- Основные особенности растительной клетки

Содержание
- 2. Органеллы эукариотической клетки Ядро содержит основную часть генома и является местом синтеза ДНК и РНК Эндоплазматичесий
- 3. Классификация органелл Ядро и цитозоль связаны между собой ядерными порами, являются топологически едиными, но выполняют разные
- 4. Два пути сортировки белков: цитоплазматический и секреторный используется для секретируемых белков, белков плазмалеммы и белков органелл
- 5. Сигнальные последовательности транспорта белков в растительной клетке
- 6. Транспорт ядерно-кодируемых белков в хлоропласт
- 7. Секреторный путь транспорта белков: общая схема
- 8. Секреторный путь транспорта белков: транспорт в ЭР
- 9. Секреторный путь транспорта белков: гликозилирование в ЭР
- 10. Клеточная стенка – это не «деревянная тюрьма» для несчастной клетки… С помощью клеточной стеки клетка решает
- 11. Полисахариды клеточной стенки построены всего из 11 сахаров
- 12. Строение микрофибрилл целлюлозы «Ядро» - ~50 цепочек целлюлозы, кристаллическая область, 3 х 5нм. Вокруг «ядра» -
- 13. Строение целлюлозо-синтазы
- 14. Электронные фотографии КС с целлюлозо-синтазой
- 15. Сшивочные гликаны (cross-linking glycans) Ксилоглюканы (XyGs) Гликаны со смешанной связью (злаки) Глюкуроно-арабиноксиланы (GAXs) Фуко-XyGs XXXG :
- 16. Гемицеллюлозы: ксилоглюкан двудольных (фуко-галакто-XyGs) (XXXG : XXFG ~ 50 : 50)
- 17. Гемицеллюлозы: ксилоглюкан двудольных (арабино-XyGs пасленовых и мяты)
- 18. Гемицеллюлозы: глюкуроно-арабиноксиланы двудольных и каммелиноидов
- 19. Гемицеллюлозы: глюкан злаковых
- 20. Состав гемицеллюлоз у представителей разных таксонов
- 21. Пектины Галактуронаны Рамногалактуронаны Гомогалактуронаны Ксилогалактуронаны Рамногалактуронаны II Рамногалактуронаны I
- 22. Пектины: галактоктуронаны (гомо- и ксило-галактуронаны)
- 23. Пектины: рамногалактуронаны I гетерополимер: линейная цепь из чередующихся остатков GalA и Rha с различными боковыми фрагментами)
- 24. Пектины: димер рамногалактуронана II мономеры RGII 4200kDa связаны диэфирными связями остатками апиозы через бор)
- 25. «Замковые зоны» пектиновой сети Синтез пектинов – В АГ в метоксилированном виде. Пектин-метил-эстераза (PME) избирательно отщепляет
- 26. Пектины: зоны «Ca2+-застежек» и количество нейтральных боковых цепочек RGI регулируют размер пор клеточной стенки
- 27. Пектины: функциональная сеть клеточной стенки Функции пектинов: определяют размер пор КС определяют поверхностный заряд КС адгезионные
- 28. Структурные белки клеточной стенки: HGRPs, PRPs, GRPs (гидроксипролин-, пролин- и глицин- обогащенные)
- 29. Структурные белки клеточной стенки: AGPs (арабино-галактановые белки - протеогликаны).
- 30. Структурные белки клеточной стенки: AGPs (арабино-галактановые белки - протеогликаны).
- 31. Трехмерная модель двух типов клеточной стенки: тип I (двудольные) и тип II (коммелиноиды)
- 32. Возможное участие ХЕТ (ксилоглюкан-эндотрансгликозилазы) и экспансина в росте клеток растяжением
- 33. Лигнины: фенилпропаниодная сеть вторичных клеточных стенок
- 34. Образование лигнина: окислительная конденсация фенилпропаноидов случайным образом.
- 35. Образование лигнинов: целенаправленная конденсация мономеров.
- 36. Некоторые особенности плазмалеммы Структурные: зависимость состава от типа клетки основные ЖК: пальмитиновая (16:0), олеиновая (18:1, ∆9),
- 37. Фитостерины, диацилглицериды и варианты «заякоривания» белков в мембранах Фосфолипиды (плазмалемма) Гликозилглицериды (пластиды)
- 42. Структура растительного аппарата Гольджи
- 43. Везикулярный транспорт, типы везикул COPII – транспорт от ER к Гольджи, COPI – «ретроградный» транспорт -
- 44. Синтез ксилоглюканов (А) и пектинов (В) проходит в разных компартментах АГ До сих пор неясно как
- 48. Некоторые особенности ядерного генома растений Размер: от ~108 тпн (Arabidopsis) до 1010 (бобы) – 1011 (Fritillaria)
- 49. Взаимопревращения пластид контролируются ядерным геномом
- 50. Хлоропласт – «главный» представитель пластид
- 51. Фитоферритин в пропластидах мезофилла сои, амилопласт
- 52. Этиопласт: структура проламеллярного тела, формирование хлоропласта
- 53. Структура хлоропластного генома риса. Два типа генома: c двумя IR размером (обычно около 20 kb). Почти
- 54. Структура хлоропластного генома разных видов растений
- 55. Сходства и отличия хлоропластного генома и белоксинтезирующей системы от бактериальных Сходства: Кольцевая ДНК Содержание G/С аналогично
- 56. Гены хлоропластов Транскрипция. 4 гена субъединиц пластидной РНК-полимеразы (rpo) Синтез белка. - 4 гена рРНК (оперон
- 57. Эукариотическая, бактериальная и пластидные РНК-полимеразы, множественность промоторов хлоропластных генов 1. Гены со стандартными эубактериальными промоторами (почти
- 58. PEP – кодируемая в пластидном геноме и NEP – кодируемая в ядре РНК-полимеразы, PEP: α и
- 59. Процессинг хлоропластной пре-рРНК растений Кстати, такой же порядок генов (rrn16–trnI–trnA–rrn23) характерен и для цианобактерий
- 60. Структуры зрелой пластидной и ядерной иРНК. Полиаденилирование выполняет для них функции с точностью до обратного… Для
- 61. Функции пластид Фотосинтез – NB Синтез: все жирные кислоты, многие аминокислоты, синтез пуринов и пиримидинов, альтернативный
- 62. Растительные митохондрии имеют разнообразный размер и форму
- 63. Предполагаемая структура «мастер-хромосомы» митохондрий кукурузы Митохондриальный геном растений имеет самый большой размер среди всех эукариотических клеток,
- 69. Скачать презентацию

