- Основные группы ионных каналов в зависимости от механизма их активации (инактивации)

Содержание
- 2. Мембранные рецепторы, активируемые медиаторами Медиаторы, выделяясь из пресинаптических нейронов в межклеточное пространство по механизму экзоцитоза, связываются
- 3. Мембранные рецепторы, активируемые медиаторами По механизму активации различных процессов (в том числе и изменения ионной проводимости)
- 4. Еще один тип мембранных рецепторов, активируемых медиаторами Фермент-связанные рецепторы представляют собой встроенные в мембрану молекулы ферментов
- 5. Еще один тип немембранных рецепторов, активируемых сигнальными молекулами Внутриклеточные рецепторы локализованы в цитоплазме или ядре клетки.
- 6. Ионотропные рецепторы Ионотропные рецепторы представляют собой лиганд-активируемые, или лиганд-зависимые ионные каналы и обеспечивают прямую синаптическую передачу.
- 7. Ионотропные рецепторы Важной особенностью ионотропного рецептора является то, что влияние медиатора на состояние ионного канала определяется
- 8. Подразделение ионотропных рецепторов по химическому составу Включают три подсемейства: 1) никотинподобные рецепторы, 2) ионотропнные глютаматные рецепторы,
- 9. Молекулярная структура ионотропных рецепторов (каналов) Трансмембранная конфигурация субъединиц и схематичная четвертичная структура ионотропных ионных каналов. L
- 10. Молекулярная структура ионотропных рецепторов (каналов) Трансмембранная конфигурация субъединиц и схематичная четвертичная структура ионотропных ионных каналов. пуриновые
- 11. Молекулярная структура ионотропных рецепторов (каналов) Трансмембранная конфигурация субъединиц и схематичная четвертичная структура ионотропных ионных каналов. четвертичная
- 12. Никотинподобные рецепторы В составе внеклеточной N-терминали имеют два цистеиновых остатка, которые формируют дисульфидный мостик, отделяющий высоко
- 13. Никотинподобные рецепторы Цистеинобразованная петля является характерной для всех никотинподобных рецепторов, включающих никотиновые ацетилхолиновые, - ионотропные гамма-аминомасляные
- 14. Эволюционное родство субъединиц никотинподобных рецепторов The nomenclature used to describe each receptor subunit is RRRsS#, where
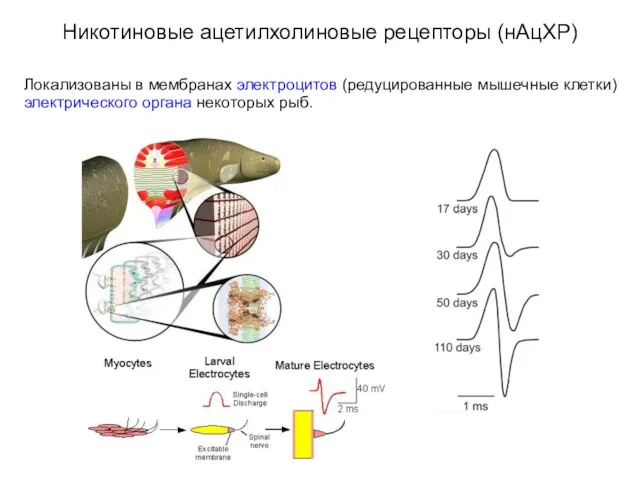
- 15. Никотиновые ацетилхолиновые рецепторы (нАцХР) Впервые выделены из мембран электроцитов (редуцированные мышечные клетки) электрического органа ската Torpedo.
- 16. Никотиновые ацетилхолиновые рецепторы (нАцХР) Локализованы в мембранах электроцитов (редуцированные мышечные клетки) электрического органа некоторых рыб.
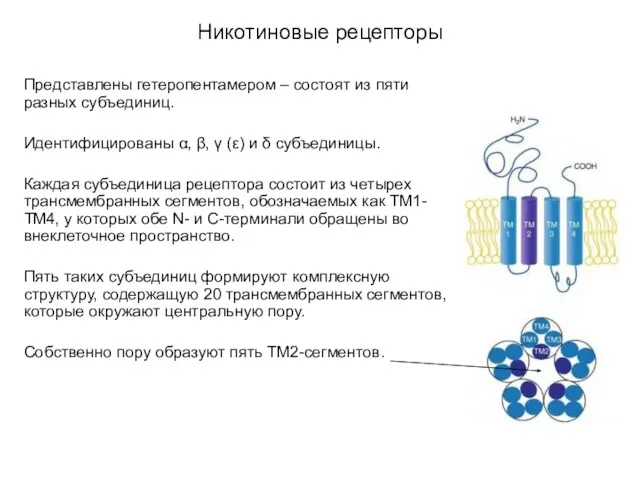
- 17. Никотиновые рецепторы Представлены гетеропентамером – состоят из пяти разных субъединиц. Идентифицированы α, β, γ (ε) и
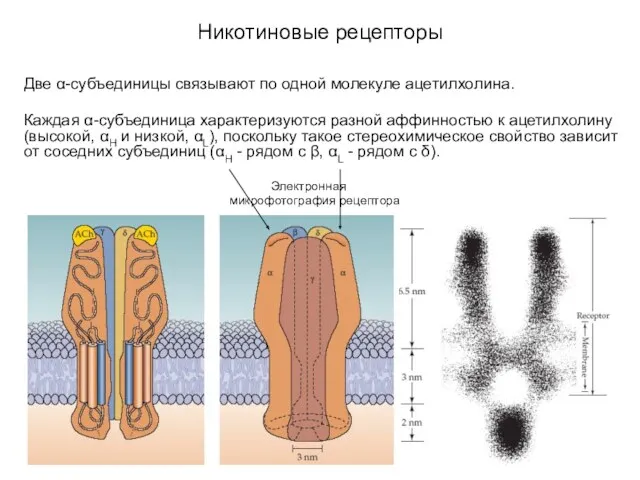
- 18. Никотиновые рецепторы Две α-субъединицы связывают по одной молекуле ацетилхолина. Каждая α-субъединица характеризуются разной аффинностью к ацетилхолину
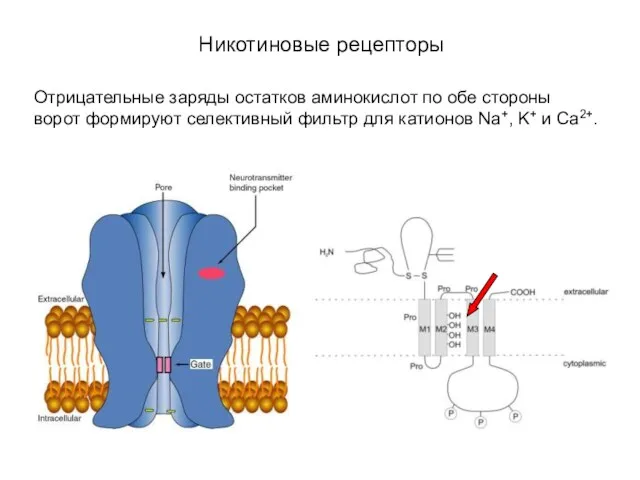
- 19. Никотиновые рецепторы Отрицательные заряды остатков аминокислот по обе стороны ворот формируют селективный фильтр для катионов Na+,
- 20. Фосфорилирование никотиновых рецепторов Субъединицы многих ионотропных рецепторов (в том числе и нАцХР) имеют участки фосфорилирования. нАцХР
- 21. Десенситизация никотиновых рецепторов Функциональное значение фосфорилирования рецепторов состоит в увеличении степени десенситизации. Десенситизация ионотропных рецепторов состоит
- 22. Постсинаптическая модуляция: ауторегуляция Отрицательная ауторегуляция Десенситизация (десенсибилизация) рецепторов при длительном воздействии медиатора. Пример: Если постсинаптическая мембрана
- 23. Субъединицы никотиновых рецепторов Методами гибридизации ДНК и генной инженерии всего идентифицировано 10 вариаций α-, четыре -
- 24. Субъединицы никотиновых рецепторов 17 субъединиц подразделяют на один мышечный и три нейронных (два в ЦНС и
- 25. Субъединицы никотиновых рецепторов Примеры рецепторов 4-х подсемейств Pharmacology, (Rang, Dale, Ritter & Moore, 5th ed., Churchill
- 26. Субъединицы никотиновых рецепторов The nomenclature used to describe each receptor subunit is RRRsS#, where RRR represent
- 27. Субъединицы никотиновых рецепторов
- 28. Ионотропные ГАМК-рецепторы (А и С типов) Гетерометрические пентамеры, состоят из 19 различных субъединиц: 6 α, 4
- 29. Ионотропные ГАМКА-рецепторы Наиболее типичный в мозге ГАМКА рецептор состоит из 2 α-, 2 β- и 1
- 30. Ионотропные ГАМКА-рецепторы Бензодиазепины увеличивают эффективность связывания ГАМК с рецептором Барбитураты увеличивают время открытого состояния Cl--канала Нейроактивные
- 31. Ионотропные ГАМКС-рецепторы ГАМКС рецепторы нечувствительны к агонистам ГАМКА рецепторов – к бензодиазепинам, барбитуратам и нейроактивным стероидам,
- 32. Глициновые рецепторы Сходны по структуре с ионотропными ГАМК рецепторами и составлены из гомологичных α- и β-субъединиц
- 33. Субъединицы ГАМК и глициновых рецепторов
- 34. Глициновые рецепторы При биохимической очистке из ткани вместе с белком глицинового рецептора выделяется также полипептид гефирин,
- 35. Другие рецепторы, управляющие Cl--проводимостью У беспозвоночных открыты ацетилхолиновые, серотониновые, глутаматные и гистаминовые ионотропные рецепторы, управляющие хлорным
- 36. !!! ГАМК является возбуждающим медиатором Рецептор EXP-1 круглого червя С. elegans связывается с ГАМК, но вызывает
- 37. !!! ГАМК является возбуждающим медиатором У позвоночных (справа) возбуждающее действие ГАМК определяется анионной проводимостью, когда равновесный
- 38. Субъединицы серотониновых рецепторов
- 39. Серотониновые ионотропные рецепторы (5-НТ3) Структура близка к структуре нАцХР. Идентифицировано две субъединицы 5-НТ3-А и 5-НТ3-В. Синтезированные
- 40. Серотониновые ионотропные рецепторы (5-НТ3) При активации 5-НТ3-рецепторы обеспечивают проводимость моновалентных катионов (Na+ и K+). Также как
- 41. Ионотропные глютаматные рецепторы В 1970-х г.г. с использованием различных фармакологических агентов ионотропные глютаматные рецепторы были разделены
- 42. Ионотропные глютаматные рецепторы В 1970-х г.г. с использованием различных фармакологических агентов ионотропные глютаматные рецепторы были разделены
- 43. Ионотропные глютаматные рецепторы Конкурентным антагонистом всех глютаматных рецепторов является 2-амино-5-фосфо-валериановая кислота (D-AP5 или APV, англ., 2-Amino-5-Phospho(5
- 44. Молекулярная структура ионотропных глютаматных рецепторов Глютаматные рецепторы происходят от потенциал-зависимых К+-каналов 2ТМ семейства. Субъединица ионотропных глютаматных
- 45. Субъединицы глютаматных рецепторов
- 46. Молекулярная структура ионотропных глютаматных рецепторов Q/R участок на сегменте ТМ2 в составе разных субъединиц глютаматных рецепторов
- 47. Молекулярная структура ионотропных глютаматных рецепторов Схема связывания каината с глютаматным рецептором. Сегменты ТМ1-ТМ4 обозначены как 1-4.
