- Общие пути катаболизма

Содержание
- 2. Дыхательная цепь
- 3. Биологическое окисление Окислительное фосфорилирование
- 4. В пище человека нет готовых первичных доноров водорода, которые могут служить субстратами для дегидрогеназ. Они образуются
- 5. В ходе метаболизма У , Ж и Б образуются 2 центральных метаболита: 1) ПВК (пировиноградная кислота)
- 7. Образование пирувата из глюкозы
- 8. Распад жирных кислот
- 9. Различают специфические пути катаболизма (разные для разных классов веществ) и Общие пути катаболизма, которые являются единым
- 10. ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПИРУВАТА В МАТРИКСЕ МИТОХОНДРИЙ
- 11. Окислительное декарбоксилирование пирувата пируватдегидрогеназный мультиферментный комплекс: 3 фермента: пируватдегидрогеназа (декарбоксилирующая) - Е1-ТПФ, дигидролипоилацетилтрансфераза – Е2-ЛК, Дигидролипоилдегидрогеназа
- 12. 5 коферментов: 1)Тиаминдифосфат (ТДФ) с Е1, 2) Липоевая кислота (ЛК) с Е2,
- 13. 3) ФАД в виде простетической группы на Е3. 4) НАД+ 5) кофермент А
- 14. Е2-ЛК составляет ядро пируватдегидрогеназного комплекса, вокруг которого расположены пируватдегидрогеназа и дигидролипоилдегидрогеназа.
- 15. Суммарная реакция: Пируват + НАД+ + HS-KoA –>Ацетил-КоА + НАДН + Н+ + СO2.
- 16. Пируватдегидрогеназный комплекс
- 18. На I стадии пируват теряет свою карбоксильную группу в результате взаимодействия с ТПФ в составе активного
- 19. На II стадии оксиэтильная группа комплекса E1–ТПФ–СНОН–СН3 окисляется с образованием ацетильной группы, которая переносится на ЛК,
- 20. Этот фермент катализирует III стадию – перенос ацетильной группы на коэнзим КоА (HS-KoA) с образованием ацетил-КоА.
- 21. На IV стадии образуется окисленная форма ЛК из восстановленного комплекса Е2-ЛК. При участии фермента дигидролипоилдегидрогеназы (Е3-ФАД)
- 22. На V стадии восстановленный ФАДН2 дигидролипоилдегидрогеназы передает водороды на кофермент НАД с образованием НАДН + Н+.
- 23. Суммарная реакция, катализируемая пируватдегидрогеназным комплексом: Пируват + НАД+ + HS-KoA –>Ацетил-КоА + НАДН + Н+ +
- 24. Дыхательная цепь
- 25. Образовавшийся в процессе окислительного декарбоксилирования ацетил-КоА подвергается дальнейшему окислению с образованием СО2 и Н2О в цикле
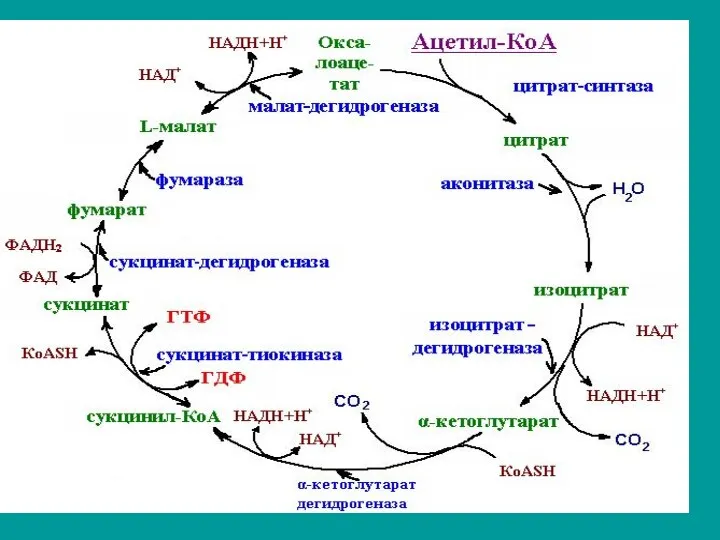