Органеллы эукариотической клетки
Ядро содержит основную часть генома и является местом
Органеллы эукариотической клетки
Ядро содержит основную часть генома и является местом

Классификация органелл
Ядро и цитозоль связаны между собой ядерными порами, являются топологически
Классификация органелл
Ядро и цитозоль связаны между собой ядерными порами, являются топологически
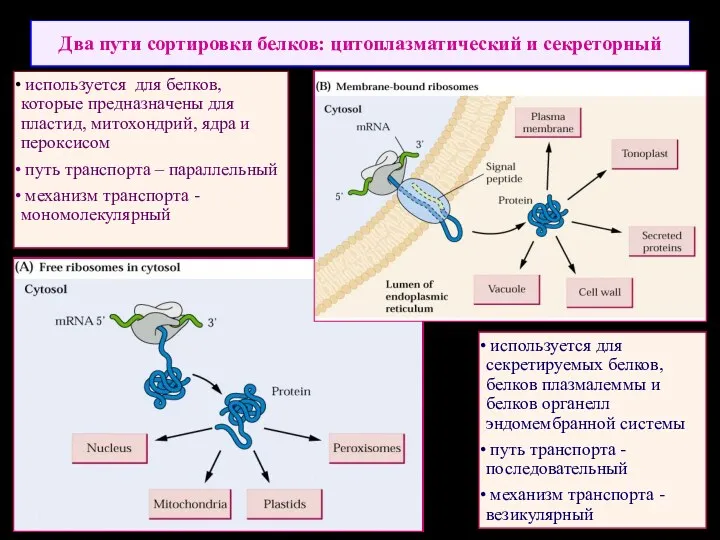
Два пути сортировки белков: цитоплазматический и секреторный
используется для секретируемых белков,
Два пути сортировки белков: цитоплазматический и секреторный
используется для секретируемых белков,
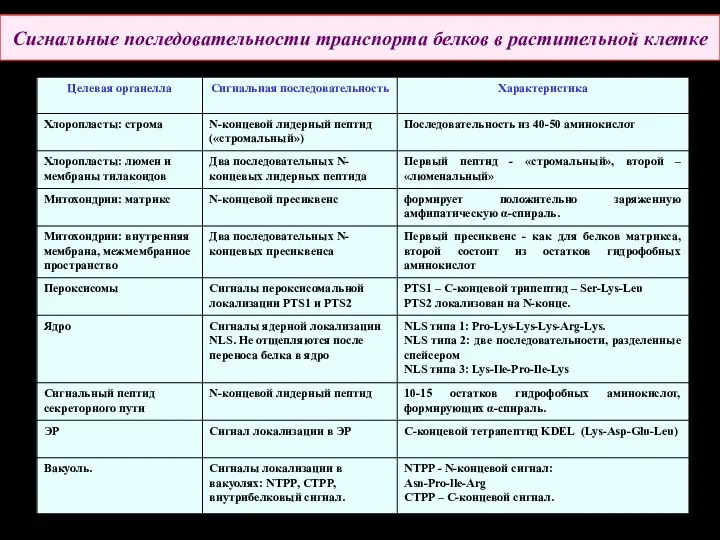
Сигнальные последовательности транспорта белков в растительной клетке
Сигнальные последовательности транспорта белков в растительной клетке

Транспорт ядерно-кодируемых белков в хлоропласт
Транспорт ядерно-кодируемых белков в хлоропласт

Секреторный путь транспорта белков: общая схема
Секреторный путь транспорта белков: общая схема

Секреторный путь транспорта белков: транспорт в ЭР
Секреторный путь транспорта белков: транспорт в ЭР

Секреторный путь транспорта белков: гликозилирование в ЭР
Секреторный путь транспорта белков: гликозилирование в ЭР