- 48. Эволюционное родство ионотропных глютаматных рецепторов Один из ионотропных глютаматных рецепторов (GluR-К1) впервые был клонирован в 1989
- 49. Субъединицы глютаматных рецепторов
- 50. Эволюционное родство ионотропных глютаматных рецепторов NMDAR1–глицин-связывающая NMDAR2D NMDAR2C NMDAR2B NMDAR2A-глютамат-связывающие субъединицы NMDA рецепторов; КА2 КА1 GluR-5
- 51. АМРА рецепторы AMPA рецептор, состоящий только из GluR1 или GluR3 (или их комбинации) с глютаминовым (Q)
- 52. АМРА рецепторы Flip/Flop – участок из 38 аминокислот, который представлен во всех субъединицах двумя вариациями flip-
- 53. Каиновые рецепторы NMDAR1 – глицин-связывающая NMDAR2D NMDAR2C NMDAR2B NMDAR2A, - глютамат-связывающие субъединицы NMDA рецепторов; КА2 КА1
- 54. АМРА и каиновые рецепторы КА2 КА1 GluR-5 GluR-7 GluR-6– субъединицы каиновых рецепторов; GluR-1 GluR-4 GluR-3 GluR-2–
- 55. NMDA рецепторы NMDA рецепторы состоят из комбинации каких-либо трех глютамат-связывающих субъединиц (из четырех возможных (NR2A, NR2B,
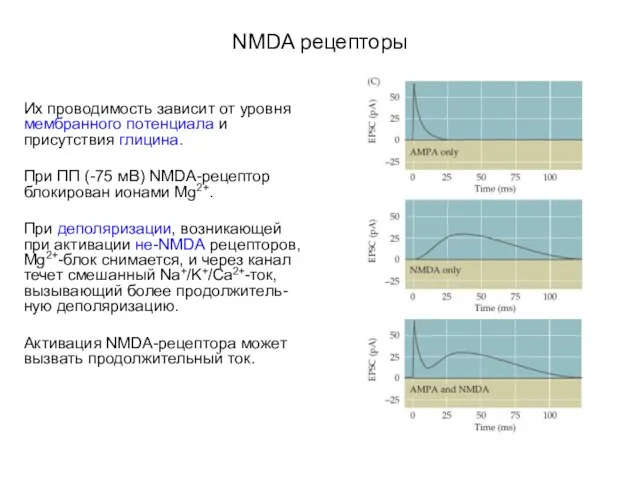
- 56. NMDA рецепторы Их проводимость зависит от уровня мембранного потенциала и от глицина. При потенциале покоя (-75
- 57. NMDA рецепторы Их проводимость зависит от уровня мембранного потенциала и присутствия глицина. При ПП (-75 мВ)
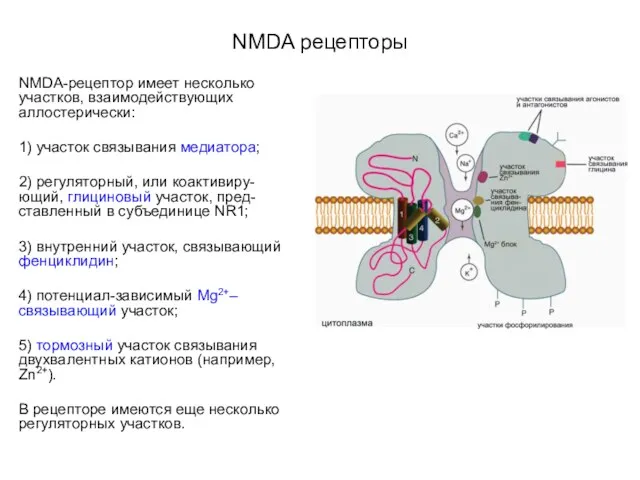
- 58. NMDA рецепторы NMDA-рецептор имеет несколько участков, взаимодействующих аллостерически: 1) участок связывания медиатора; 2) регуляторный, или коактивиру-ющий,
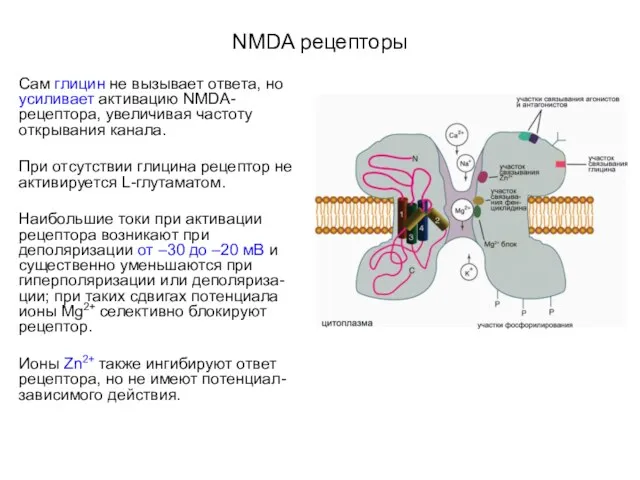
- 59. NMDA рецепторы Сам глицин не вызывает ответа, но усиливает активацию NMDA-рецептора, увеличивая частоту открывания канала. При
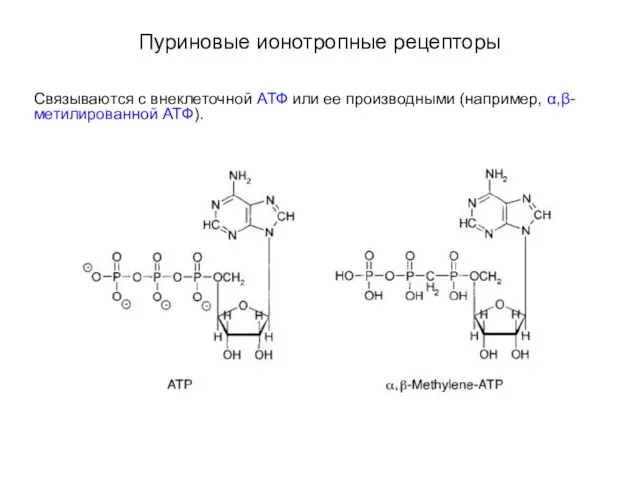
- 60. Пуриновые ионотропные рецепторы Связываются с внеклеточной АТФ или ее производными (например, α,β-метилированной АТФ).