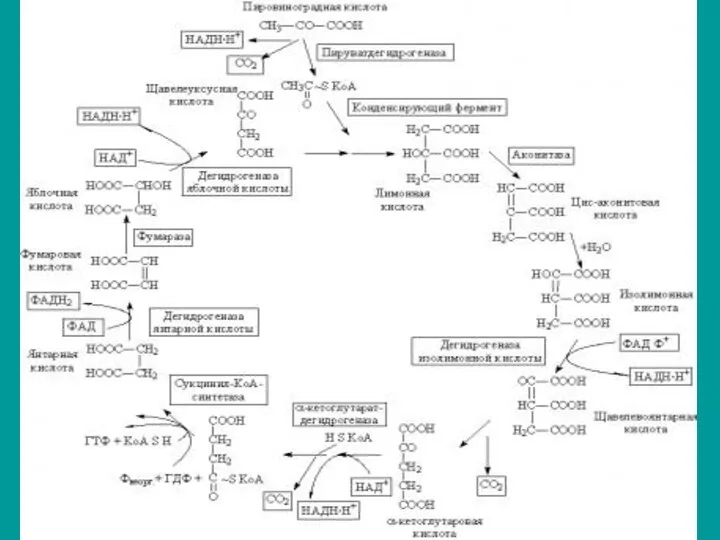
- 26. Цикл трикарбоновых кислот Полное «сгорание» как жирных кислот, так и углеводов требует окисления до СО2 и
- 27. ЦТК – цикл Кребса Сгорание происходит в цикле трикарбоновых кислот — циклом Кребса. Как и окислительное
- 28. Первая реакция Присоединение ацетильного остатка ацетилкофермента А к оксалоацетату с образованием трикарбоновой лимонной кислоты — цитрата.
- 29. Первая стадия Взаимодействие ацетилкофермента А с оксалоацетатом, катализируемое ферментом цитратсинтазой:
- 31. Вторая стадия Изомеризация цитрата в изоцитрат, катализируется аконитазой. Проходит через образование аконитата путем дегидратации цитрата и
- 33. Третья стадия 3. Окисление гидроксигруппы изоцитрата до карбонильной группы с помощью NAD+, сопровождается элиминацией карбоксильной группы
- 35. Четвертая стадия 4. Окислительное декарбоксилирование aльфа-кетоглутарата, катализируется aльфа‑кетоглутаратдегидрогеназным комплексом. Образуется сукцинилкофермент А и выделяется вторая молекула
- 37. Пятая стадия 5. Фосфорилирование GTP, сопряженное с гидролизом макроэргической тиоэфирной связи в сукцинилкоферменте А, катализируется сукцинатСоА
- 39. Шестая стадия 6. Превращение сукцината в фумарат, катализируется сукцинатдегидрогеназой, ( в составе комплекса II ЦПЭ с
- 41. Седьмая стадия 7. Гидратация двойной связи фумарата с образованием малата, катализируется фумарат- гидратазой:
- 43. Восьмая стадия 8. Окисление гидроксигруппы малата до кетогруппы, приводит к регенерации оксалоацетата, катализируется малатдегидрогеназой:
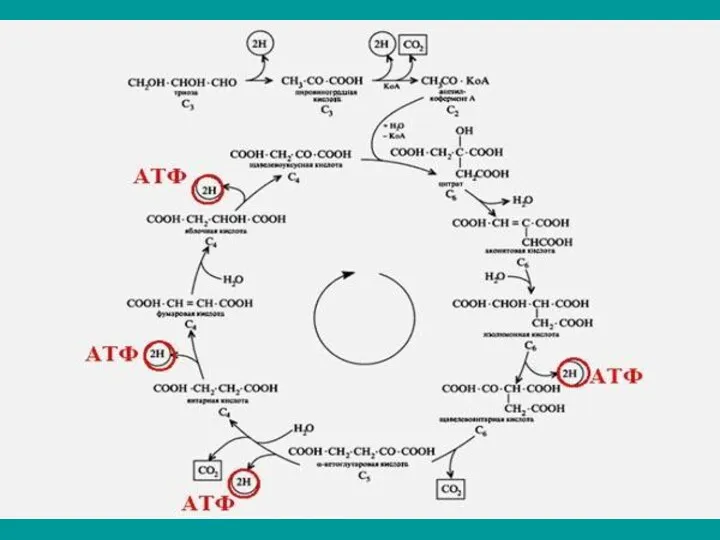
- 45. Значение ЦТК В ходе ЦТК восстанавливается до НАДH три молекулы НАД+, пара электронов посылается в комплексы
- 47. Энергетика ЦТК С учетом АТФ, образующихся в ЦПЭ при окислении НАДH2 и ФАДH2, сгорание ацетильного остатка
- 48. Роль ЦТК для анаболизма При стационарном функционировании ЦТК никакие компоненты цикла не расходуются. Некоторые компоненты ЦТК
- 50. Дыхательная цепь
- 54. Скачать презентацию
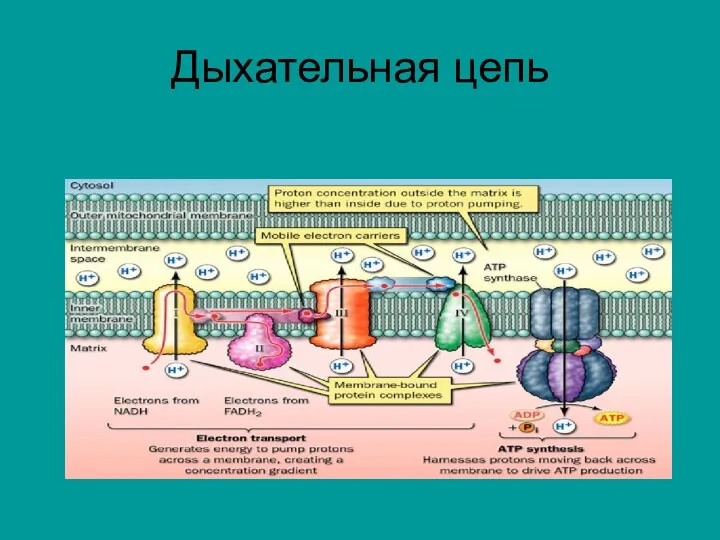
Дыхательная цепь
Дыхательная цепь
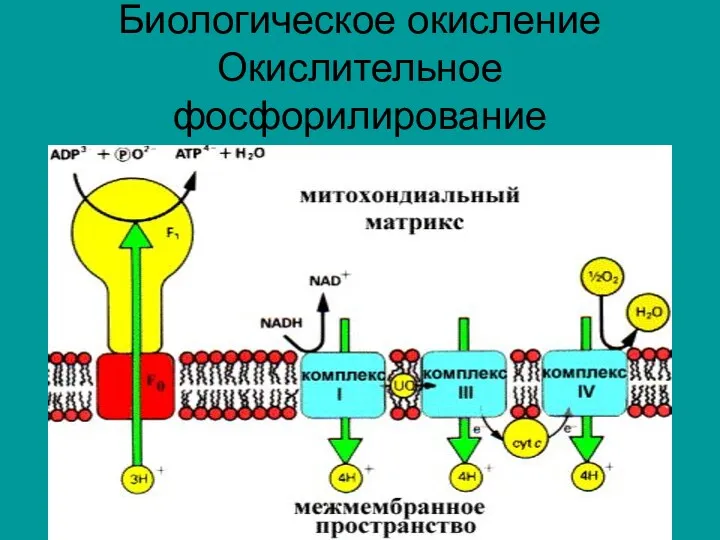
Биологическое окисление Окислительное фосфорилирование
Биологическое окисление Окислительное фосфорилирование