Клеточная стенка – это не «деревянная тюрьма» для несчастной клетки…
С
Клеточная стенка – это не «деревянная тюрьма» для несчастной клетки…
С
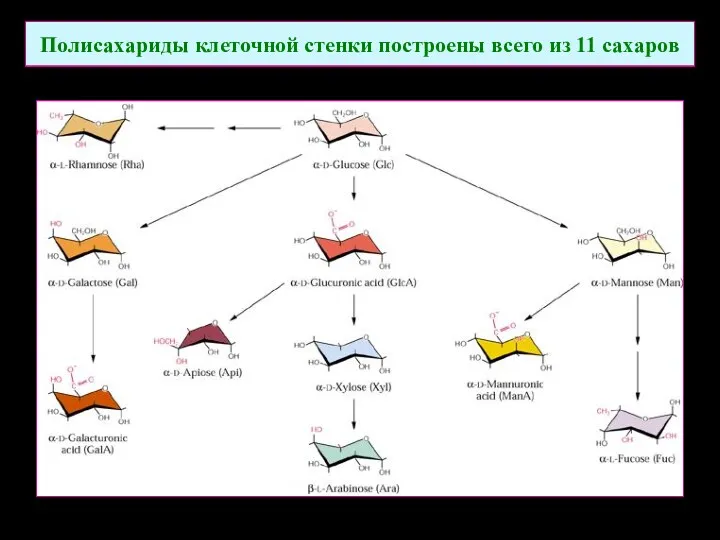
Полисахариды клеточной стенки построены всего из 11 сахаров
Полисахариды клеточной стенки построены всего из 11 сахаров
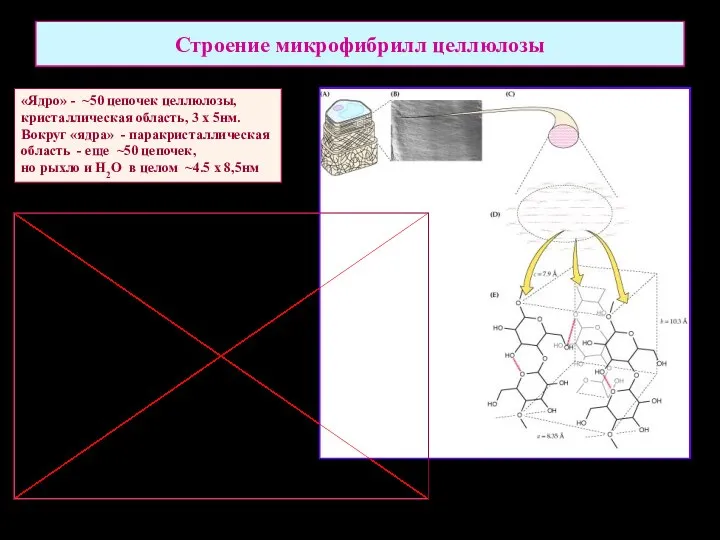
Строение микрофибрилл целлюлозы
«Ядро» - ~50 цепочек целлюлозы,
кристаллическая область, 3 х
Строение микрофибрилл целлюлозы
«Ядро» - ~50 цепочек целлюлозы,
кристаллическая область, 3 х
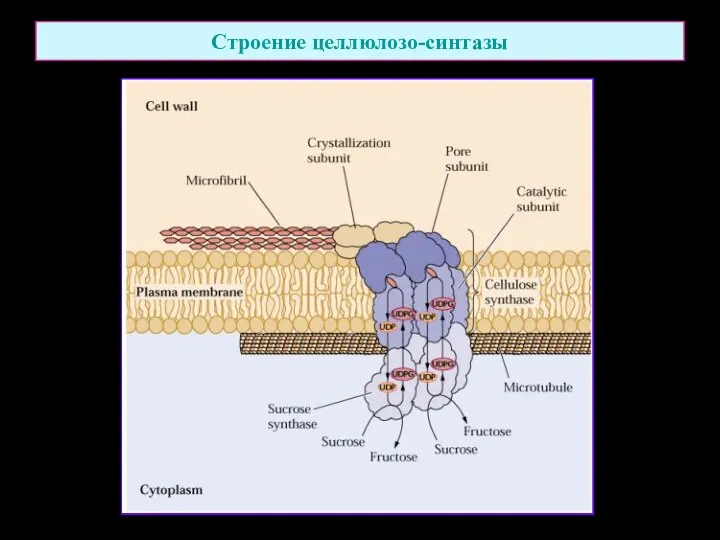
Строение целлюлозо-синтазы
Строение целлюлозо-синтазы

