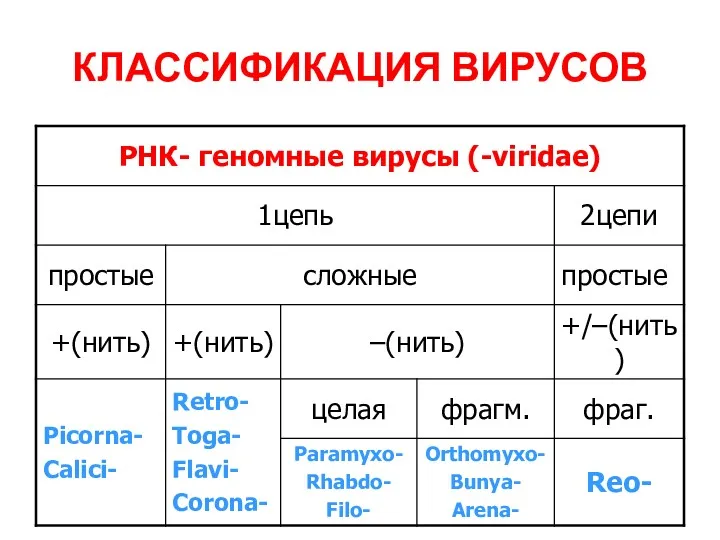
КЛАССИФИКАЦИЯ ВИРУСОВ
КЛАССИФИКАЦИЯ ВИРУСОВ

Формы существования вирусов
внеклеточная = вирион (структура) :
НК
капсид
[суперкапсид]
. Н-р, вирион имеет форму…
внутриклеточная
Формы существования вирусов
внеклеточная = вирион (структура) :
НК
капсид
[суперкапсид]
. Н-р, вирион имеет форму…
внутриклеточная
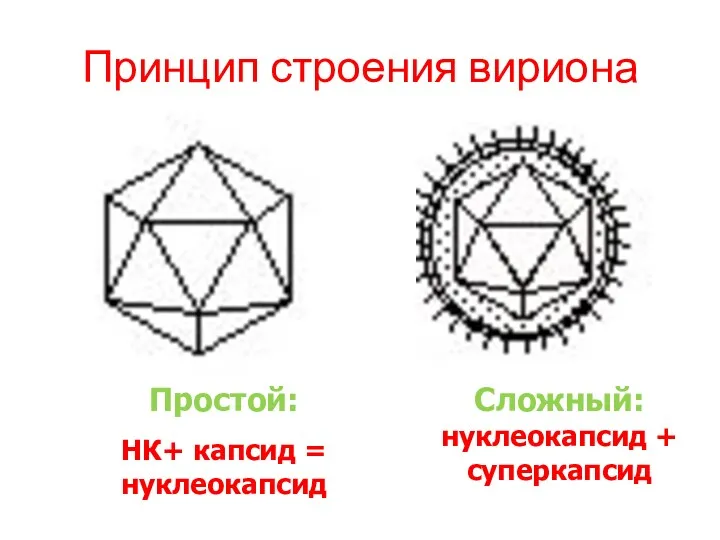
Принцип строения вириона
Простой:
НК+ капсид = нуклеокапсид
Сложный: нуклеокапсид + суперкапсид
Принцип строения вириона
Простой:
НК+ капсид = нуклеокапсид
Сложный: нуклеокапсид + суперкапсид
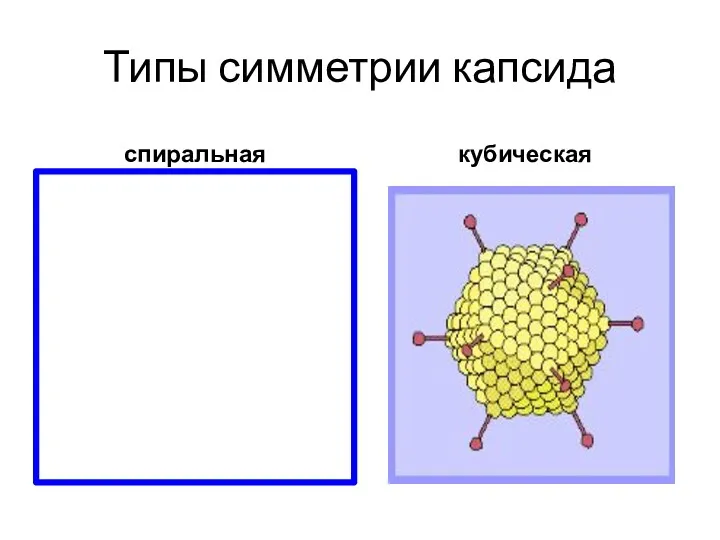
Типы симметрии капсида
спиральная
кубическая
Типы симметрии капсида
спиральная
кубическая
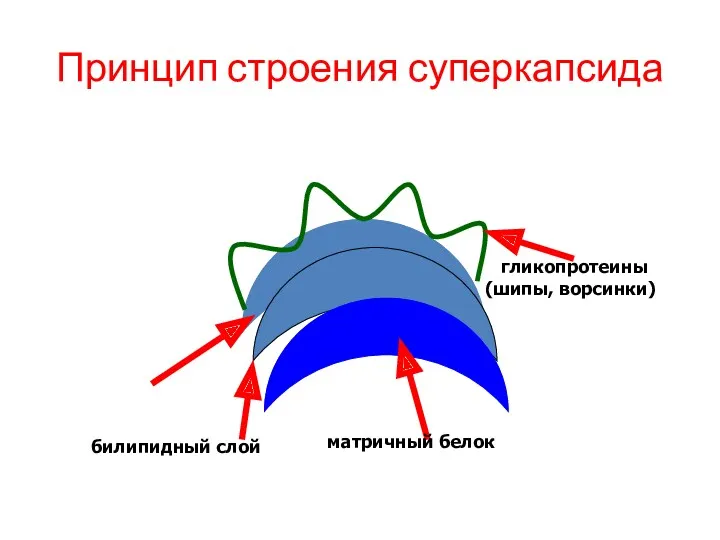
Принцип строения суперкапсида
билипидный слой
матричный белок
гликопротеины (шипы, ворсинки)
Принцип строения суперкапсида
билипидный слой
матричный белок
гликопротеины (шипы, ворсинки)
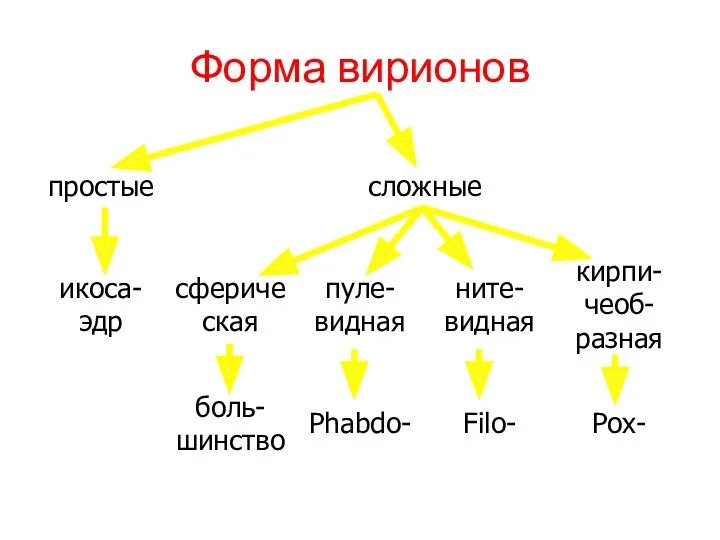
Форма вирионов
Форма вирионов