В пище человека нет готовых первичных доноров водорода, которые могут служить
В пище человека нет готовых первичных доноров водорода, которые могут служить

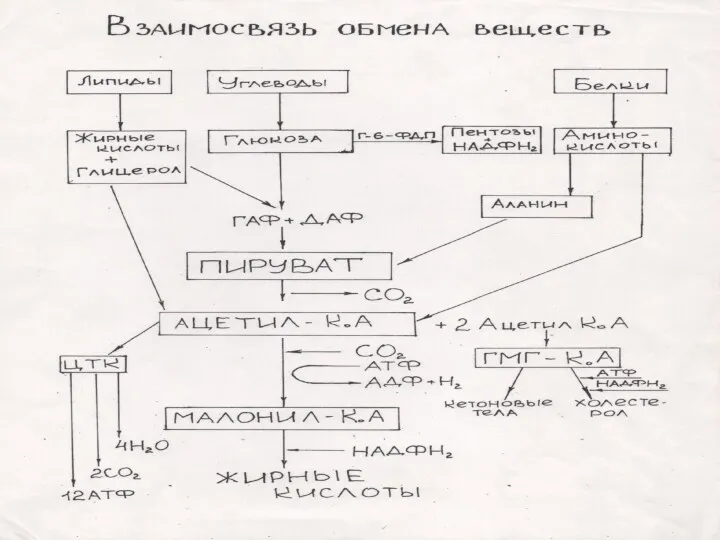
В ходе метаболизма У , Ж и Б образуются 2 центральных
В ходе метаболизма У , Ж и Б образуются 2 центральных
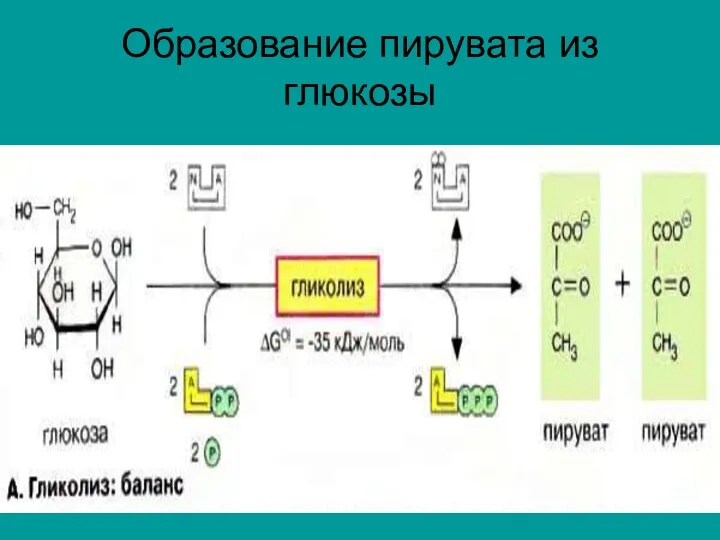
Образование пирувата из глюкозы
Образование пирувата из глюкозы
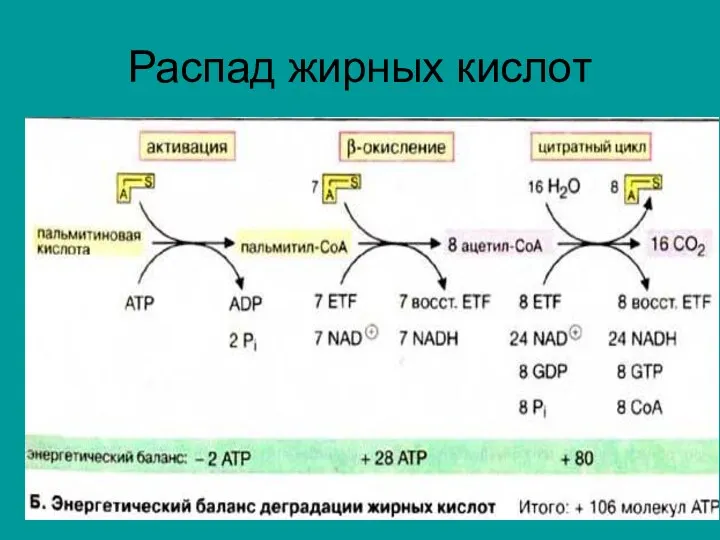
Распад жирных кислот
Распад жирных кислот