- 61. Пуриновые ионотропные рецепторы Связываются с внеклеточной АТФ или ее производными (например, α,β-метилированной АТФ). АТФ-связывающие рецепторы подразделяют
- 62. Разнообразие пуриновых ионотропных рецепторов
- 63. Пуриновые ионотропные рецепторы Все субъединицы P2X рецепторов имеют структурную топологию, сходную с фрагментом потенциал-зависимых К+IR-каналов, образующим
- 65. Скачать презентацию
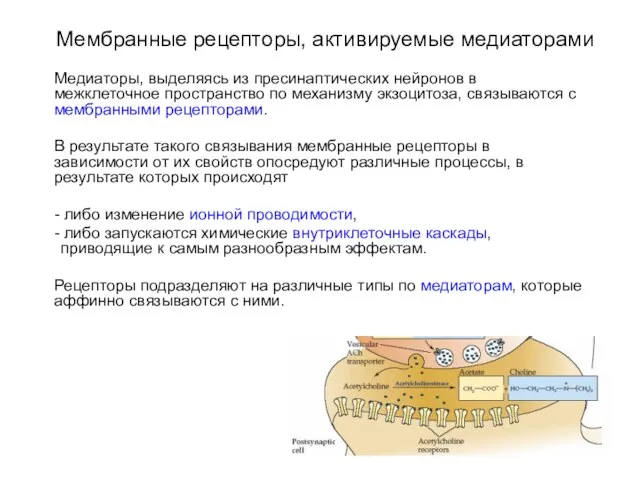
Мембранные рецепторы, активируемые медиаторами
Медиаторы, выделяясь из пресинаптических нейронов в межклеточное пространство
Мембранные рецепторы, активируемые медиаторами
Медиаторы, выделяясь из пресинаптических нейронов в межклеточное пространство
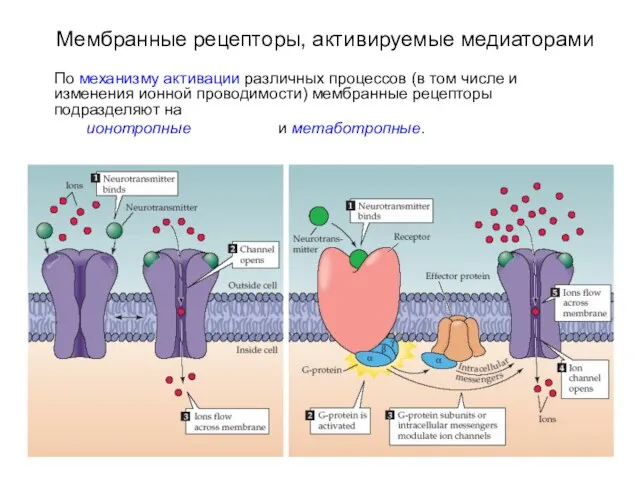
Мембранные рецепторы, активируемые медиаторами
По механизму активации различных процессов (в том числе
Мембранные рецепторы, активируемые медиаторами
По механизму активации различных процессов (в том числе
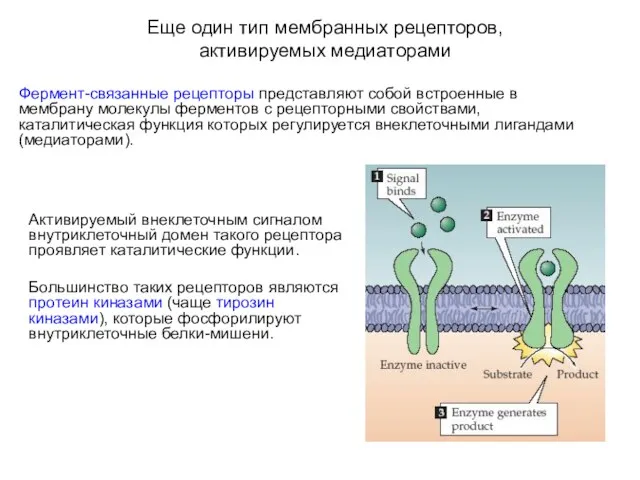
Еще один тип мембранных рецепторов,
активируемых медиаторами
Фермент-связанные рецепторы представляют собой встроенные в
Еще один тип мембранных рецепторов,
активируемых медиаторами
Фермент-связанные рецепторы представляют собой встроенные в

Еще один тип немембранных рецепторов,
активируемых сигнальными молекулами
Внутриклеточные рецепторы локализованы в цитоплазме
Еще один тип немембранных рецепторов,
активируемых сигнальными молекулами
Внутриклеточные рецепторы локализованы в цитоплазме
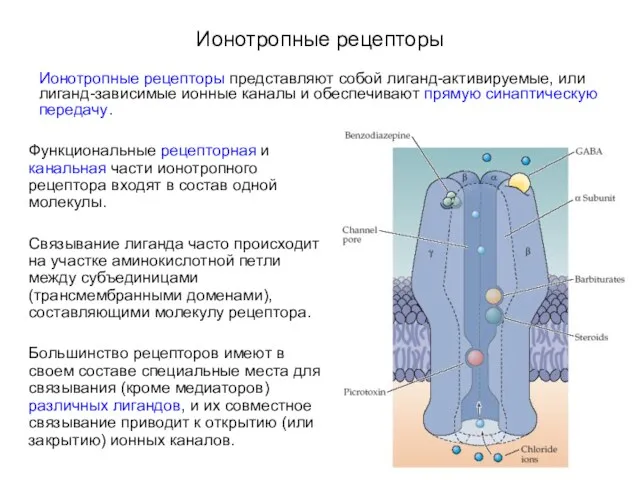
Ионотропные рецепторы
Ионотропные рецепторы представляют собой лиганд-активируемые, или лиганд-зависимые ионные каналы и
Ионотропные рецепторы
Ионотропные рецепторы представляют собой лиганд-активируемые, или лиганд-зависимые ионные каналы и
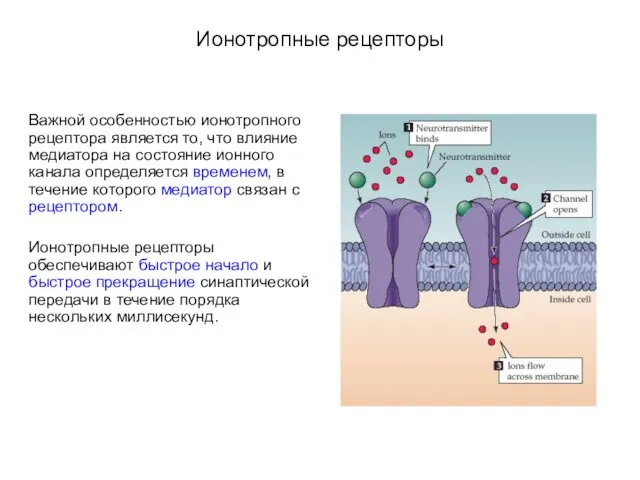
Ионотропные рецепторы
Важной особенностью ионотропного рецептора является то, что влияние медиатора на
Ионотропные рецепторы
Важной особенностью ионотропного рецептора является то, что влияние медиатора на
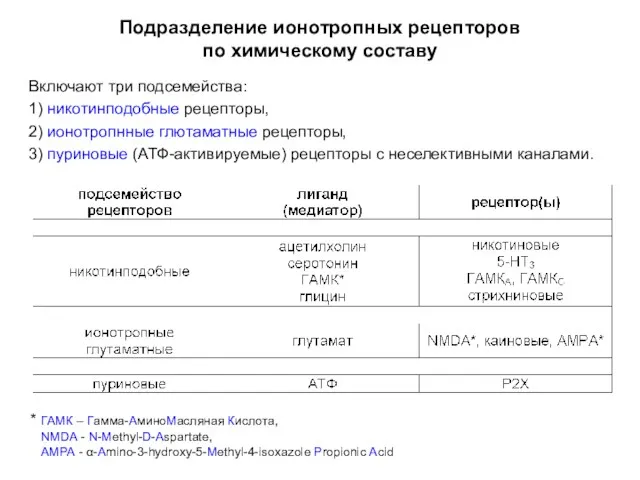
Подразделение ионотропных рецепторов
по химическому составу
Включают три подсемейства:
1) никотинподобные рецепторы,
2) ионотропнные глютаматные
Подразделение ионотропных рецепторов
по химическому составу
Включают три подсемейства:
1) никотинподобные рецепторы,
2) ионотропнные глютаматные

Молекулярная структура ионотропных рецепторов (каналов)
Трансмембранная конфигурация субъединиц и схематичная четвертичная структура
Молекулярная структура ионотропных рецепторов (каналов)
Трансмембранная конфигурация субъединиц и схематичная четвертичная структура
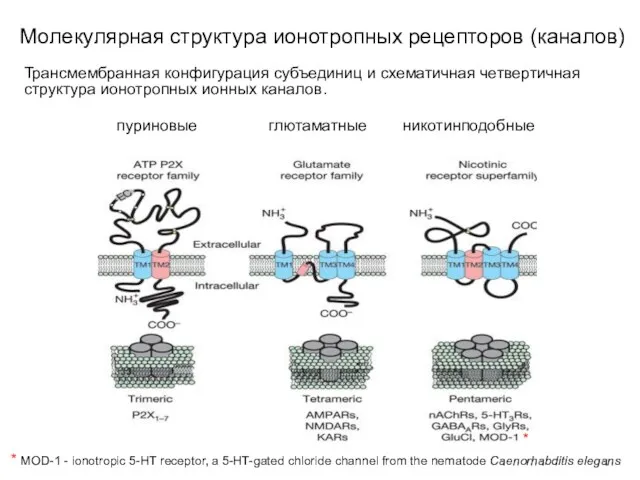
Молекулярная структура ионотропных рецепторов (каналов)
Трансмембранная конфигурация субъединиц и схематичная четвертичная структура
Молекулярная структура ионотропных рецепторов (каналов)
Трансмембранная конфигурация субъединиц и схематичная четвертичная структура
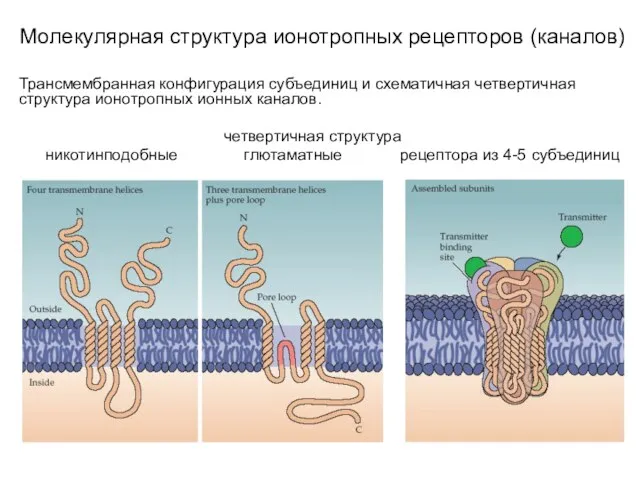
Молекулярная структура ионотропных рецепторов (каналов)
Трансмембранная конфигурация субъединиц и схематичная четвертичная структура