Общая характеристика ДНК вирионов
форма:
линейная
кольцевая
на концах – идентичные повторы:
маркеры вирусной (не клеточной)
Общая характеристика ДНК вирионов
форма:
линейная
кольцевая
на концах – идентичные повторы:
маркеры вирусной (не клеточной)

Общая характеристика РНК вирусов
форма:
линейная
кольцевая
структура:
цельная
фрагментированная
информационная функция:
+нить (позитивный геном) = иРНК
-нить (негативный геном)
Общая характеристика РНК вирусов
форма:
линейная
кольцевая
структура:
цельная
фрагментированная
информационная функция:
+нить (позитивный геном) = иРНК
-нить (негативный геном)

Общая характеристика белков вирусов
Структурные
капсидные
«внутренние», гистоноподобные (НК ⇒ рибо/дезоксирибонуклеопротеин)
Функциональные (ферменты)
вирионные
вирусиндуцированные
вирус может модифицировать
Общая характеристика белков вирусов
Структурные
капсидные
«внутренние», гистоноподобные (НК ⇒ рибо/дезоксирибонуклеопротеин)
Функциональные (ферменты)
вирионные
вирусиндуцированные
вирус может модифицировать
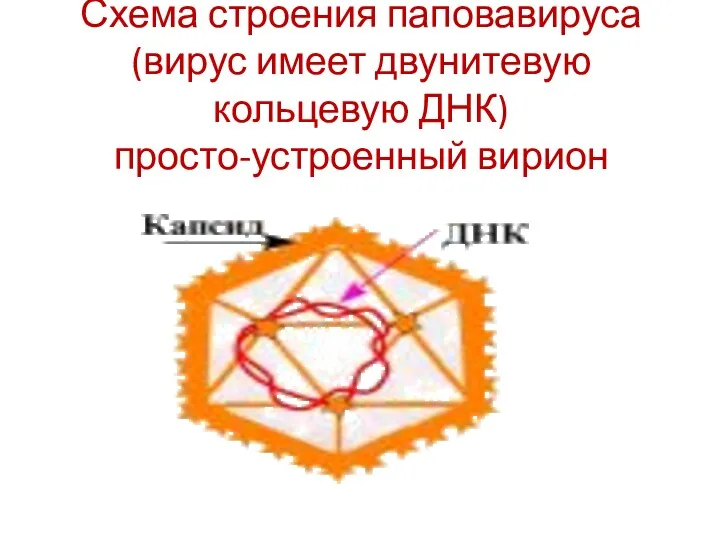
Схема строения паповавируса (вирус имеет двунитевую кольцевую ДНК)
просто-устроенный вирион
Схема строения паповавируса (вирус имеет двунитевую кольцевую ДНК)
просто-устроенный вирион
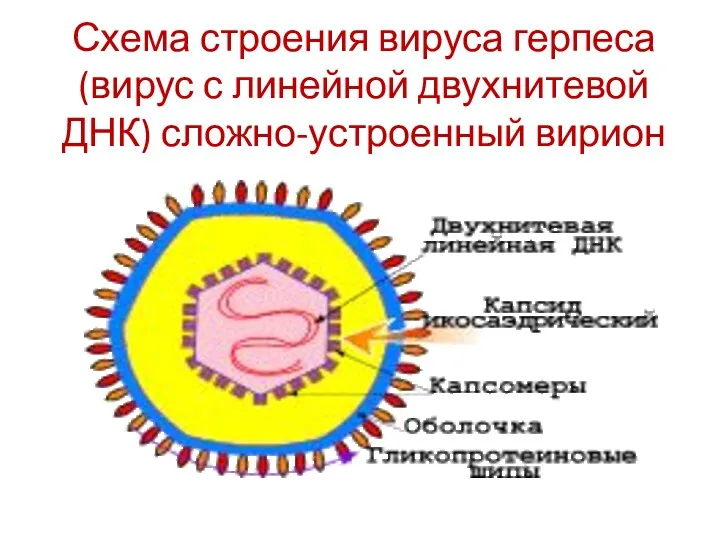
Схема строения вируса герпеса (вирус с линейной двухнитевой ДНК) сложно-устроенный вирион
Схема строения вируса герпеса (вирус с линейной двухнитевой ДНК) сложно-устроенный вирион
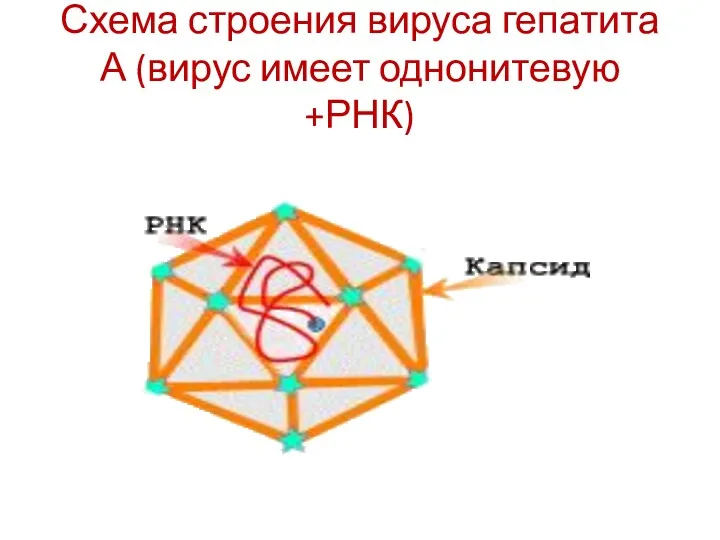
Схема строения вируса гепатита А (вирус имеет однонитевую +РНК)
Схема строения вируса гепатита А (вирус имеет однонитевую +РНК)
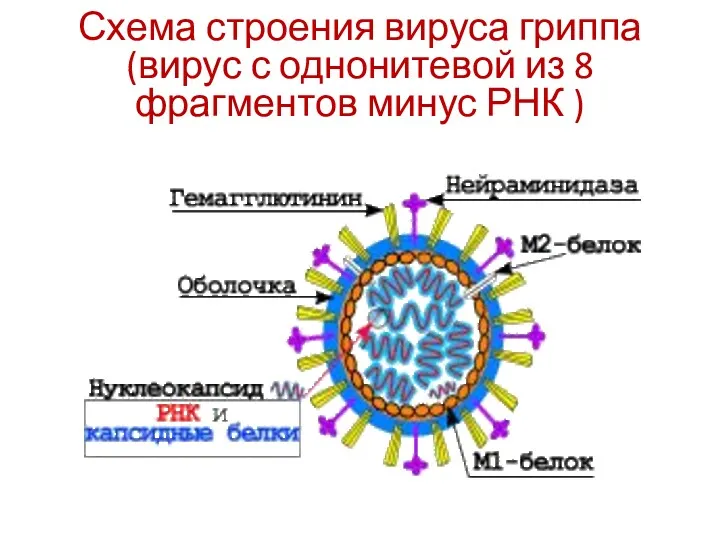
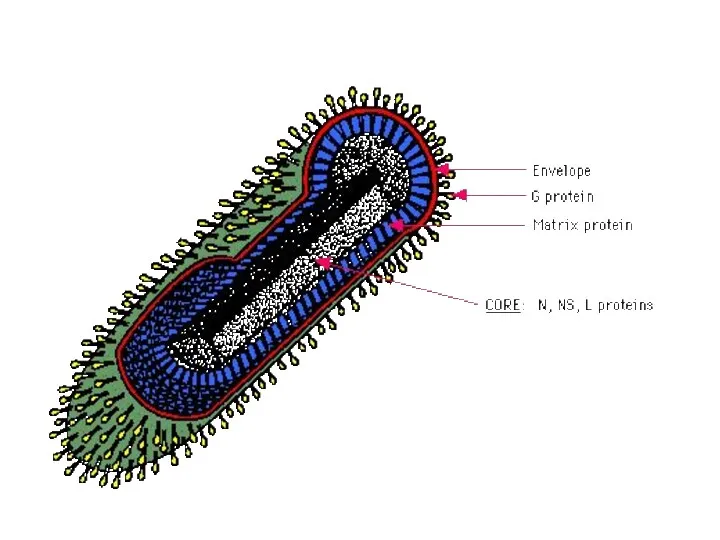
Схема строения вируса гриппа (вирус с однонитевой из 8 фрагментов минус
Схема строения вируса гриппа (вирус с однонитевой из 8 фрагментов минус