Различают специфические пути катаболизма (разные для разных классов веществ) и
Общие пути
Различают специфические пути катаболизма (разные для разных классов веществ) и
Общие пути
ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПИРУВАТА
В МАТРИКСЕ МИТОХОНДРИЙ
ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПИРУВАТА
В МАТРИКСЕ МИТОХОНДРИЙ

Окислительное декарбоксилирование пирувата
пируватдегидрогеназный мультиферментный комплекс:
3 фермента:
пируватдегидрогеназа (декарбоксилирующая) -
Окислительное декарбоксилирование пирувата
пируватдегидрогеназный мультиферментный комплекс:
3 фермента:
пируватдегидрогеназа (декарбоксилирующая) -

5 коферментов:
1)Тиаминдифосфат (ТДФ) с Е1,
2) Липоевая кислота (ЛК) с
5 коферментов:
1)Тиаминдифосфат (ТДФ) с Е1,
2) Липоевая кислота (ЛК) с

3) ФАД в виде простетической группы на Е3.
3) ФАД в виде простетической группы на Е3.

Е2-ЛК составляет ядро пируватдегидрогеназного комплекса, вокруг которого расположены пируватдегидрогеназа и
Е2-ЛК составляет ядро пируватдегидрогеназного комплекса, вокруг которого расположены пируватдегидрогеназа и

Суммарная реакция:
Пируват + НАД+ + HS-KoA
–>Ацетил-КоА +
Суммарная реакция:
Пируват + НАД+ + HS-KoA
–>Ацетил-КоА +
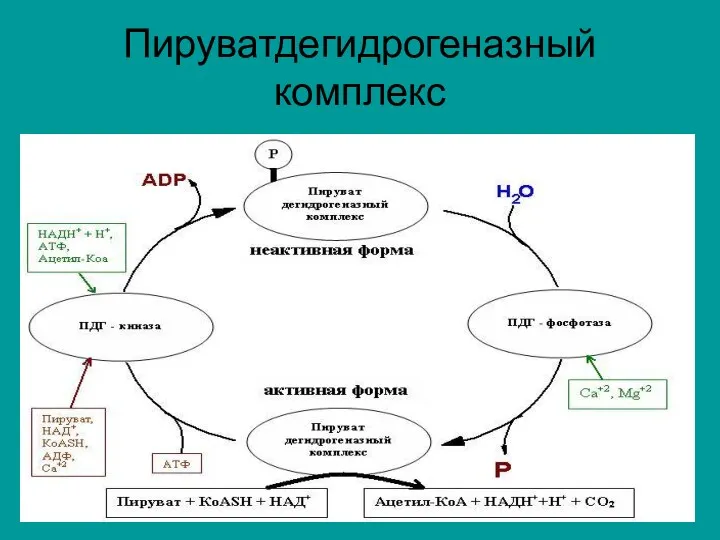
Пируватдегидрогеназный комплекс
Пируватдегидрогеназный комплекс
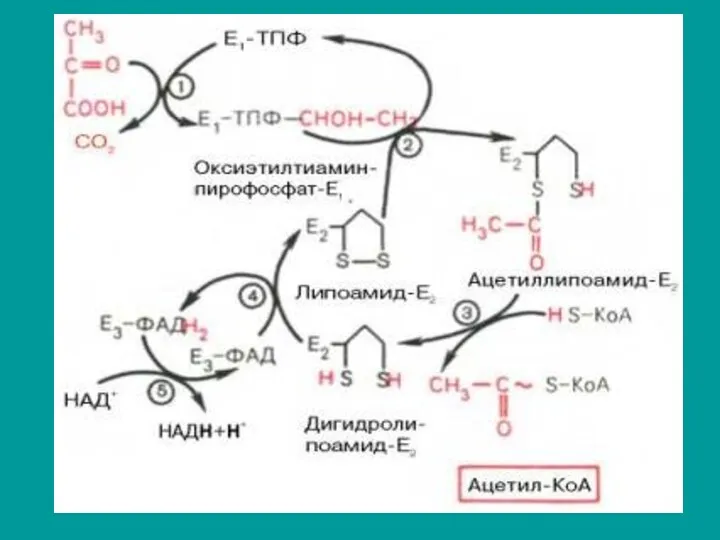

На I стадии пируват теряет свою карбоксильную группу в результате взаимодействия

На II стадии оксиэтильная группа комплекса E1–ТПФ–СНОН–СН3 окисляется с образованием ацетильной
На II стадии оксиэтильная группа комплекса E1–ТПФ–СНОН–СН3 окисляется с образованием ацетильной

Этот фермент катализирует III стадию – перенос ацетильной группы на
Этот фермент катализирует III стадию – перенос ацетильной группы на

На IV стадии образуется окисленная форма ЛК из восстановленного комплекса Е2-ЛК.
На IV стадии образуется окисленная форма ЛК из восстановленного комплекса Е2-ЛК.