Электронные фотографии КС с целлюлозо-синтазой
Электронные фотографии КС с целлюлозо-синтазой
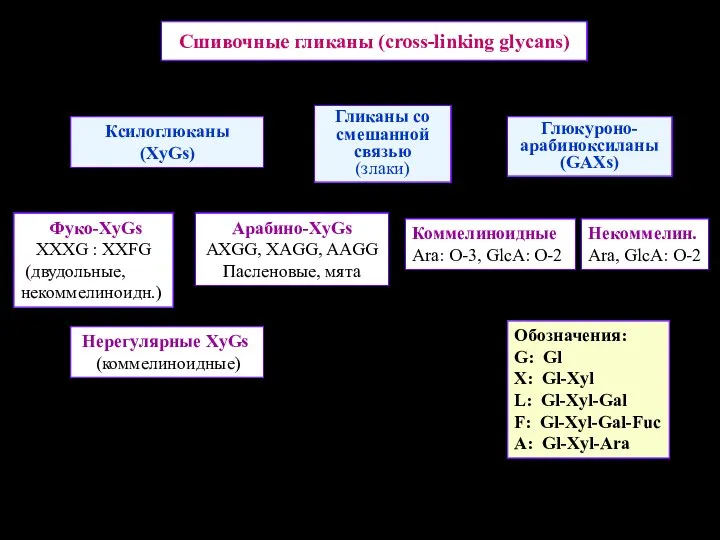
Сшивочные гликаны (cross-linking glycans)
Ксилоглюканы (XyGs)
Гликаны со смешанной связью
(злаки)
Глюкуроно-арабиноксиланы (GAXs)
Фуко-XyGs XXXG
Сшивочные гликаны (cross-linking glycans)
Ксилоглюканы (XyGs)
Гликаны со смешанной связью
(злаки)
Глюкуроно-арабиноксиланы (GAXs)
Фуко-XyGs XXXG
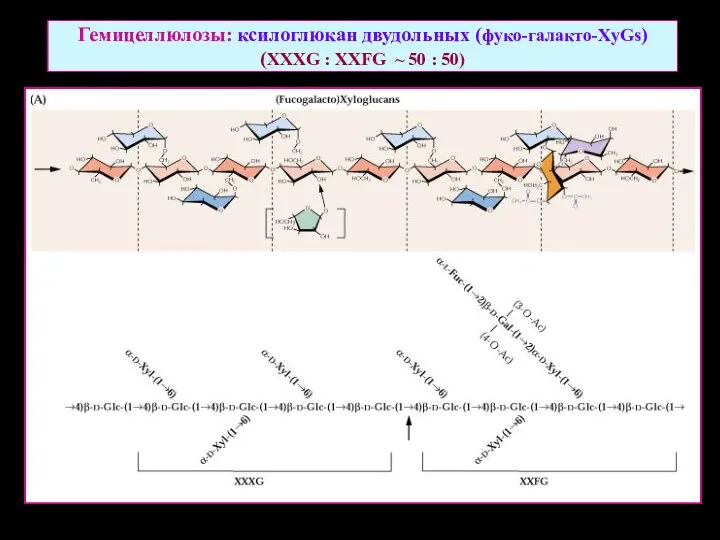
Гемицеллюлозы: ксилоглюкан двудольных (фуко-галакто-XyGs)
(XXXG : XXFG ~ 50 : 50)
Гемицеллюлозы: ксилоглюкан двудольных (фуко-галакто-XyGs)
(XXXG : XXFG ~ 50 : 50)
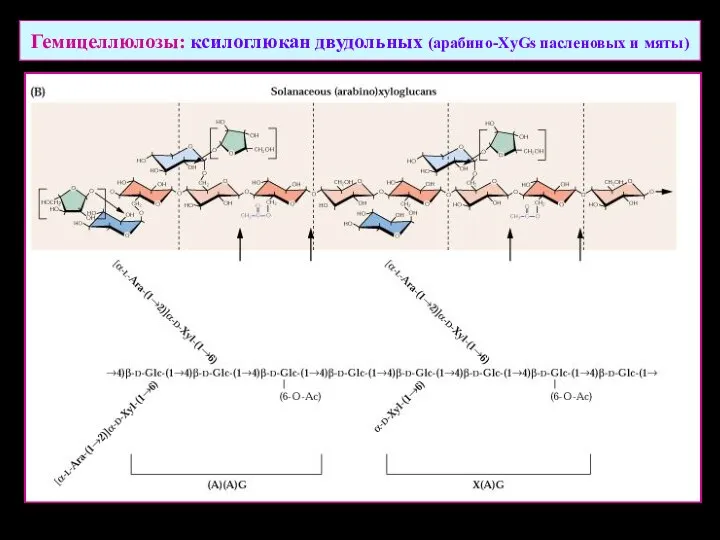
Гемицеллюлозы: ксилоглюкан двудольных (арабино-XyGs пасленовых и мяты)
Гемицеллюлозы: ксилоглюкан двудольных (арабино-XyGs пасленовых и мяты)
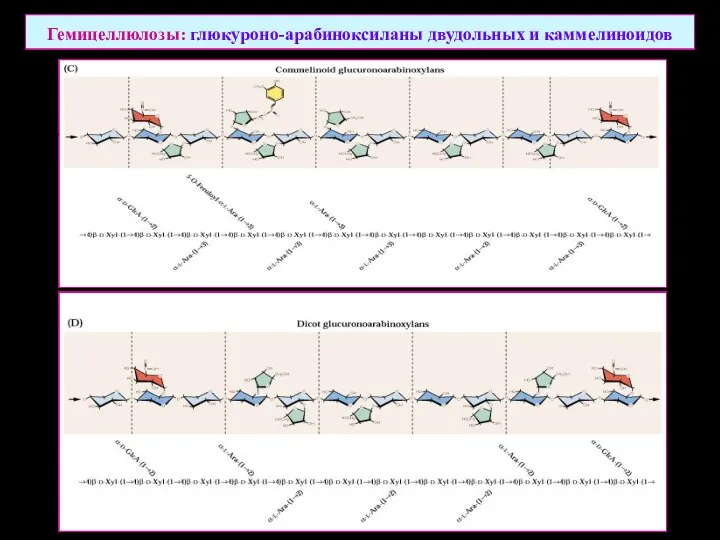
Гемицеллюлозы: глюкуроно-арабиноксиланы двудольных и каммелиноидов
Гемицеллюлозы: глюкуроно-арабиноксиланы двудольных и каммелиноидов
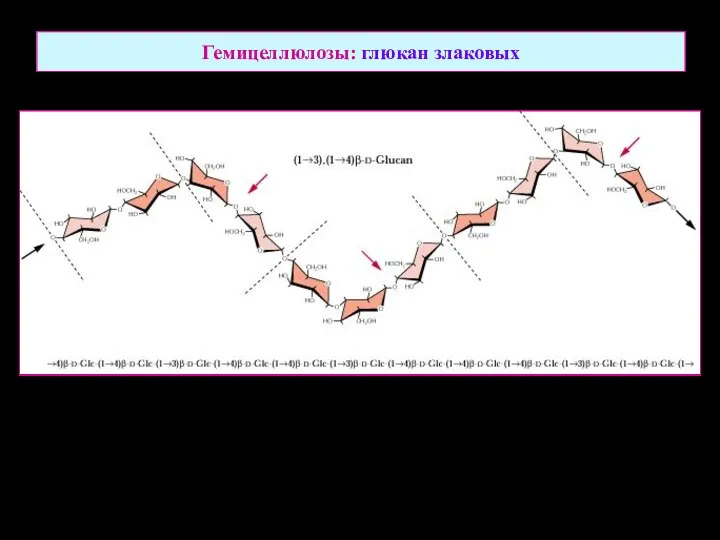
Гемицеллюлозы: глюкан злаковых
Гемицеллюлозы: глюкан злаковых