Строгий цитотропизм вирусов
= Способность вирусов к репликации только в строго определённых
Строгий цитотропизм вирусов
= Способность вирусов к репликации только в строго определённых

Патологические процессы, вызываемые вирусами
инфекционные (микробные) болезни = вирусные инфекции
опухоли
Патологические процессы, вызываемые вирусами
инфекционные (микробные) болезни = вирусные инфекции
опухоли

Репродукция вирусов
Различают три типа взаимодействия вируса с клеткой:
1. продуктивный тип,
Репродукция вирусов
Различают три типа взаимодействия вируса с клеткой:
1. продуктивный тип,
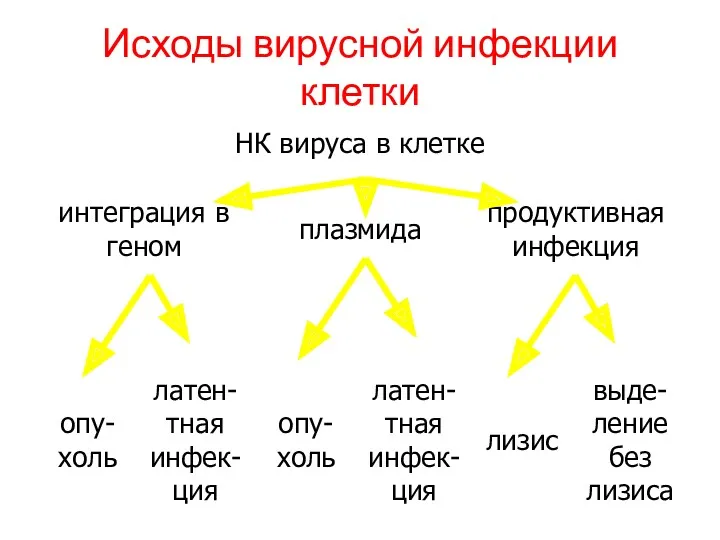
Исходы вирусной инфекции клетки
Исходы вирусной инфекции клетки

Продуктивный тип взаимодействия вируса с клеткой
Этапы размножения вирусов в чувствительной клетке:
адсорбция
Продуктивный тип взаимодействия вируса с клеткой
Этапы размножения вирусов в чувствительной клетке:
адсорбция

Адсорбция вирионов на клетке
= прикрепление вириона к поверхности клетки:
2 фазы:
-
Адсорбция вирионов на клетке
= прикрепление вириона к поверхности клетки:
2 фазы:
-

Адсорбция вирионов на клетке
Рецепторами клеток м.б. белки, липиды, глигопротеины, гликолипиды и
Адсорбция вирионов на клетке
Рецепторами клеток м.б. белки, липиды, глигопротеины, гликолипиды и

Проникновение вируса в клетку
3 пути:
Рецептор-зависимый эндоцитоз,
слияние оболочки вириона с клеточной мембраной,
смешанный.
Проникновение вируса в клетку
3 пути:
Рецептор-зависимый эндоцитоз,
слияние оболочки вириона с клеточной мембраной,
смешанный.