На V стадии восстановленный ФАДН2 дигидролипоилдегидрогеназы передает водороды на кофермент
На V стадии восстановленный ФАДН2 дигидролипоилдегидрогеназы передает водороды на кофермент

Суммарная реакция, катализируемая пируватдегидрогеназным комплексом:
Пируват + НАД+ + HS-KoA
Суммарная реакция, катализируемая пируватдегидрогеназным комплексом:
Пируват + НАД+ + HS-KoA
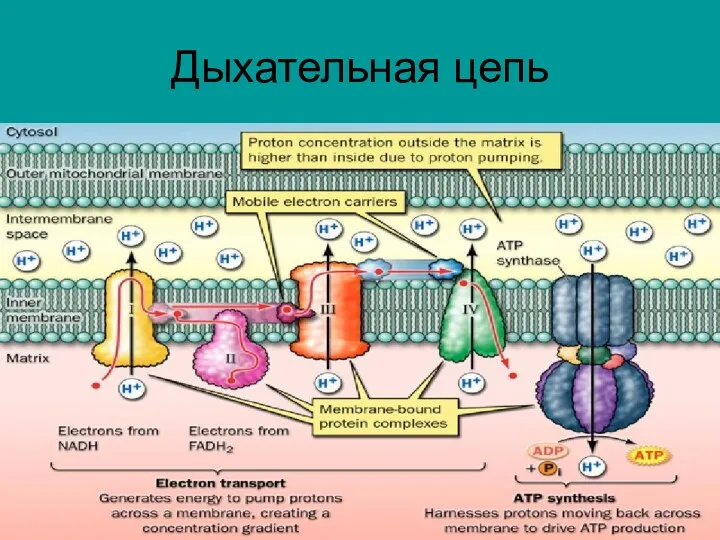
Дыхательная цепь
Дыхательная цепь

Образовавшийся в процессе окислительного декарбоксилирования ацетил-КоА подвергается дальнейшему окислению с образованием

Цикл трикарбоновых кислот
Полное «сгорание» как жирных кислот, так и углеводов требует
Цикл трикарбоновых кислот
Полное «сгорание» как жирных кислот, так и углеводов требует

ЦТК – цикл Кребса
Сгорание происходит в цикле трикарбоновых кислот — циклом
ЦТК – цикл Кребса
Сгорание происходит в цикле трикарбоновых кислот — циклом

Первая реакция
Присоединение ацетильного остатка ацетилкофермента А к оксалоацетату с образованием трикарбоновой
Первая реакция
Присоединение ацетильного остатка ацетилкофермента А к оксалоацетату с образованием трикарбоновой

Первая стадия
Взаимодействие ацетилкофермента А с оксалоацетатом, катализируемое ферментом цитратсинтазой:
Первая стадия
Взаимодействие ацетилкофермента А с оксалоацетатом, катализируемое ферментом цитратсинтазой:
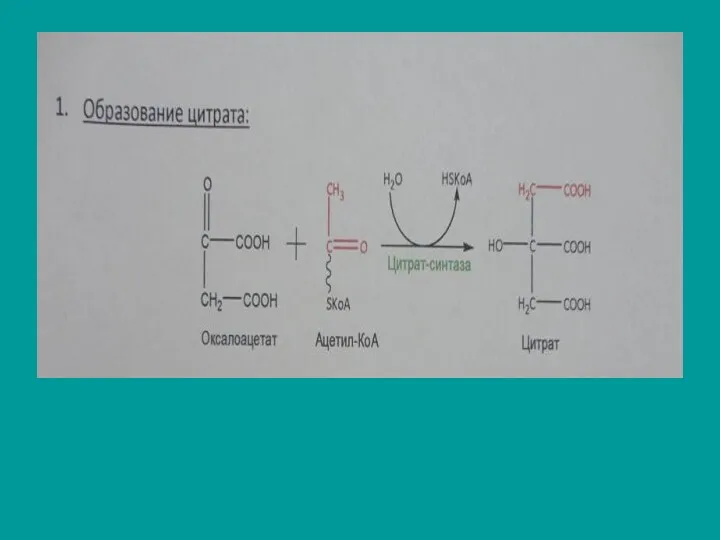