Состав гемицеллюлоз у представителей разных таксонов
Состав гемицеллюлоз у представителей разных таксонов
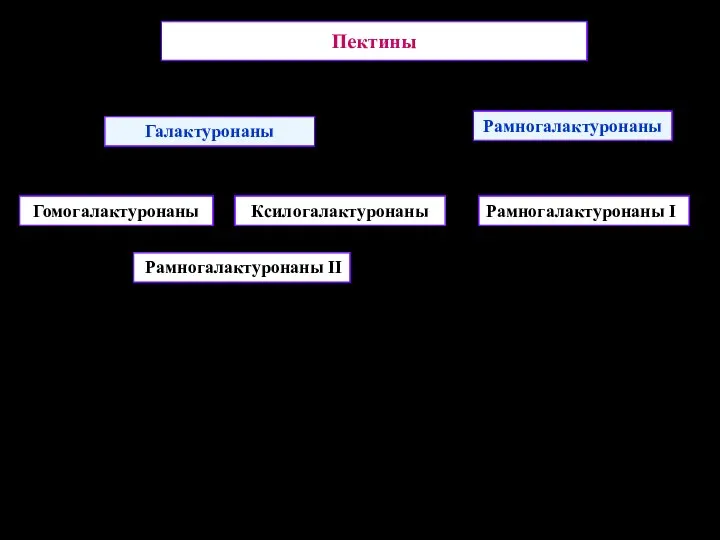
Пектины
Галактуронаны
Рамногалактуронаны
Гомогалактуронаны
Ксилогалактуронаны
Рамногалактуронаны II
Рамногалактуронаны I
Пектины
Галактуронаны
Рамногалактуронаны
Гомогалактуронаны
Ксилогалактуронаны
Рамногалактуронаны II
Рамногалактуронаны I
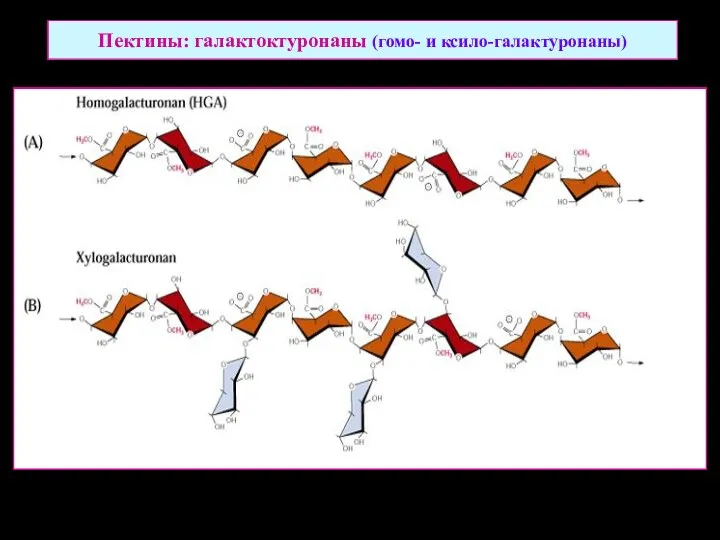
Пектины: галактоктуронаны (гомо- и ксило-галактуронаны)
Пектины: галактоктуронаны (гомо- и ксило-галактуронаны)
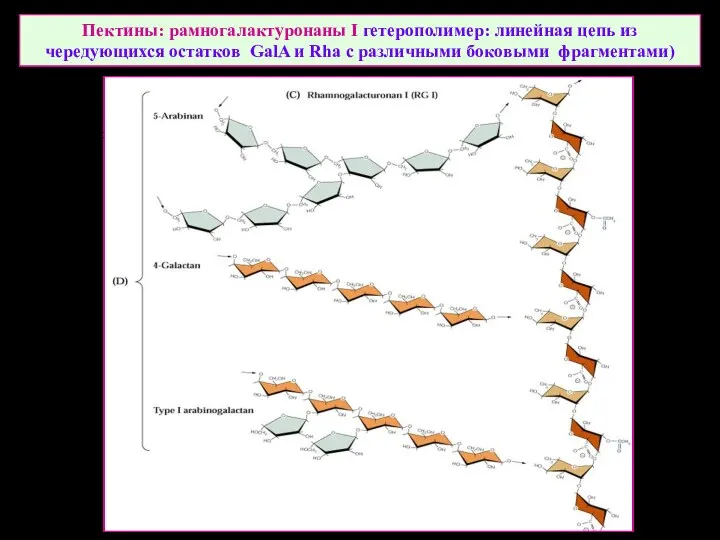
Пектины: рамногалактуронаны I гетерополимер: линейная цепь из чередующихся остатков GalA и
Пектины: рамногалактуронаны I гетерополимер: линейная цепь из чередующихся остатков GalA и