Проникновение вируса в клетку
1.Рецептор-зависимый эндоцитоз – захватывание и поглощение вириона клеткой:
Клеточная
Проникновение вируса в клетку
1.Рецептор-зависимый эндоцитоз – захватывание и поглощение вириона клеткой:
Клеточная

Проникновение вируса в клетку
2. Слияние оболочки вириона с клеточной мембраной =
Проникновение вируса в клетку
2. Слияние оболочки вириона с клеточной мембраной =

Депротеинизация вирусов
= Освобождение нуклеиновой кислоты путём сброса вирусом белковой (-ых) оболочки
Депротеинизация вирусов
= Освобождение нуклеиновой кислоты путём сброса вирусом белковой (-ых) оболочки

«Раздевание» = депротеинизация вирусов
начинается сразу после прикрепления к рецепторам и
«Раздевание» = депротеинизация вирусов
начинается сразу после прикрепления к рецепторам и

Синтез вирусных компонентов = дизъюнктивная репродукция
= синтез вирусных белков и нуклеиновых
Синтез вирусных компонентов = дизъюнктивная репродукция
= синтез вирусных белков и нуклеиновых

Синтез вирусных компонентов = дизъюнктивная репродукция
1. Синтез вирусных белков
В зараженной клетке
Синтез вирусных компонентов = дизъюнктивная репродукция
1. Синтез вирусных белков
В зараженной клетке

Синтез вирусных компонентов = дизъюнктивная репродукция
1. Синтез вирусных белков
2 процесса составляют
Синтез вирусных компонентов = дизъюнктивная репродукция
1. Синтез вирусных белков
2 процесса составляют

Синтез вирусных компонентов = дизъюнктивная репродукция
1.Синтез вирусных белков – варианты:
ДНК-содержащие вирусы:
Геномная
Синтез вирусных компонентов = дизъюнктивная репродукция
1.Синтез вирусных белков – варианты:
ДНК-содержащие вирусы:
Геномная

Синтез вирусных компонентов = дизъюнктивная репродукция
1. Синтез вирусных белков -варианты:
Плюс-нитевые РНК-содержащие
Синтез вирусных компонентов = дизъюнктивная репродукция
1. Синтез вирусных белков -варианты:
Плюс-нитевые РНК-содержащие

Синтез вирусных компонентов = дизъюнктивная репродукция
Синтез вирусных белков -варианты:
Ретровирусы:
– геном состоит
Синтез вирусных компонентов = дизъюнктивная репродукция
Синтез вирусных белков -варианты:
Ретровирусы:
– геном состоит

Синтез вирусных компонентов = дизъюнктивная репродукция
2. Репликация вирусных геномов
– зависит
Синтез вирусных компонентов = дизъюнктивная репродукция
2. Репликация вирусных геномов
– зависит

Синтез вирусных компонентов = дизъюнктивная репродукция
2. Репликация вирусных геномов
1.Двунитевые ДНК-вирусы
Синтез вирусных компонентов = дизъюнктивная репродукция
2. Репликация вирусных геномов
1.Двунитевые ДНК-вирусы

Синтез вирусных компонентов = дизъюнктивная репродукция
2.Репликация вирусных геномов
2.2. Однонитевые ДНК-вирусы(парвовирусы)
Синтез вирусных компонентов = дизъюнктивная репродукция
2.Репликация вирусных геномов
2.2. Однонитевые ДНК-вирусы(парвовирусы)

Синтез вирусных компонентов = дизъюнктивная репродукция
2.Репликация вирусных геномов
2.3. Плюс-однонитевые РНК-вирусы
Синтез вирусных компонентов = дизъюнктивная репродукция
2.Репликация вирусных геномов
2.3. Плюс-однонитевые РНК-вирусы

Синтез вирусных компонентов = дизъюнктивная репродукция
2.Репликация вирусных геномов
2.4. Минус-однонитевые РНК-вирусы
Синтез вирусных компонентов = дизъюнктивная репродукция
2.Репликация вирусных геномов
2.4. Минус-однонитевые РНК-вирусы

Синтез вирусных компонентов = дизъюнктивная репродукция
2.Репликация вирусных геномов
2.5. Двунитевые РНК-вирусы
Синтез вирусных компонентов = дизъюнктивная репродукция
2.Репликация вирусных геномов
2.5. Двунитевые РНК-вирусы

Синтез вирусных компонентов = дизъюнктивная репродукция
2.Репликация вирусных геномов
2.6. Ретровирусы =
Синтез вирусных компонентов = дизъюнктивная репродукция
2.Репликация вирусных геномов
2.6. Ретровирусы =

Формирование вирусов
Происходит путем самосборки = составные части вируса транспортируются в определенный
Формирование вирусов
Происходит путем самосборки = составные части вируса транспортируются в определенный

Выход вирусов из клетки
1. взрывной путь: клетка погибает и вирусы выходят
Выход вирусов из клетки
1. взрывной путь: клетка погибает и вирусы выходят

Абортивный тип взаимодействия вируса с клеткой
= прерывание инфекционного процесса в клетке
Абортивный тип взаимодействия вируса с клеткой
= прерывание инфекционного процесса в клетке

Абортивный тип взаимодействия вируса с клеткой
2. стандартным вирусом заражаются генетически резистентные
Абортивный тип взаимодействия вируса с клеткой
2. стандартным вирусом заражаются генетически резистентные

Интегративный тип взаимодействия вируса с клеткой = вирогения
= нуклеиновая кислота вируса
Интегративный тип взаимодействия вируса с клеткой = вирогения
= нуклеиновая кислота вируса