Вторая стадия
Изомеризация цитрата в изоцитрат, катализируется аконитазой.
Проходит через образование аконитата путем
Вторая стадия
Изомеризация цитрата в изоцитрат, катализируется аконитазой.
Проходит через образование аконитата путем
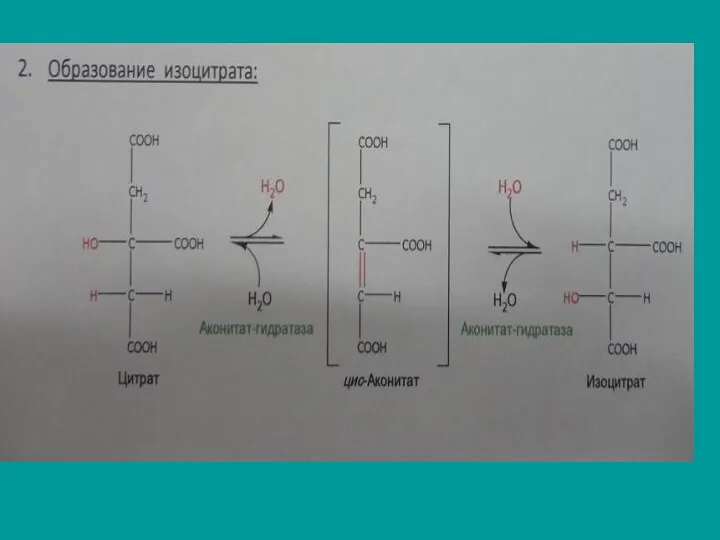

Третья стадия
3. Окисление гидроксигруппы изоцитрата до карбонильной группы с помощью NAD+,
Третья стадия
3. Окисление гидроксигруппы изоцитрата до карбонильной группы с помощью NAD+,
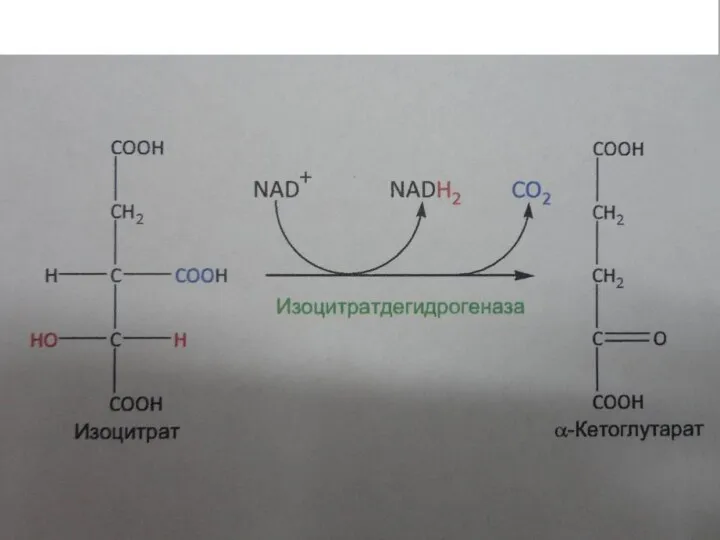

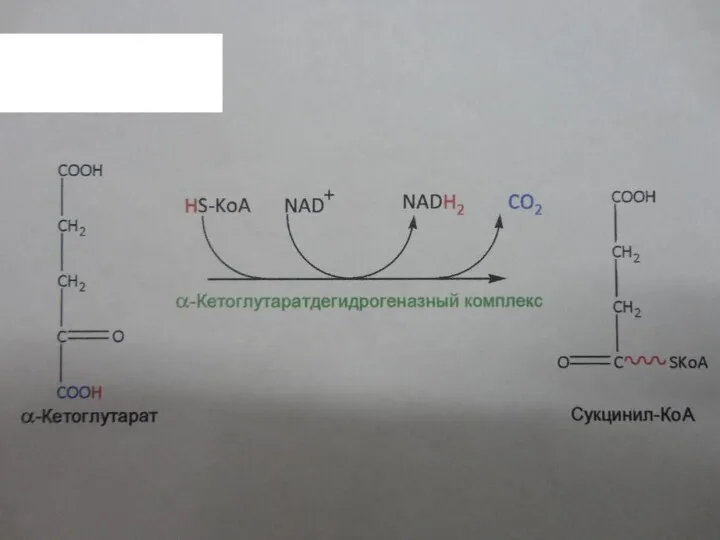
Четвертая стадия
4. Окислительное декарбоксилирование
aльфа-кетоглутарата, катализируется aльфа‑кетоглутаратдегидрогеназным комплексом. Образуется сукцинилкофермент А
Четвертая стадия
4. Окислительное декарбоксилирование
aльфа-кетоглутарата, катализируется aльфа‑кетоглутаратдегидрогеназным комплексом. Образуется сукцинилкофермент А

Пятая стадия
5. Фосфорилирование GTP, сопряженное с гидролизом макроэргической тиоэфирной связи в
Пятая стадия
5. Фосфорилирование GTP, сопряженное с гидролизом макроэргической тиоэфирной связи в