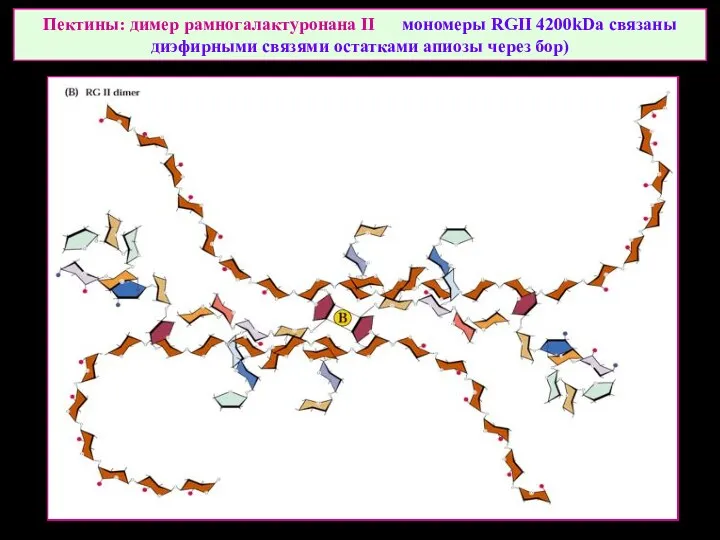
Пектины: димер рамногалактуронана II мономеры RGII 4200kDa связаны диэфирными связями остатками
Пектины: димер рамногалактуронана II мономеры RGII 4200kDa связаны диэфирными связями остатками
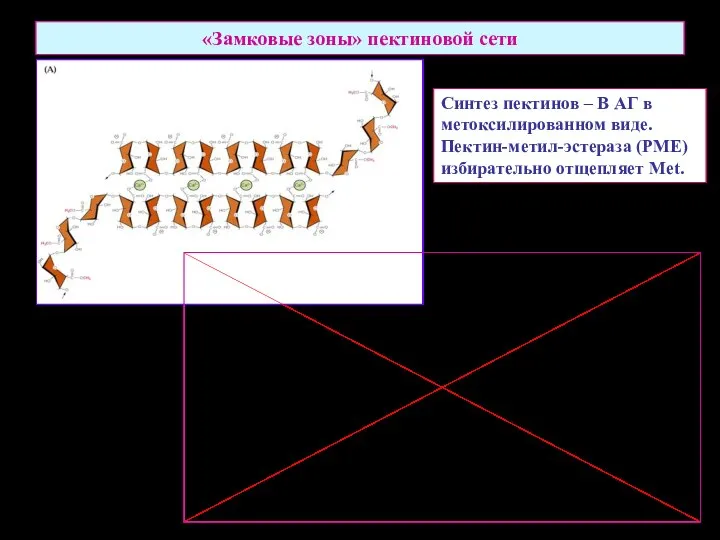
«Замковые зоны» пектиновой сети
Синтез пектинов – В АГ в
метоксилированном виде.
Пектин-метил-эстераза (PME)
избирательно
«Замковые зоны» пектиновой сети
Синтез пектинов – В АГ в
метоксилированном виде.
Пектин-метил-эстераза (PME)
избирательно
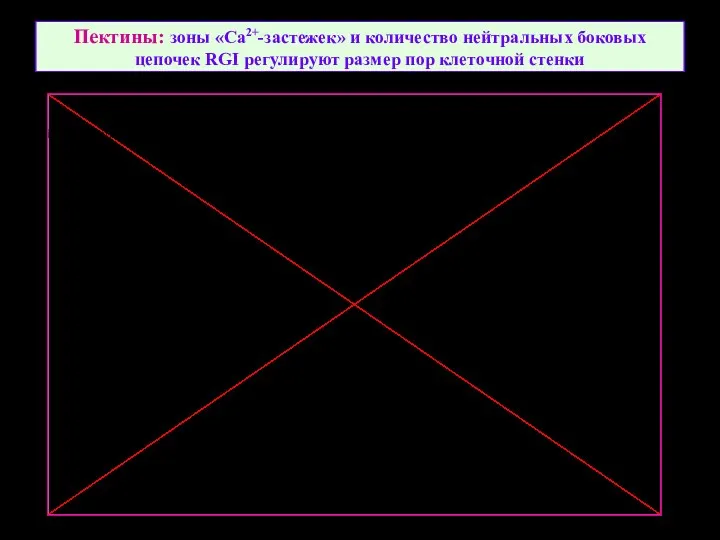
Пектины: зоны «Ca2+-застежек» и количество нейтральных боковых цепочек RGI регулируют размер
Пектины: зоны «Ca2+-застежек» и количество нейтральных боковых цепочек RGI регулируют размер