Значение вирогении
1. Сохранение вирусной информации в составе клеточного генома = персистенция→
Значение вирогении
1. Сохранение вирусной информации в составе клеточного генома = персистенция→
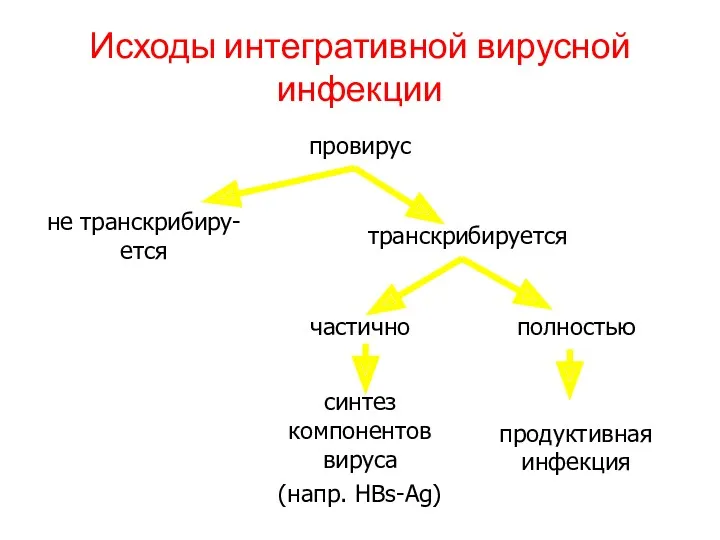
Исходы интегративной вирусной инфекции
Исходы интегративной вирусной инфекции

Исходы активации персистирующего вируса
рецидив того же заболевания
развитие другого заболевания, вызываемого тем
Исходы активации персистирующего вируса
рецидив того же заболевания
развитие другого заболевания, вызываемого тем

Способы культивирования вирусов
куриный эмбрион
культура клеток
организм лабораторного животного
⇓
обнаружение наличия вируса
(индикация)
⇓
определение типа вируса
(идентификация)
Способы культивирования вирусов
куриный эмбрион
культура клеток
организм лабораторного животного
⇓
обнаружение наличия вируса
(индикация)
⇓
определение типа вируса
(идентификация)
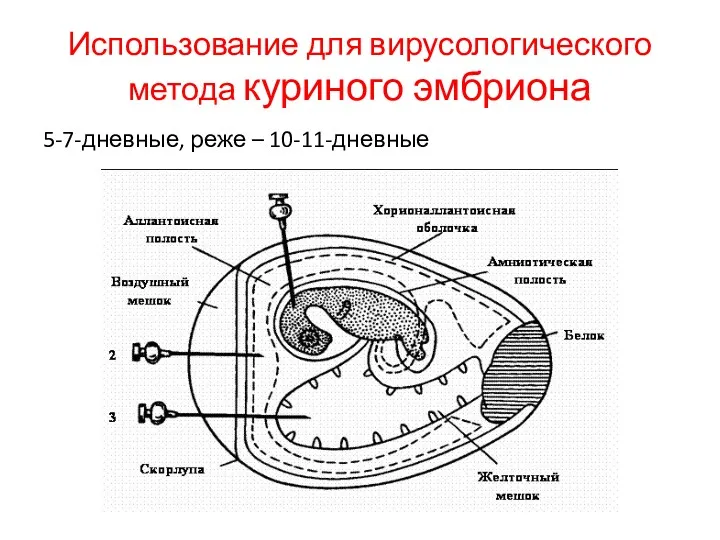
Использование для вирусологического метода куриного эмбриона
5-7-дневные, реже – 10-11-дневные
Использование для вирусологического метода куриного эмбриона
5-7-дневные, реже – 10-11-дневные

Основные способы заражения куриных эмбрионов
на хорион-аллантоисную оболочку
в хорион-аллантоисную полость
в полость желточного
Основные способы заражения куриных эмбрионов
на хорион-аллантоисную оболочку
в хорион-аллантоисную полость
в полость желточного

Обнаружение вирусов в курином эмбрионе
индикация:
гибель эмбриона
морфологические изменения эмбриона/оболочек
РГА с жидкостью из
Обнаружение вирусов в курином эмбрионе
индикация:
гибель эмбриона
морфологические изменения эмбриона/оболочек
РГА с жидкостью из

Использование культур клеток
Культуры клеток = соматические или эмбриональные клетки человека или
Использование культур клеток
Культуры клеток = соматические или эмбриональные клетки человека или

Использование культур клеток
Чаще – перевиваемые монослойные
индикация:
ЦПД (цитопатическое действие вирусов – любое
Использование культур клеток
Чаще – перевиваемые монослойные
индикация:
ЦПД (цитопатическое действие вирусов – любое

Первичные культуры клеток
получают из тканей (эмбриональных или нормальных) многоклеточных организмов. Такие
Первичные культуры клеток
получают из тканей (эмбриональных или нормальных) многоклеточных организмов. Такие

Перевиваемые культуры клеток
Перевиваемые = стабильные = готовят из опухолевых клеток, способных
Перевиваемые культуры клеток
Перевиваемые = стабильные = готовят из опухолевых клеток, способных

Преимущества перевиваемых культур клеток перед первичными:
продолжительность культивирования – десятки лет,
высокая
Преимущества перевиваемых культур клеток перед первичными:
продолжительность культивирования – десятки лет,
высокая

Полуперевиваемые культуры клеток
– диплоидные клетки различных тканей и органов, способные к
Полуперевиваемые культуры клеток
– диплоидные клетки различных тканей и органов, способные к

Условия культивирования клеток:
Питательные среды сложного состава (среда 199, Игла), сод-т источники
Условия культивирования клеток:
Питательные среды сложного состава (среда 199, Игла), сод-т источники

Обнаружение = индикация вирусов в культуре клеток
проводят на основе следующих
Обнаружение = индикация вирусов в культуре клеток
проводят на основе следующих
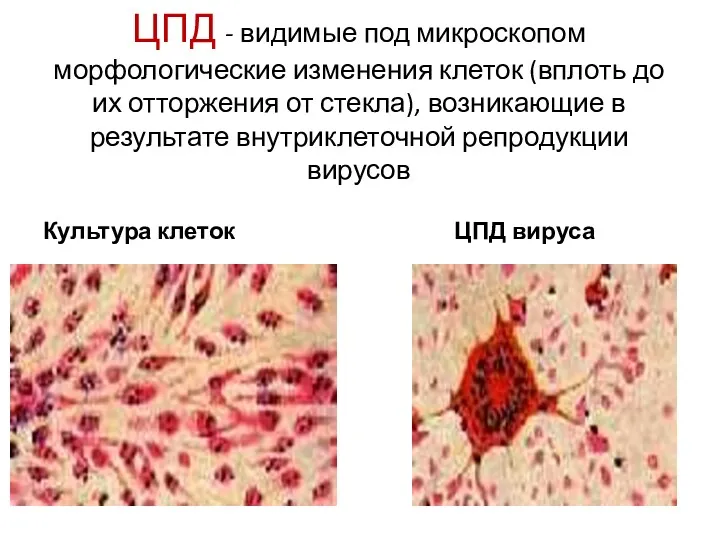
ЦПД - видимые под микроскопом морфологические изменения клеток (вплоть до их
ЦПД - видимые под микроскопом морфологические изменения клеток (вплоть до их