Молекулярная структура ионотропных рецепторов (каналов)
Трансмембранная конфигурация субъединиц и схематичная четвертичная структура
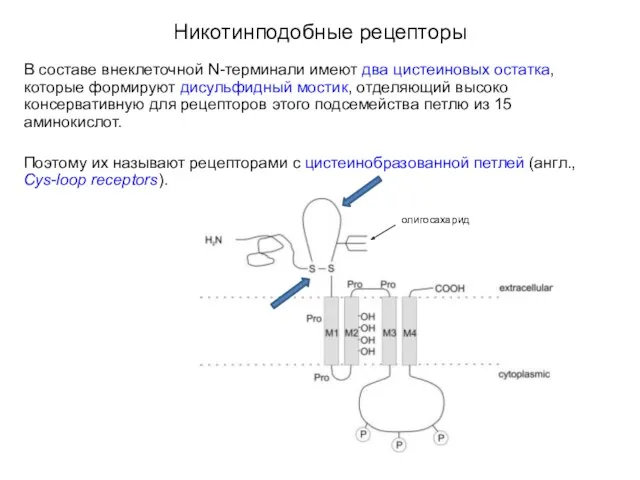
Никотинподобные рецепторы
В составе внеклеточной N-терминали имеют два цистеиновых остатка, которые
Никотинподобные рецепторы
В составе внеклеточной N-терминали имеют два цистеиновых остатка, которые
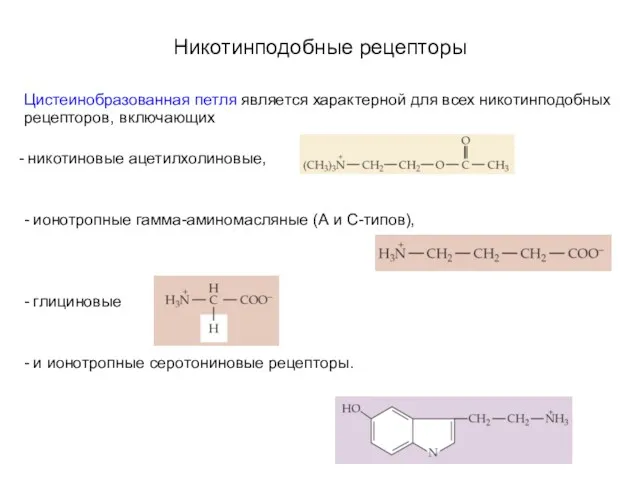
Никотинподобные рецепторы
Цистеинобразованная петля является характерной для всех никотинподобных рецепторов, включающих
Никотинподобные рецепторы
Цистеинобразованная петля является характерной для всех никотинподобных рецепторов, включающих
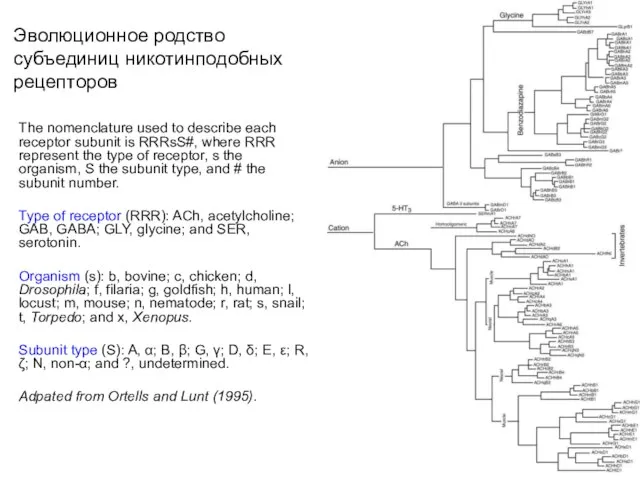
Эволюционное родство субъединиц никотинподобных рецепторов
The nomenclature used to describe each receptor
Эволюционное родство субъединиц никотинподобных рецепторов
The nomenclature used to describe each receptor
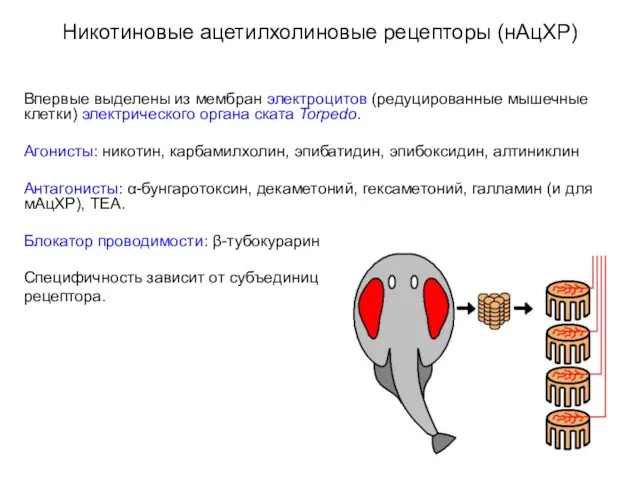
Никотиновые ацетилхолиновые рецепторы (нАцХР)
Впервые выделены из мембран электроцитов (редуцированные мышечные клетки)
Никотиновые ацетилхолиновые рецепторы (нАцХР)
Впервые выделены из мембран электроцитов (редуцированные мышечные клетки)
Никотиновые ацетилхолиновые рецепторы (нАцХР)
Локализованы в мембранах электроцитов (редуцированные мышечные клетки) электрического
Никотиновые ацетилхолиновые рецепторы (нАцХР)
Локализованы в мембранах электроцитов (редуцированные мышечные клетки) электрического
Никотиновые рецепторы
Представлены гетеропентамером – состоят из пяти разных субъединиц.
Идентифицированы α, β,
Никотиновые рецепторы
Представлены гетеропентамером – состоят из пяти разных субъединиц.
Идентифицированы α, β,
Никотиновые рецепторы
Две α-субъединицы связывают по одной молекуле ацетилхолина.
Каждая α-субъединица характеризуются разной
Никотиновые рецепторы
Две α-субъединицы связывают по одной молекуле ацетилхолина.
Каждая α-субъединица характеризуются разной
Никотиновые рецепторы
Отрицательные заряды остатков аминокислот по обе стороны ворот формируют селективный
Никотиновые рецепторы
Отрицательные заряды остатков аминокислот по обе стороны ворот формируют селективный
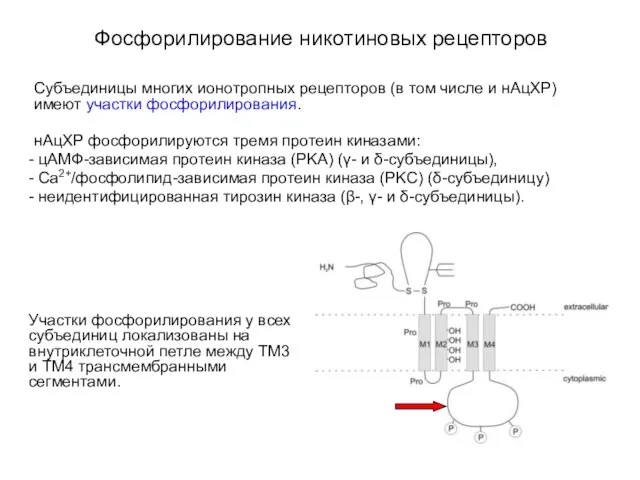
Фосфорилирование никотиновых рецепторов
Субъединицы многих ионотропных рецепторов (в том числе и нАцХР)
Фосфорилирование никотиновых рецепторов
Субъединицы многих ионотропных рецепторов (в том числе и нАцХР)

Десенситизация никотиновых рецепторов
Функциональное значение фосфорилирования рецепторов состоит в увеличении степени десенситизации.
Десенситизация
Десенситизация никотиновых рецепторов
Функциональное значение фосфорилирования рецепторов состоит в увеличении степени десенситизации.
Десенситизация

Постсинаптическая модуляция: ауторегуляция
Отрицательная ауторегуляция
Десенситизация (десенсибилизация) рецепторов при длительном воздействии медиатора.
Пример:
Если постсинаптическая
Постсинаптическая модуляция: ауторегуляция
Отрицательная ауторегуляция
Десенситизация (десенсибилизация) рецепторов при длительном воздействии медиатора.