Пектины: функциональная сеть клеточной стенки
Функции пектинов:
определяют размер пор КС
определяют
Пектины: функциональная сеть клеточной стенки
Функции пектинов:
определяют размер пор КС
определяют

Структурные белки клеточной стенки:
HGRPs, PRPs, GRPs (гидроксипролин-, пролин- и глицин-
Структурные белки клеточной стенки: HGRPs, PRPs, GRPs (гидроксипролин-, пролин- и глицин-

Структурные белки клеточной стенки:
AGPs (арабино-галактановые белки - протеогликаны).
Структурные белки клеточной стенки:
AGPs (арабино-галактановые белки - протеогликаны).

Структурные белки клеточной стенки:
AGPs (арабино-галактановые белки - протеогликаны).
Структурные белки клеточной стенки:
AGPs (арабино-галактановые белки - протеогликаны).

Трехмерная модель двух типов клеточной стенки:
тип I (двудольные) и тип
Трехмерная модель двух типов клеточной стенки: тип I (двудольные) и тип
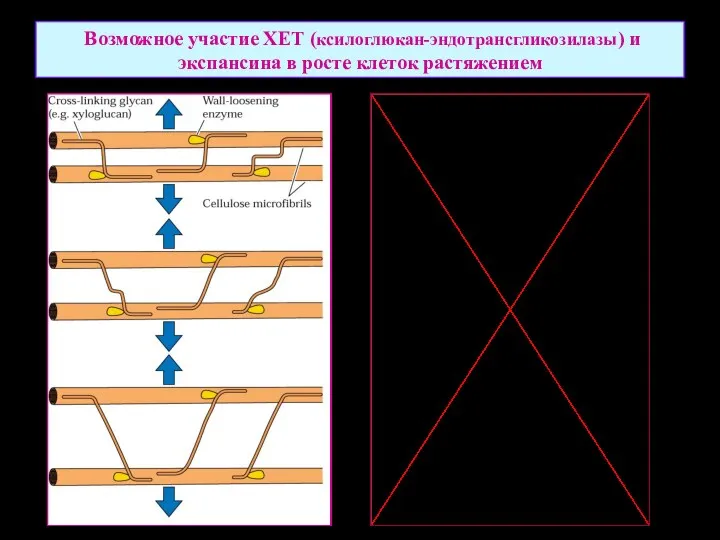
Возможное участие ХЕТ (ксилоглюкан-эндотрансгликозилазы) и экспансина в росте клеток растяжением
Возможное участие ХЕТ (ксилоглюкан-эндотрансгликозилазы) и экспансина в росте клеток растяжением

Лигнины: фенилпропаниодная сеть вторичных клеточных стенок
Лигнины: фенилпропаниодная сеть вторичных клеточных стенок
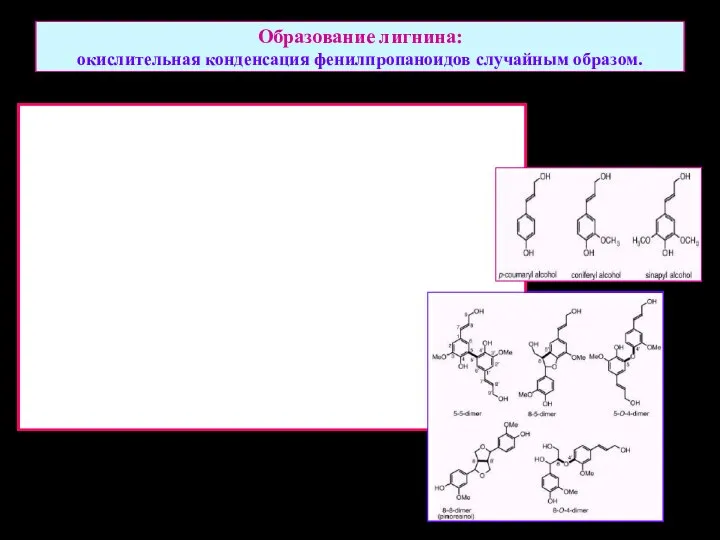
Образование лигнина:
окислительная конденсация фенилпропаноидов случайным образом.
Образование лигнина:
окислительная конденсация фенилпропаноидов случайным образом.

Образование лигнинов: целенаправленная конденсация мономеров.
Образование лигнинов: целенаправленная конденсация мономеров.