Виды ЦПД
округление и сморщивание клеток – пикорнавирусы,
нарастающая деструкция – герпесвирусы,
пролиферация (образование
Виды ЦПД
округление и сморщивание клеток – пикорнавирусы,
нарастающая деструкция – герпесвирусы,
пролиферация (образование
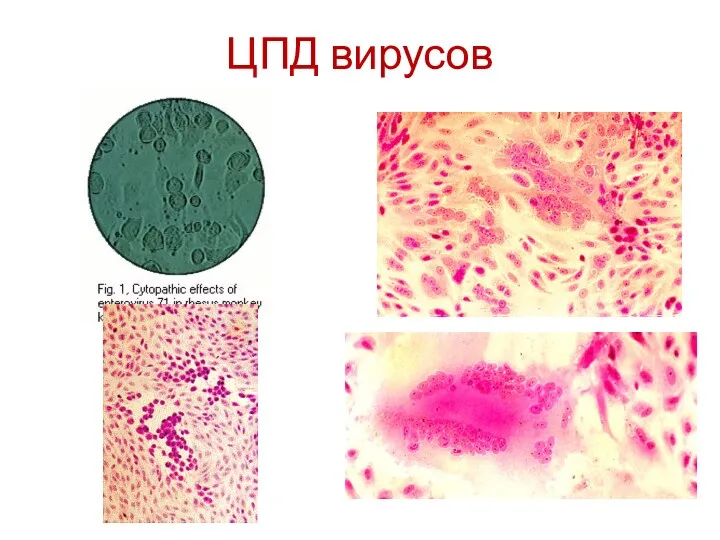
ЦПД вирусов
ЦПД вирусов

Включения
= скопление вирионов или отдельных их компонентов в цитоплазме или ядре
Включения
= скопление вирионов или отдельных их компонентов в цитоплазме или ядре
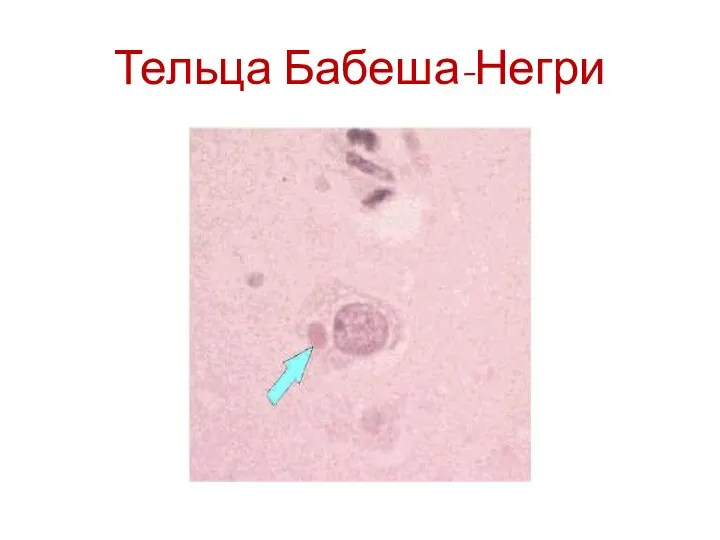
Тельца Бабеша-Негри
Тельца Бабеша-Негри

Бляшки, или “негативные” колонии
= ограниченные участки разрушенных вирусами клеток, культивируемых на
Бляшки, или “негативные” колонии
= ограниченные участки разрушенных вирусами клеток, культивируемых на

Реакция гемагглютинации (РГА)
основана на способности некоторых вирусов вызывать агглютинацию (склеивание) эритроцитов
Реакция гемагглютинации (РГА)
основана на способности некоторых вирусов вызывать агглютинацию (склеивание) эритроцитов
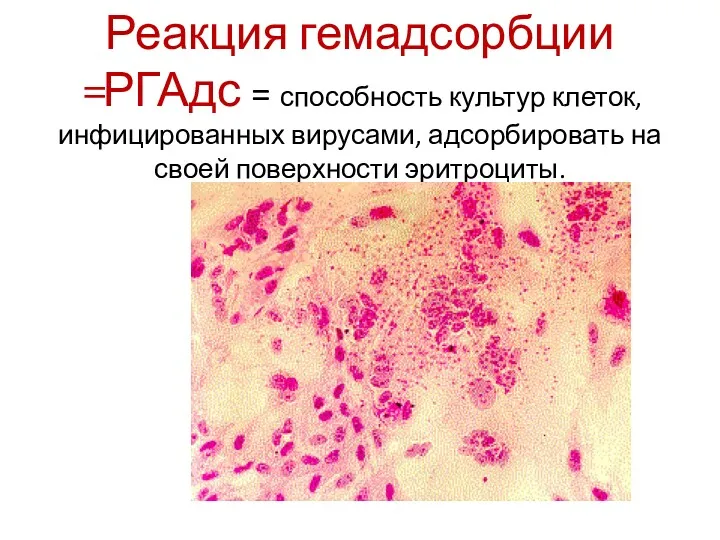
Реакция гемадсорбции =РГАдс = способность культур клеток, инфицированных вирусами, адсорбировать на
Реакция гемадсорбции =РГАдс = способность культур клеток, инфицированных вирусами, адсорбировать на
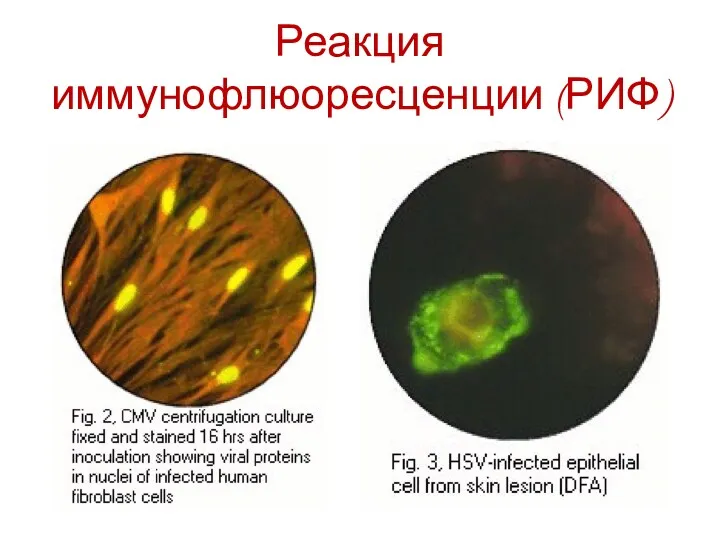
Реакция иммунофлюоресценции (РИФ)
Реакция иммунофлюоресценции (РИФ)