Пример:
Если постсинаптическая

Субъединицы никотиновых рецепторов
Методами гибридизации ДНК и генной инженерии всего идентифицировано
10
Субъединицы никотиновых рецепторов
Методами гибридизации ДНК и генной инженерии всего идентифицировано
10
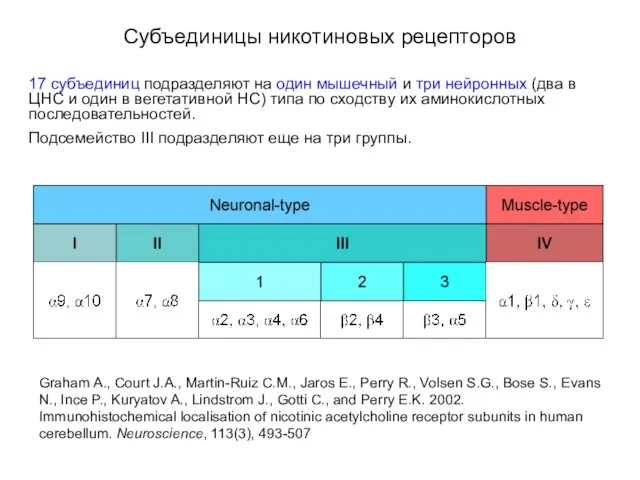
Субъединицы никотиновых рецепторов
17 субъединиц подразделяют на один мышечный и три нейронных
Субъединицы никотиновых рецепторов
17 субъединиц подразделяют на один мышечный и три нейронных
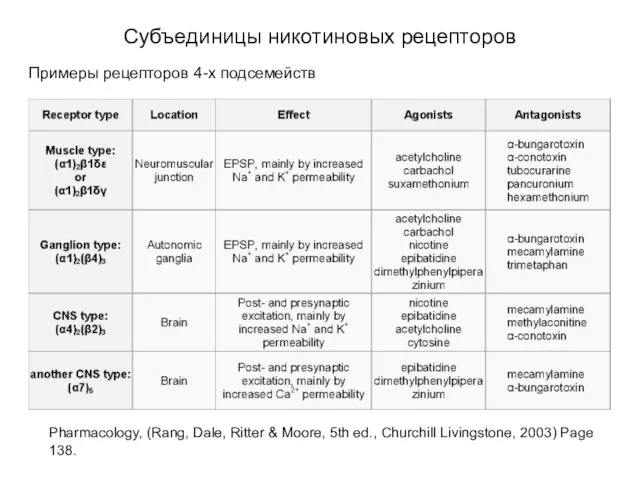
Субъединицы никотиновых рецепторов
Примеры рецепторов 4-х подсемейств
Pharmacology, (Rang, Dale, Ritter & Moore,
Субъединицы никотиновых рецепторов
Примеры рецепторов 4-х подсемейств
Pharmacology, (Rang, Dale, Ritter & Moore,
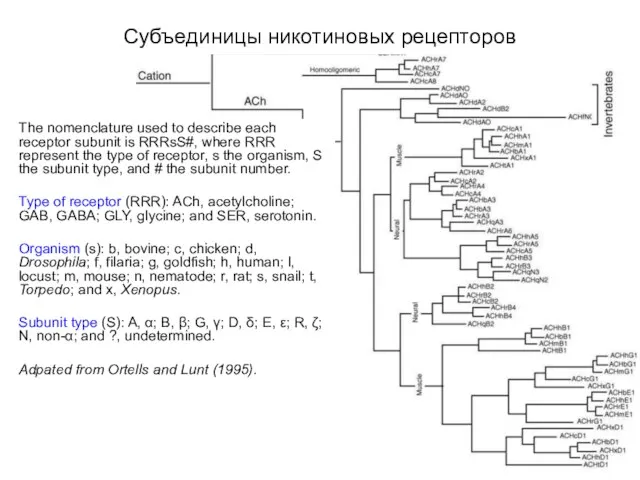
Субъединицы никотиновых рецепторов
The nomenclature used to describe each receptor subunit is
Субъединицы никотиновых рецепторов
The nomenclature used to describe each receptor subunit is
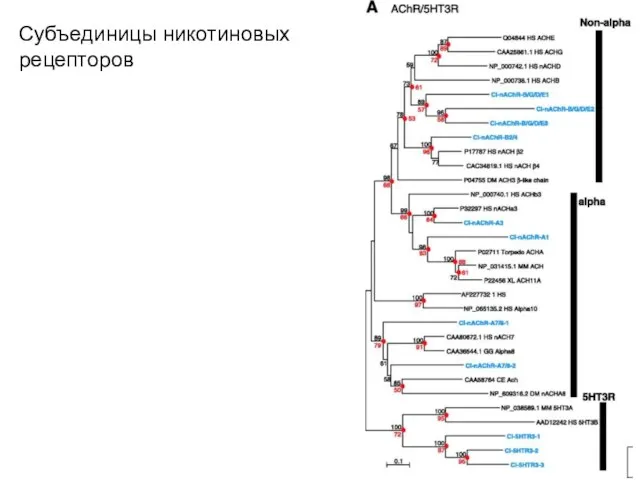
Субъединицы никотиновых рецепторов
Субъединицы никотиновых рецепторов
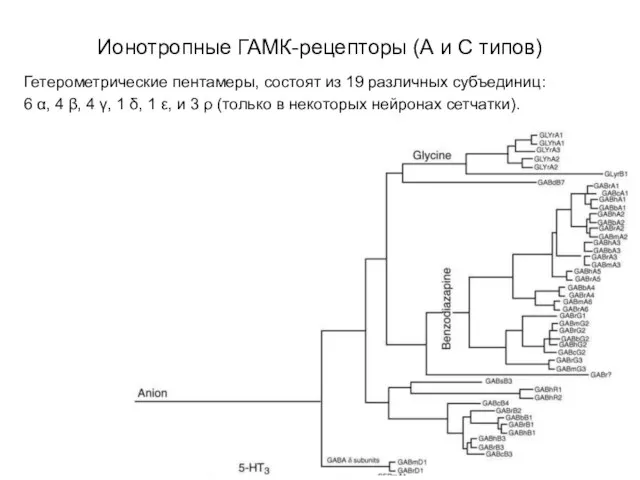
Ионотропные ГАМК-рецепторы (А и С типов)
Гетерометрические пентамеры, состоят из 19 различных
Ионотропные ГАМК-рецепторы (А и С типов)
Гетерометрические пентамеры, состоят из 19 различных
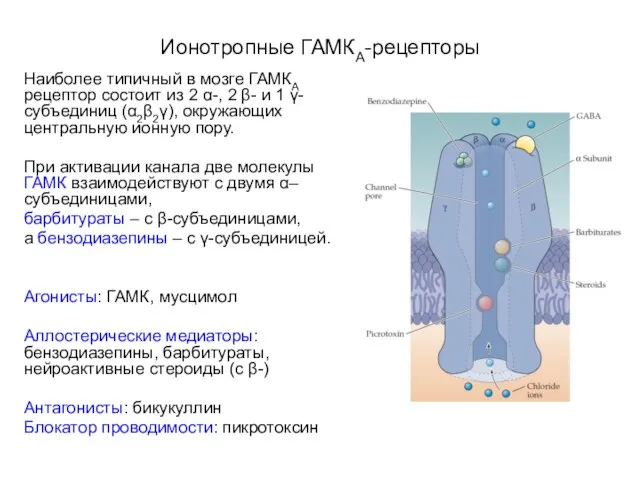
Ионотропные ГАМКА-рецепторы
Наиболее типичный в мозге ГАМКА рецептор состоит из 2 α-,
Ионотропные ГАМКА-рецепторы
Наиболее типичный в мозге ГАМКА рецептор состоит из 2 α-,
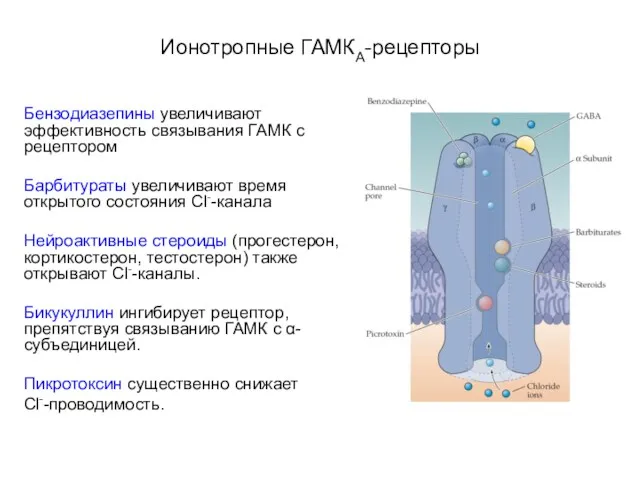
Ионотропные ГАМКА-рецепторы
Бензодиазепины увеличивают эффективность связывания ГАМК с рецептором
Барбитураты увеличивают время открытого
Ионотропные ГАМКА-рецепторы
Бензодиазепины увеличивают эффективность связывания ГАМК с рецептором
Барбитураты увеличивают время открытого

Ионотропные ГАМКС-рецепторы
ГАМКС рецепторы нечувствительны к агонистам ГАМКА рецепторов – к бензодиазепинам,
Ионотропные ГАМКС-рецепторы
ГАМКС рецепторы нечувствительны к агонистам ГАМКА рецепторов – к бензодиазепинам,
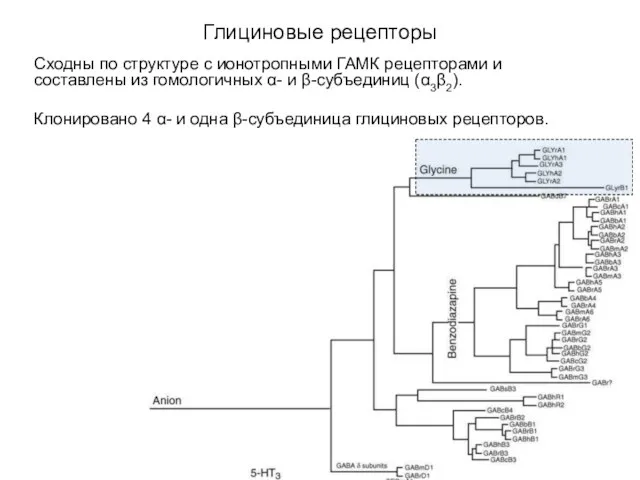
Глициновые рецепторы
Сходны по структуре с ионотропными ГАМК рецепторами и составлены
Глициновые рецепторы
Сходны по структуре с ионотропными ГАМК рецепторами и составлены
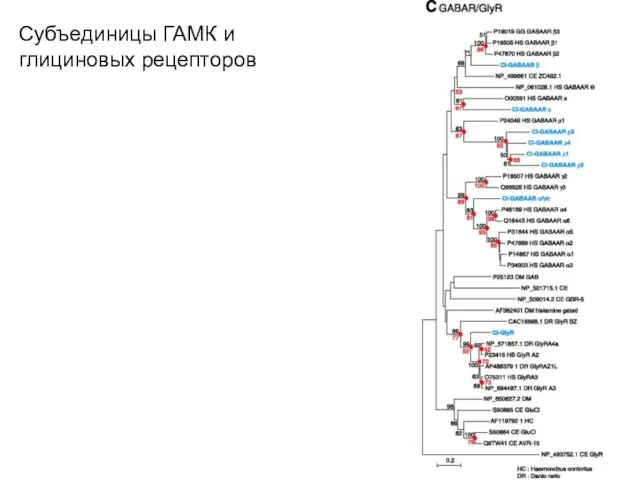
Субъединицы ГАМК и глициновых рецепторов
Субъединицы ГАМК и глициновых рецепторов

Глициновые рецепторы
При биохимической очистке из ткани вместе с белком глицинового
Глициновые рецепторы
При биохимической очистке из ткани вместе с белком глицинового
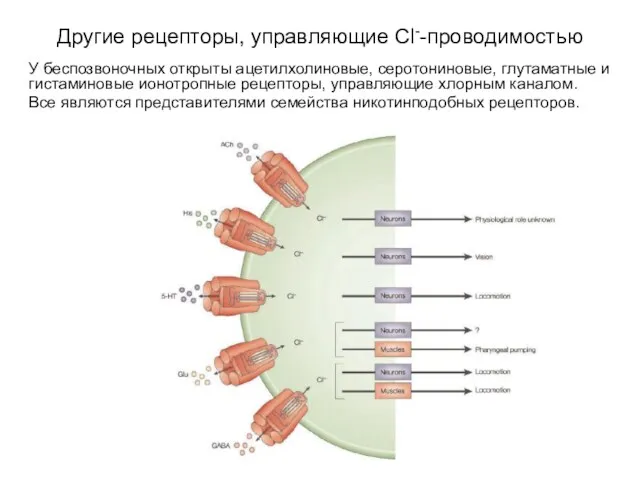
Другие рецепторы, управляющие Cl--проводимостью
У беспозвоночных открыты ацетилхолиновые, серотониновые, глутаматные и