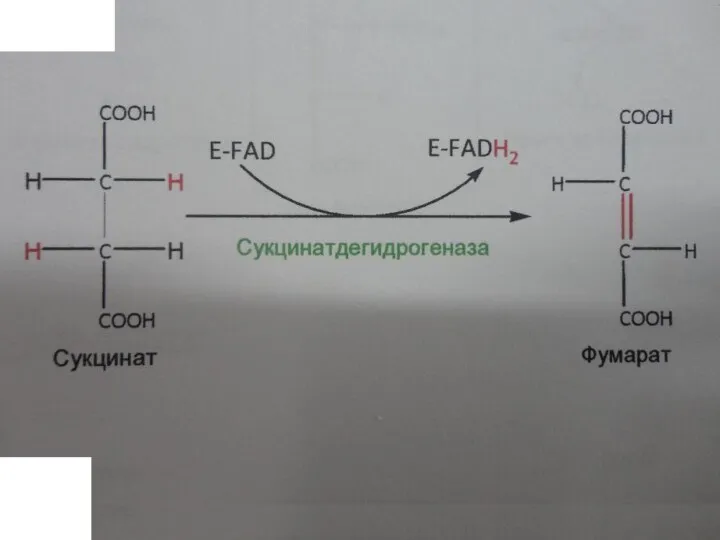
Шестая стадия
6. Превращение сукцината в фумарат, катализируется сукцинатдегидрогеназой,
( в
Шестая стадия
6. Превращение сукцината в фумарат, катализируется сукцинатдегидрогеназой,
( в

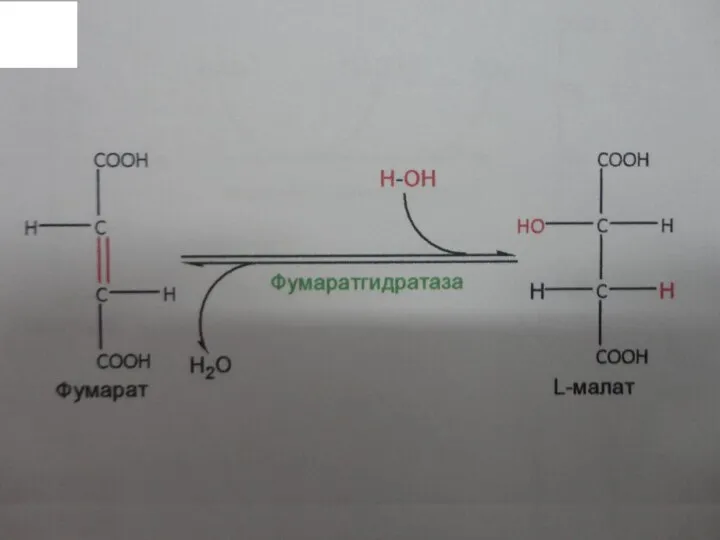
Седьмая стадия
7. Гидратация двойной связи фумарата с образованием малата, катализируется фумарат-
Седьмая стадия
7. Гидратация двойной связи фумарата с образованием малата, катализируется фумарат-

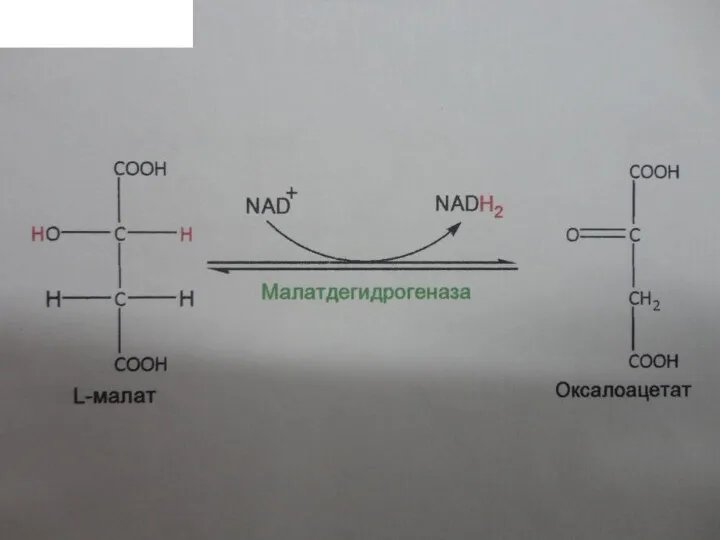
Восьмая стадия
8. Окисление гидроксигруппы малата до кетогруппы, приводит к регенерации оксалоацетата,
Восьмая стадия
8. Окисление гидроксигруппы малата до кетогруппы, приводит к регенерации оксалоацетата,

Значение ЦТК
В ходе ЦТК восстанавливается до НАДH три молекулы НАД+, пара
Значение ЦТК
В ходе ЦТК восстанавливается до НАДH три молекулы НАД+, пара