Использование лабораторных животных
взрослые или новорожденные белые мыши, хомяки, кролики, обезьяны
применяется для
Использование лабораторных животных
взрослые или новорожденные белые мыши, хомяки, кролики, обезьяны
применяется для

Способы заражения лабораторных животных
интраназально,
подкожно,
внутримышечно,
внутрибрюшинно,
интрацеребрально,
Способы заражения лабораторных животных
интраназально,
подкожно,
внутримышечно,
внутрибрюшинно,
интрацеребрально,

Обнаружение вируса при заражении лабораторных животных
обнаруживают вирус по:
- развитию видимых
Обнаружение вируса при заражении лабораторных животных
обнаруживают вирус по:
- развитию видимых

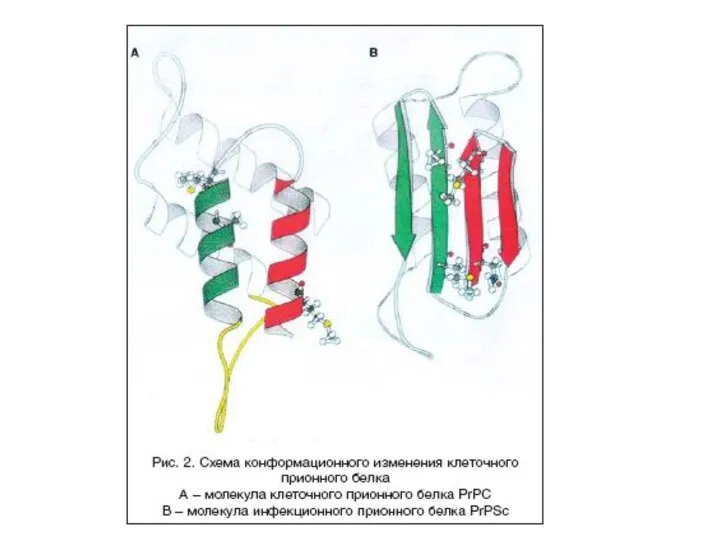
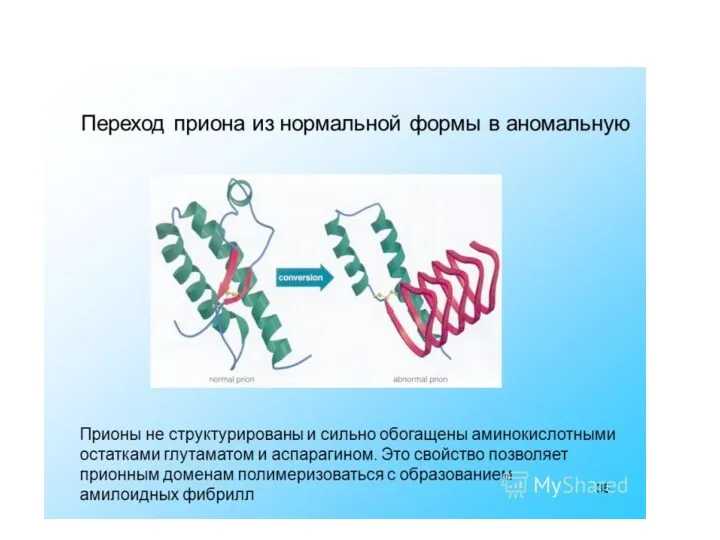
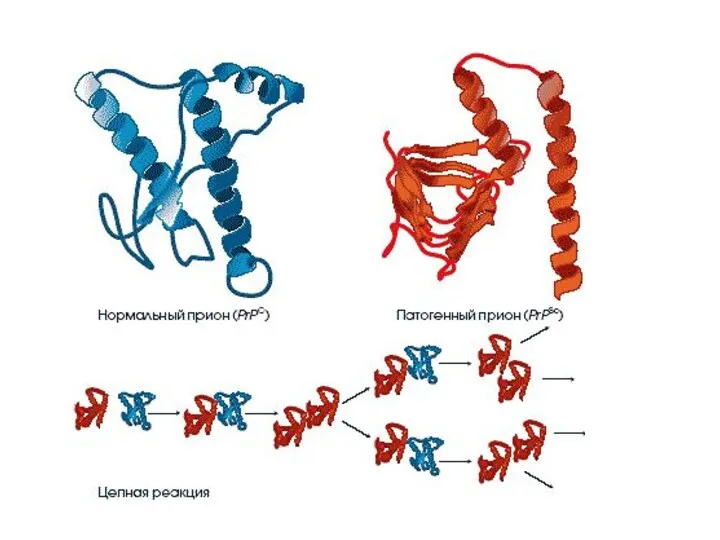
Прионы
– белковые молекулы, способные вызывать разрушение клеток организма человека и
Прионы
– белковые молекулы, способные вызывать разрушение клеток организма человека и

Прионы
Прионный белок может существовать в двух формах:
нормальная клеточная форма(РrPc) -
Прионы
Прионный белок может существовать в двух формах:
нормальная клеточная форма(РrPc) -