Другие рецепторы, управляющие Cl--проводимостью
У беспозвоночных открыты ацетилхолиновые, серотониновые, глутаматные и
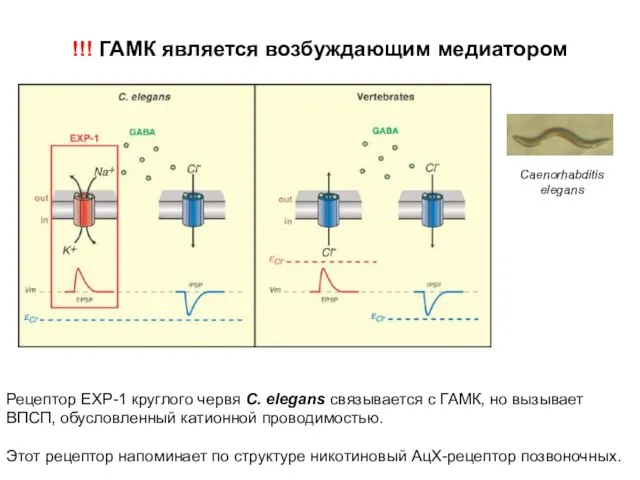
!!! ГАМК является возбуждающим медиатором
Рецептор EXP-1 круглого червя С. elegans связывается
!!! ГАМК является возбуждающим медиатором
Рецептор EXP-1 круглого червя С. elegans связывается
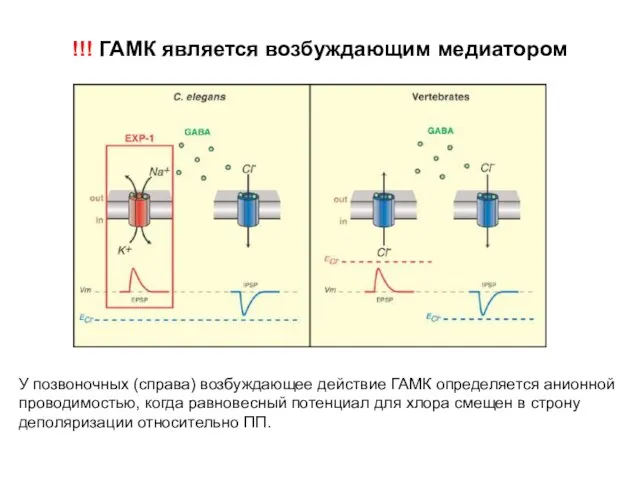
!!! ГАМК является возбуждающим медиатором
У позвоночных (справа) возбуждающее действие ГАМК определяется
!!! ГАМК является возбуждающим медиатором
У позвоночных (справа) возбуждающее действие ГАМК определяется
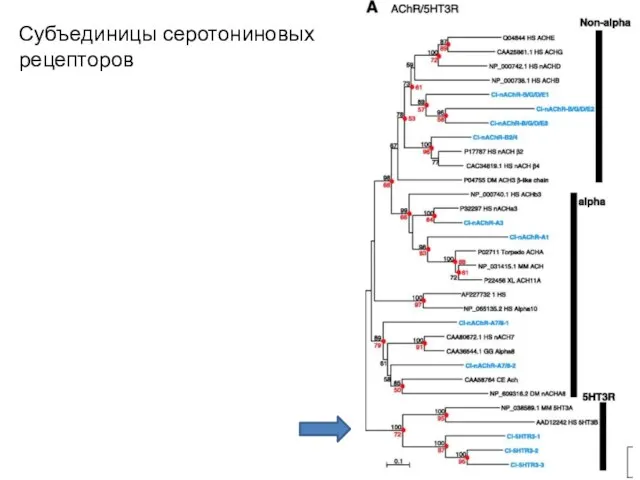
Субъединицы серотониновых рецепторов
Субъединицы серотониновых рецепторов

Серотониновые ионотропные рецепторы (5-НТ3)
Структура близка к структуре нАцХР.
Идентифицировано две субъединицы
Серотониновые ионотропные рецепторы (5-НТ3)
Структура близка к структуре нАцХР.
Идентифицировано две субъединицы

Серотониновые ионотропные рецепторы (5-НТ3)
При активации 5-НТ3-рецепторы обеспечивают проводимость моновалентных катионов
Серотониновые ионотропные рецепторы (5-НТ3)
При активации 5-НТ3-рецепторы обеспечивают проводимость моновалентных катионов
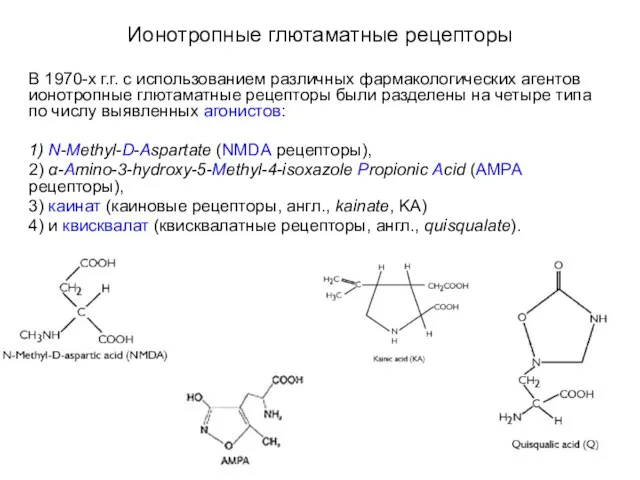
Ионотропные глютаматные рецепторы
В 1970-х г.г. с использованием различных фармакологических агентов
Ионотропные глютаматные рецепторы
В 1970-х г.г. с использованием различных фармакологических агентов

Ионотропные глютаматные рецепторы
В 1970-х г.г. с использованием различных фармакологических агентов
Ионотропные глютаматные рецепторы
В 1970-х г.г. с использованием различных фармакологических агентов
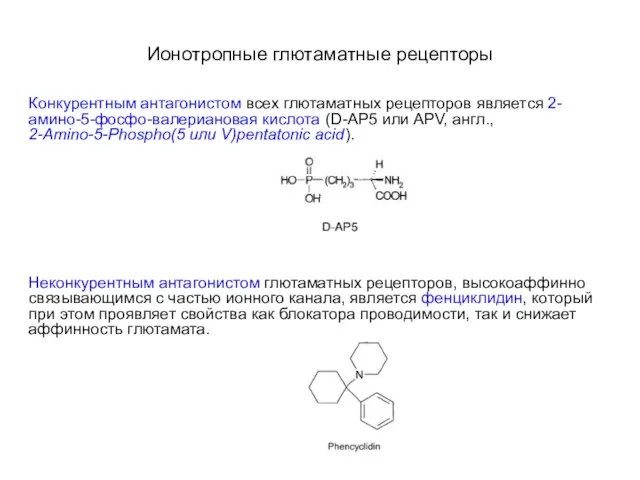
Ионотропные глютаматные рецепторы
Конкурентным антагонистом всех глютаматных рецепторов является 2-амино-5-фосфо-валериановая кислота
Ионотропные глютаматные рецепторы
Конкурентным антагонистом всех глютаматных рецепторов является 2-амино-5-фосфо-валериановая кислота
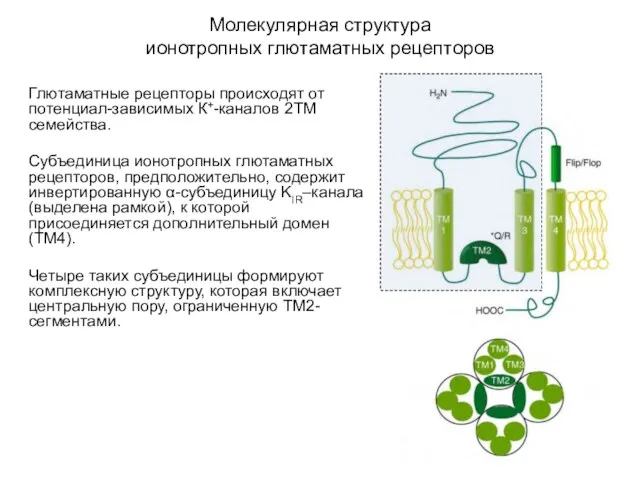
Молекулярная структура
ионотропных глютаматных рецепторов
Глютаматные рецепторы происходят от потенциал-зависимых К+-каналов 2ТМ
Молекулярная структура
ионотропных глютаматных рецепторов
Глютаматные рецепторы происходят от потенциал-зависимых К+-каналов 2ТМ
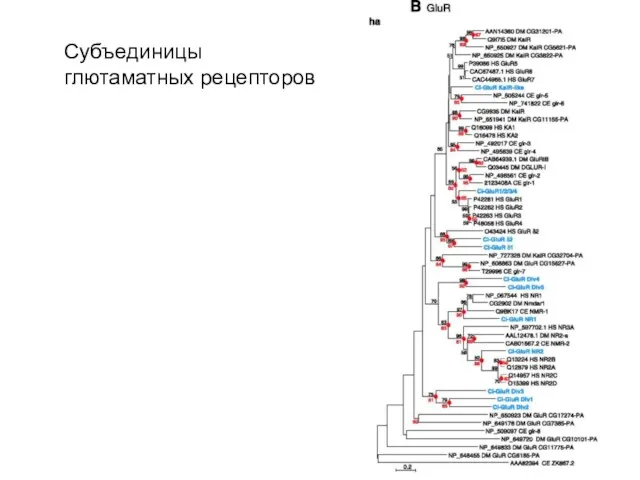
Субъединицы глютаматных рецепторов
Субъединицы глютаматных рецепторов
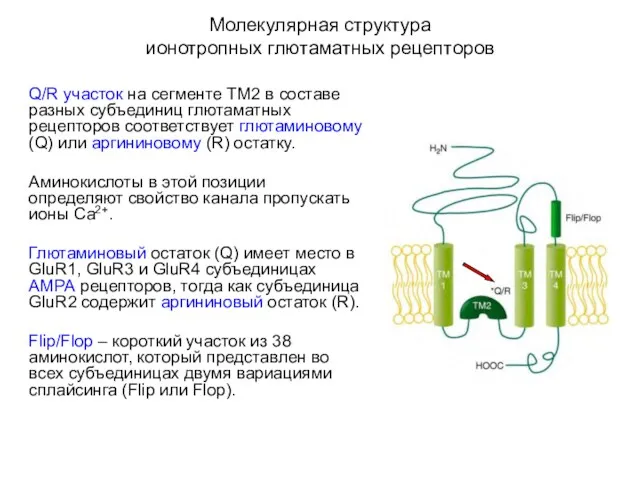
Молекулярная структура
ионотропных глютаматных рецепторов
Q/R участок на сегменте ТМ2 в составе
Молекулярная структура
ионотропных глютаматных рецепторов
Q/R участок на сегменте ТМ2 в составе
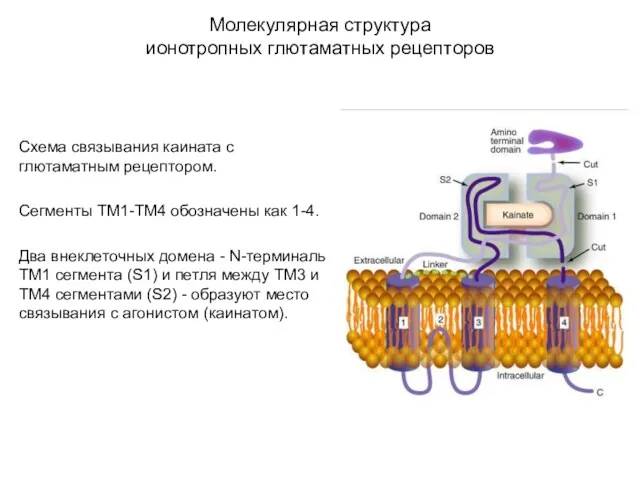
Молекулярная структура
ионотропных глютаматных рецепторов
Схема связывания каината с глютаматным рецептором.