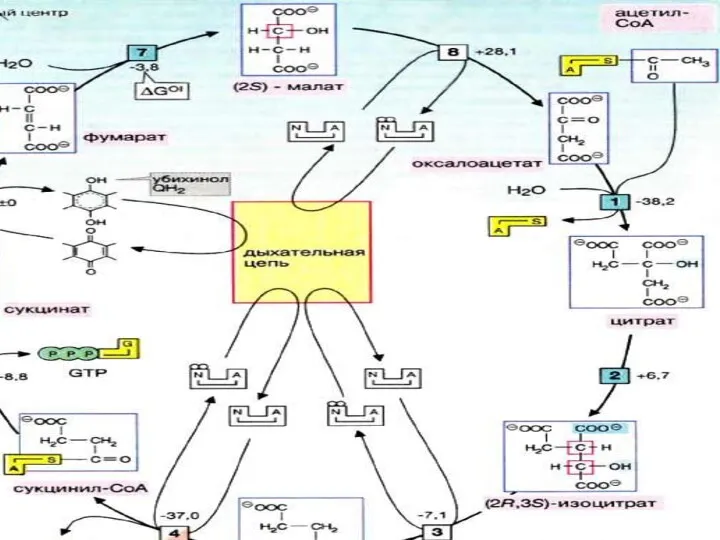
Энергетика ЦТК
С учетом АТФ, образующихся в ЦПЭ при окислении НАДH2 и
Энергетика ЦТК
С учетом АТФ, образующихся в ЦПЭ при окислении НАДH2 и

Роль ЦТК для анаболизма
При стационарном функционировании ЦТК никакие компоненты цикла не
Роль ЦТК для анаболизма
При стационарном функционировании ЦТК никакие компоненты цикла не
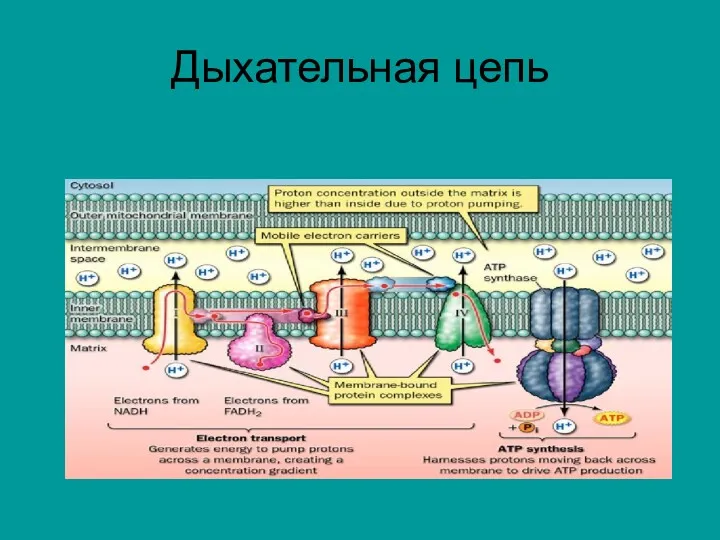
Дыхательная цепь
Дыхательная цепь
 Науки о человеке: анатомия, физиология, гигиена
Науки о человеке: анатомия, физиология, гигиена Биология оқыту әдістемесі ғылым ретінде. Биология оқыту әдістемесінің пәні
Биология оқыту әдістемесі ғылым ретінде. Биология оқыту әдістемесінің пәні Грегор Мендель. Закон единообразия гибридов
Грегор Мендель. Закон единообразия гибридов Микроскопические методы исследования морфологии микроорганизмов
Микроскопические методы исследования морфологии микроорганизмов Этапы эволюции человека
Этапы эволюции человека Как определить птицу
Как определить птицу Пути воздействия организмов на среду обитания
Пути воздействия организмов на среду обитания Організмовий рівень організації генетичної інформації. Взаємодія генів
Організмовий рівень організації генетичної інформації. Взаємодія генів Продуктивность кукурузы в зависимости от приемов выращивания в центральной зоне Краснодарского края
Продуктивность кукурузы в зависимости от приемов выращивания в центральной зоне Краснодарского края Сердечно-сосудистая система
Сердечно-сосудистая система Презентация по биологии Семейство крестоцветные
Презентация по биологии Семейство крестоцветные Бактериофаги. Строение бактериофагов
Бактериофаги. Строение бактериофагов Дикие и домашние животные
Дикие и домашние животные По грибы. Классный час, возраст: 7-10 лет
По грибы. Классный час, возраст: 7-10 лет Двигательные системы
Двигательные системы Индивидуальное развитие организмов (онтогенез)
Индивидуальное развитие организмов (онтогенез) Урок 8 класс. Осанка. Первая помощь при травмах скелета
Урок 8 класс. Осанка. Первая помощь при травмах скелета Характеристика класса птицы. Внешнее строение птиц
Характеристика класса птицы. Внешнее строение птиц Конспект открытого урока по теме : Опорно-двигательная система. Биология 8 класс.
Конспект открытого урока по теме : Опорно-двигательная система. Биология 8 класс. Царство животные
Царство животные Життя риб навесні. 2 клас
Життя риб навесні. 2 клас Анатомия женских половых органов (1)
Анатомия женских половых органов (1) V.A. Motsart hayoti va ijodi
V.A. Motsart hayoti va ijodi Гиповитаминозы животных
Гиповитаминозы животных Отряды насекомых
Отряды насекомых Презентация непрямой онтогенез
Презентация непрямой онтогенез В поисках витамина С. 2 класс
В поисках витамина С. 2 класс Гіпотези виникнення життя на землі
Гіпотези виникнення життя на землі