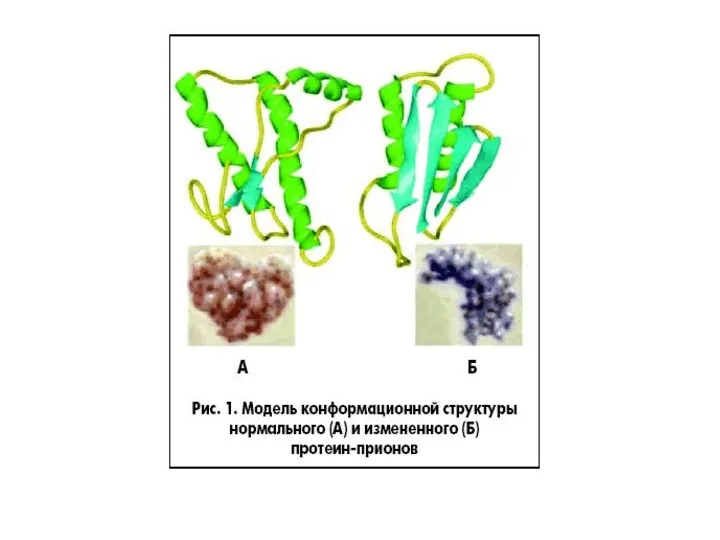
Прионы
инфекционная форма (PrPs) – характеризуется:
измененной вторичной и третичной структурой молекулы,
Прионы
инфекционная форма (PrPs) – характеризуется:
измененной вторичной и третичной структурой молекулы,
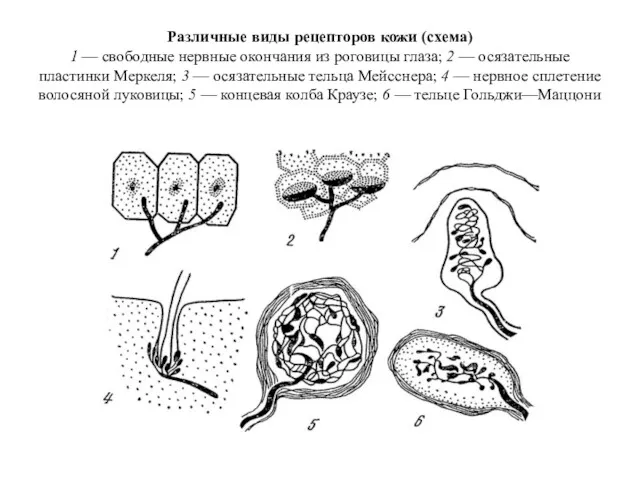 Различные виды рецепторов кожи
Различные виды рецепторов кожи Тест по теме Кровь.
Тест по теме Кровь.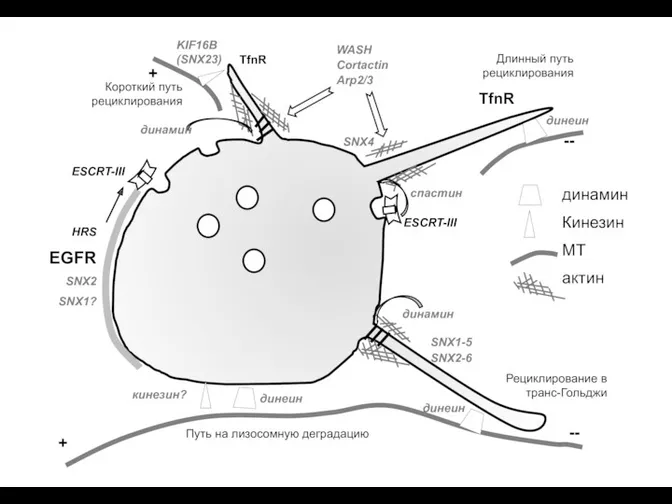 Везикулярный транспорт (эндосомы)
Везикулярный транспорт (эндосомы) Царство Растения
Царство Растения Однодольные растения
Однодольные растения Закономерности изменчивости. Наследственная изменчивость
Закономерности изменчивости. Наследственная изменчивость Перші мікроскопи
Перші мікроскопи Сон і сноведіння
Сон і сноведіння Популяция. Биологическая классификация
Популяция. Биологическая классификация Витамины
Витамины Фотосинтез як характерна особливість рослин
Фотосинтез як характерна особливість рослин Регуляция пищеварения. Заболевания органов пищеварения и их предупреждение. Биология 8 кл (Пасечник)
Регуляция пищеварения. Заболевания органов пищеварения и их предупреждение. Биология 8 кл (Пасечник) Тип Nemathelminthes (Круглые Черви)
Тип Nemathelminthes (Круглые Черви) Сорняки как индикаторы среды обитания
Сорняки как индикаторы среды обитания Рыбы, земноводные, пресмыкающиеся (фотографии)
Рыбы, земноводные, пресмыкающиеся (фотографии)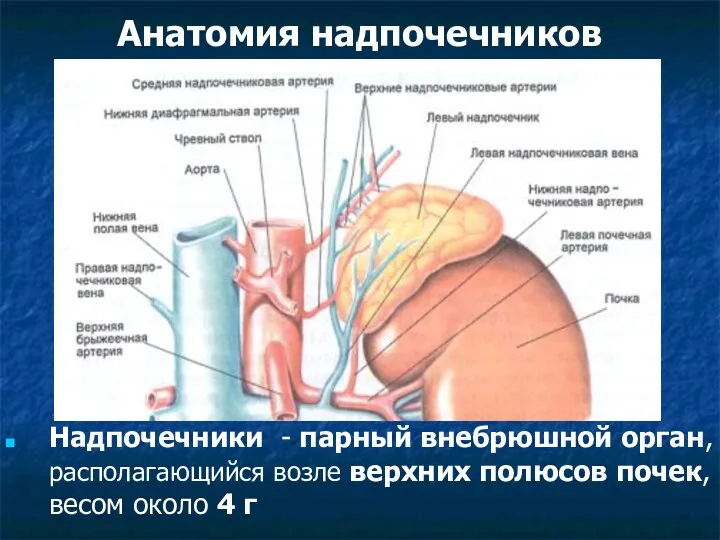 Анатомия надпочечников
Анатомия надпочечников Клеткалық биотехнология
Клеткалық биотехнология Хрящевые рыбы
Хрящевые рыбы Генетика пола. Наследование, сцепленное с полом
Генетика пола. Наследование, сцепленное с полом Презентация к уроку Анализаторы
Презентация к уроку Анализаторы Возрастное развитие сенсорной системы
Возрастное развитие сенсорной системы Размножение растений
Размножение растений Любите и берегите природу Алтайского края Внеклассное мероприятие для учащихся 7-8 классов.
Любите и берегите природу Алтайского края Внеклассное мероприятие для учащихся 7-8 классов. Лекарственные растения
Лекарственные растения Анатомия, как наука. Клетка
Анатомия, как наука. Клетка Апоптоз. Типы клеточной гибели
Апоптоз. Типы клеточной гибели Асимметрия функций больших полушарий. Моторика
Асимметрия функций больших полушарий. Моторика Презентация к уроку Хрящевые рыбы
Презентация к уроку Хрящевые рыбы