Сегменты ТМ1-ТМ4
Молекулярная структура
ионотропных глютаматных рецепторов
Схема связывания каината с глютаматным рецептором.
Сегменты ТМ1-ТМ4
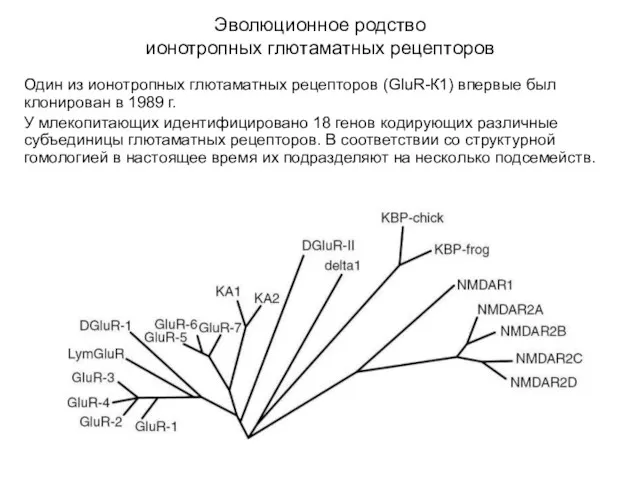
Эволюционное родство
ионотропных глютаматных рецепторов
Один из ионотропных глютаматных рецепторов (GluR-К1) впервые был
Эволюционное родство
ионотропных глютаматных рецепторов
Один из ионотропных глютаматных рецепторов (GluR-К1) впервые был
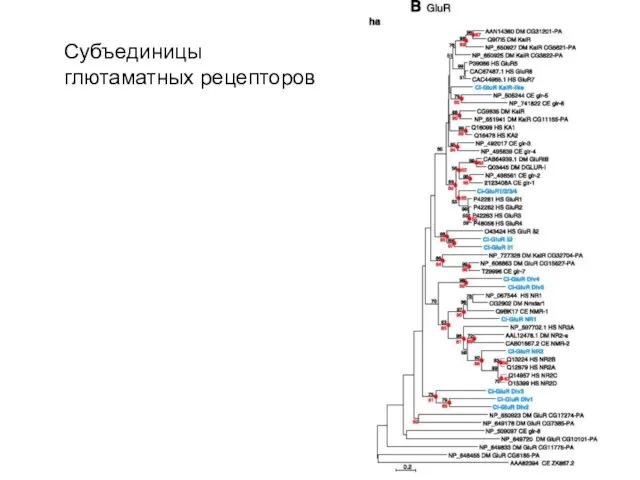
Субъединицы глютаматных рецепторов
Субъединицы глютаматных рецепторов
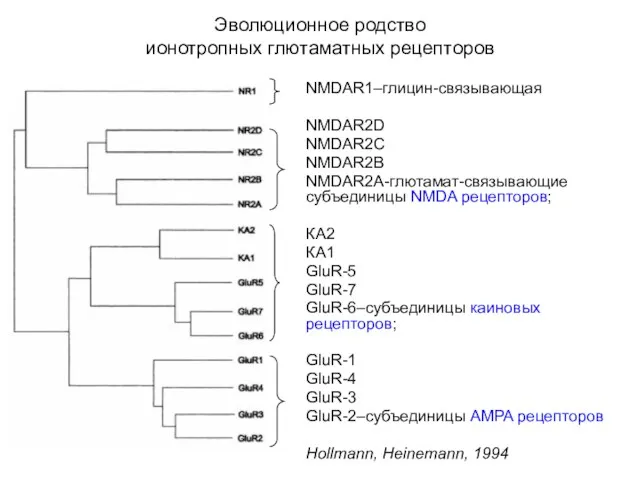
Эволюционное родство
ионотропных глютаматных рецепторов
NMDAR1–глицин-связывающая
NMDAR2D
NMDAR2C
NMDAR2B
NMDAR2A-глютамат-связывающие субъединицы NMDA рецепторов;
КА2
КА1
GluR-5
GluR-7
GluR-6–субъединицы каиновых рецепторов;
GluR-1
GluR-4
GluR-3
GluR-2–субъединицы AMPA рецепторов
Hollmann,
Эволюционное родство
ионотропных глютаматных рецепторов
NMDAR1–глицин-связывающая
NMDAR2D
NMDAR2C
NMDAR2B
NMDAR2A-глютамат-связывающие субъединицы NMDA рецепторов;
КА2
КА1
GluR-5
GluR-7
GluR-6–субъединицы каиновых рецепторов;
GluR-1
GluR-4
GluR-3
GluR-2–субъединицы AMPA рецепторов
Hollmann,
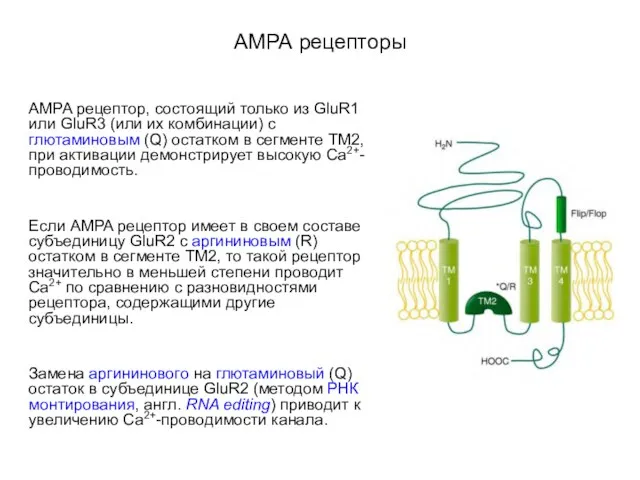
АМРА рецепторы
AMPA рецептор, состоящий только из GluR1 или GluR3 (или их
АМРА рецепторы
AMPA рецептор, состоящий только из GluR1 или GluR3 (или их

АМРА рецепторы
Flip/Flop – участок из 38 аминокислот, который представлен во всех
АМРА рецепторы
Flip/Flop – участок из 38 аминокислот, который представлен во всех
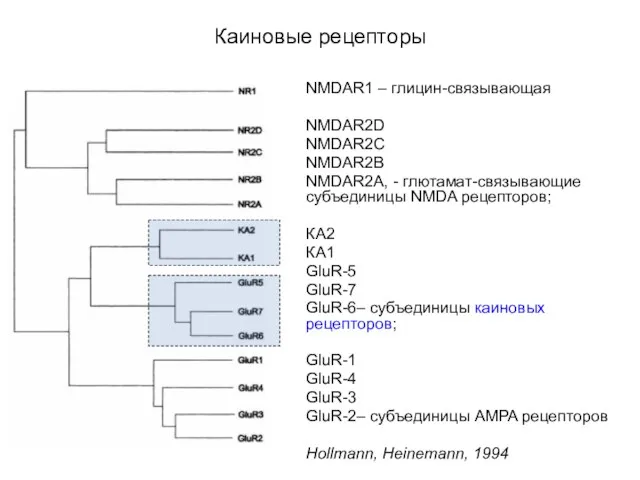
Каиновые рецепторы
NMDAR1 – глицин-связывающая
NMDAR2D
NMDAR2C
NMDAR2B
NMDAR2A, - глютамат-связывающие субъединицы NMDA рецепторов;
КА2
КА1
GluR-5
GluR-7
GluR-6– субъединицы каиновых
Каиновые рецепторы
NMDAR1 – глицин-связывающая
NMDAR2D
NMDAR2C
NMDAR2B
NMDAR2A, - глютамат-связывающие субъединицы NMDA рецепторов;
КА2
КА1
GluR-5
GluR-7
GluR-6– субъединицы каиновых
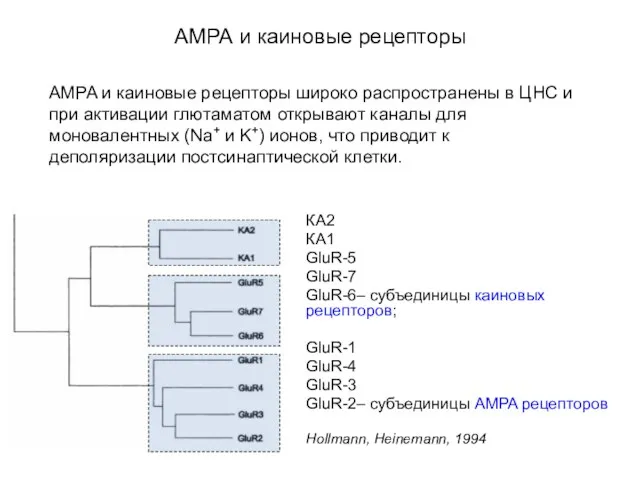
АМРА и каиновые рецепторы
КА2
КА1
GluR-5
GluR-7
GluR-6– субъединицы каиновых рецепторов;
GluR-1
GluR-4
GluR-3
GluR-2– субъединицы AMPA рецепторов
Hollmann, Heinemann,
АМРА и каиновые рецепторы
КА2
КА1
GluR-5
GluR-7
GluR-6– субъединицы каиновых рецепторов;
GluR-1
GluR-4
GluR-3
GluR-2– субъединицы AMPA рецепторов
Hollmann, Heinemann,
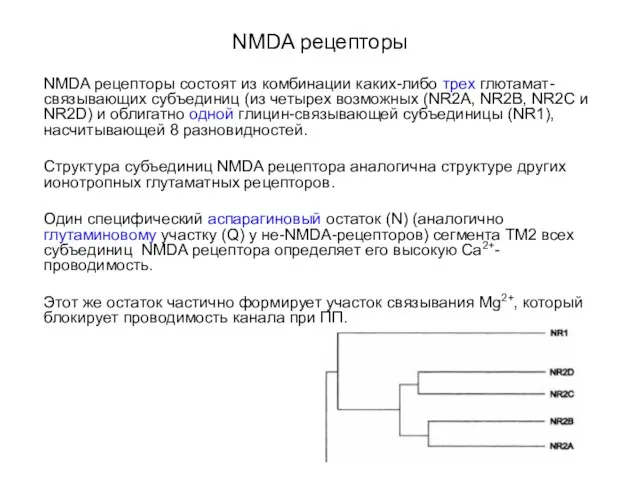
NMDA рецепторы
NMDA рецепторы состоят из комбинации каких-либо трех глютамат-связывающих субъединиц (из
NMDA рецепторы
NMDA рецепторы состоят из комбинации каких-либо трех глютамат-связывающих субъединиц (из
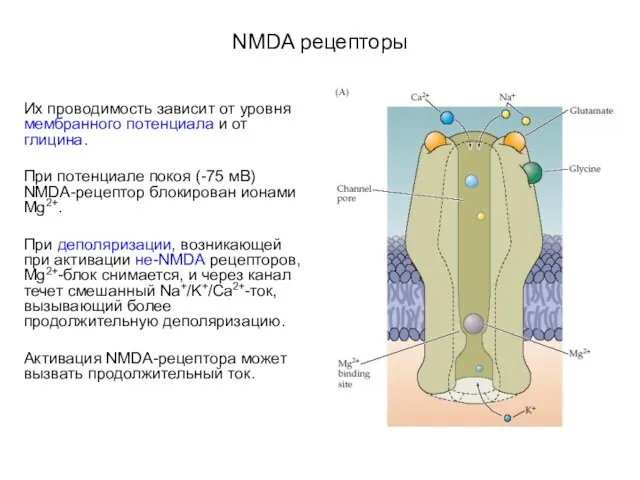
NMDA рецепторы
Их проводимость зависит от уровня мембранного потенциала и от глицина.
При
NMDA рецепторы
Их проводимость зависит от уровня мембранного потенциала и от глицина.
При
NMDA рецепторы
Их проводимость зависит от уровня мембранного потенциала и присутствия глицина.
При
NMDA рецепторы
Их проводимость зависит от уровня мембранного потенциала и присутствия глицина.
При
NMDA рецепторы
NMDA-рецептор имеет несколько участков, взаимодействующих аллостерически:
1) участок связывания медиатора;
2) регуляторный,
NMDA рецепторы
NMDA-рецептор имеет несколько участков, взаимодействующих аллостерически:
1) участок связывания медиатора;
2) регуляторный,
NMDA рецепторы
Сам глицин не вызывает ответа, но усиливает активацию NMDA-рецептора, увеличивая
NMDA рецепторы
Сам глицин не вызывает ответа, но усиливает активацию NMDA-рецептора, увеличивая
Пуриновые ионотропные рецепторы
Связываются с внеклеточной АТФ или ее производными (например,
Пуриновые ионотропные рецепторы
Связываются с внеклеточной АТФ или ее производными (например,

Пуриновые ионотропные рецепторы
Связываются с внеклеточной АТФ или ее производными (например,
Пуриновые ионотропные рецепторы
Связываются с внеклеточной АТФ или ее производными (например,
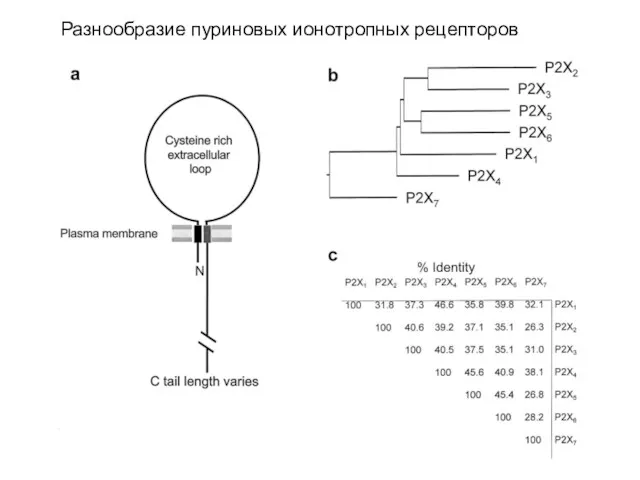
Разнообразие пуриновых ионотропных рецепторов
Разнообразие пуриновых ионотропных рецепторов
Пуриновые ионотропные рецепторы
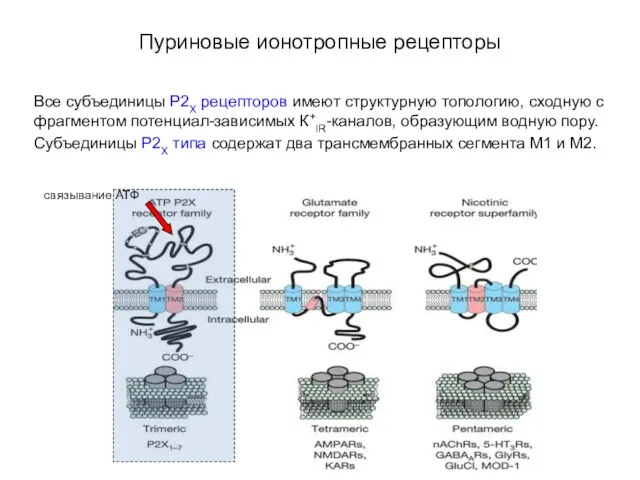
Все субъединицы P2X рецепторов имеют структурную топологию, сходную
Пуриновые ионотропные рецепторы
Все субъединицы P2X рецепторов имеют структурную топологию, сходную
 Пищеварение в кишечнике
Пищеварение в кишечнике презентация Значение ОДС. Строение костей
презентация Значение ОДС. Строение костей Пищевые добавки. Классификация пищевых добавок
Пищевые добавки. Классификация пищевых добавок Ткани человека. Виды и строение
Ткани человека. Виды и строение Сторінка дослідника. Твій город на підвіконні. Урок №76. Я досліджую світ
Сторінка дослідника. Твій город на підвіконні. Урок №76. Я досліджую світ Простейшие. Одноклеточные животные и колониальные организмы
Простейшие. Одноклеточные животные и колониальные организмы Пищеварение в кишечнике. Регуляция пищеварения
Пищеварение в кишечнике. Регуляция пищеварения Эволюция органов и функций
Эволюция органов и функций Прокариоты, бактерии, цианобактерии
Прокариоты, бактерии, цианобактерии Characteristic of families Lamiaceae and Asteraceae, typical species and practical uses
Characteristic of families Lamiaceae and Asteraceae, typical species and practical uses Условия полноценного развития системы кровообращения
Условия полноценного развития системы кровообращения Виды чеснока. Способы выращивания
Виды чеснока. Способы выращивания Эволюция органов размножения у животных
Эволюция органов размножения у животных Дистанционный курс по биологии. Тема Обмен веществ
Дистанционный курс по биологии. Тема Обмен веществ Строение растительной и животной клеток
Строение растительной и животной клеток Презентация к уроку биологии 8 класс на тему: Общая характеристика пресмыкающихся
Презентация к уроку биологии 8 класс на тему: Общая характеристика пресмыкающихся Алдыңғы іш қабырғасының анатомиялық құрылымы
Алдыңғы іш қабырғасының анатомиялық құрылымы 20231101_vestniki_vesny_viktorina
20231101_vestniki_vesny_viktorina Чарльз Роберт Дарвин: фрагменты жизни
Чарльз Роберт Дарвин: фрагменты жизни Систематика растений и животных в Новое время
Систематика растений и животных в Новое время Ластоногі. Група водних хижих ссавців
Ластоногі. Група водних хижих ссавців СРС на тему: “Везикулярный транспорт”
СРС на тему: “Везикулярный транспорт” Ақтөбе облыстық білім алуға ерекше қажеттіліктері бар балаларға арналған арнайы (түзетім) балабақшасы
Ақтөбе облыстық білім алуға ерекше қажеттіліктері бар балаларға арналған арнайы (түзетім) балабақшасы Дидактическая игра Самые умные
Дидактическая игра Самые умные מבנה תא חיידק
מבנה תא חיידק Анатомо - функциональные особенности скелета туловища
Анатомо - функциональные особенности скелета туловища Екологічна мережа
Екологічна мережа Земляника садовая
Земляника садовая