Некоторые особенности плазмалеммы
Структурные: зависимость состава от типа клетки
основные ЖК: пальмитиновая
Некоторые особенности плазмалеммы
Структурные: зависимость состава от типа клетки
основные ЖК: пальмитиновая
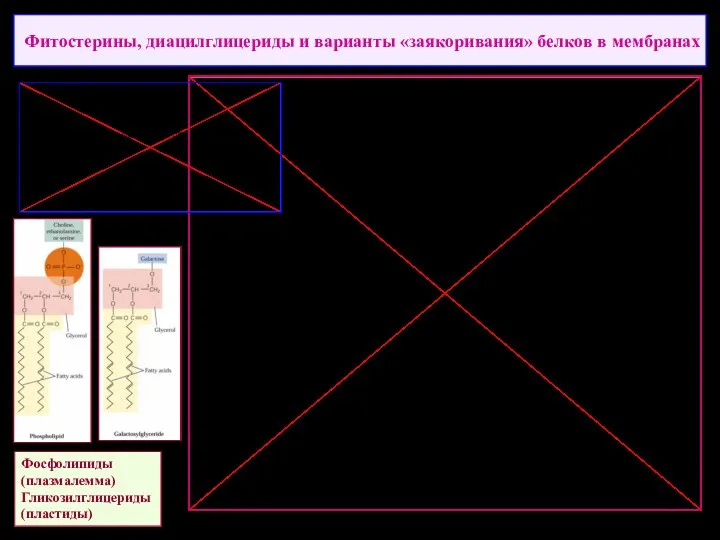
Фитостерины, диацилглицериды и варианты «заякоривания» белков в мембранах
Фосфолипиды
(плазмалемма)
Гликозилглицериды
(пластиды)
Фитостерины, диацилглицериды и варианты «заякоривания» белков в мембранах
Фосфолипиды
(плазмалемма)
Гликозилглицериды
(пластиды)





Структура растительного аппарата Гольджи
Структура растительного аппарата Гольджи

Везикулярный транспорт, типы везикул
COPII – транспорт от ER к Гольджи,
Везикулярный транспорт, типы везикул
COPII – транспорт от ER к Гольджи,

Синтез ксилоглюканов (А) и пектинов (В) проходит в разных компартментах АГ
До
Синтез ксилоглюканов (А) и пектинов (В) проходит в разных компартментах АГ
До




Некоторые особенности ядерного генома растений
Размер: от ~108 тпн (Arabidopsis) до 1010
Некоторые особенности ядерного генома растений
Размер: от ~108 тпн (Arabidopsis) до 1010
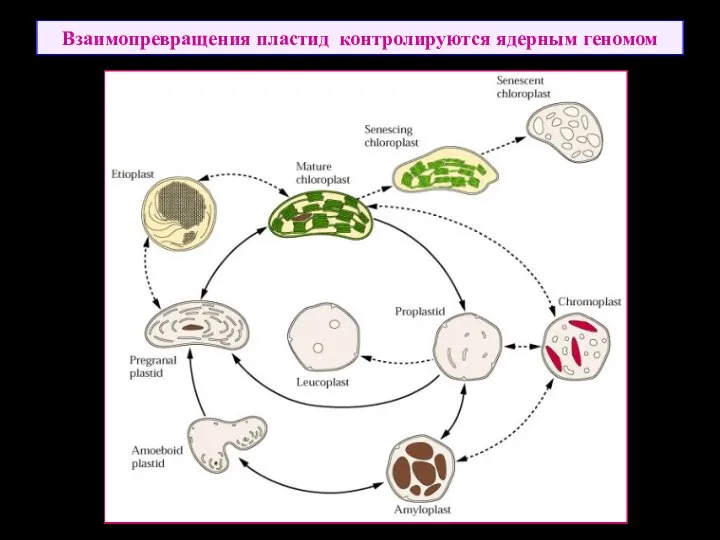
Взаимопревращения пластид контролируются ядерным геномом
Взаимопревращения пластид контролируются ядерным геномом

Хлоропласт – «главный» представитель пластид
Хлоропласт – «главный» представитель пластид

Фитоферритин в пропластидах мезофилла сои, амилопласт
Фитоферритин в пропластидах мезофилла сои, амилопласт

Этиопласт: структура проламеллярного тела, формирование хлоропласта
Этиопласт: структура проламеллярного тела, формирование хлоропласта

Структура хлоропластного генома риса.
Два типа генома:
c двумя IR размером
Структура хлоропластного генома риса.
Два типа генома:
c двумя IR размером
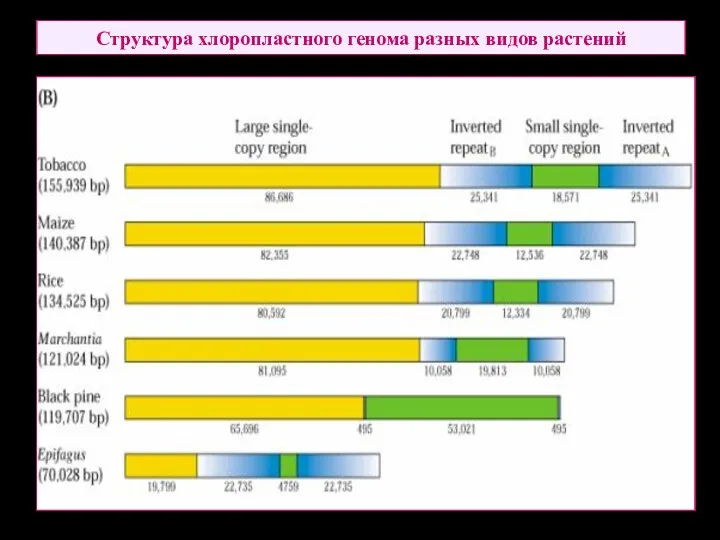
Структура хлоропластного генома разных видов растений
Структура хлоропластного генома разных видов растений

Сходства и отличия хлоропластного генома и белоксинтезирующей системы от бактериальных
Сходства:
Кольцевая ДНК
Содержание
Сходства и отличия хлоропластного генома и белоксинтезирующей системы от бактериальных
Сходства:
Кольцевая ДНК
Содержание

Гены хлоропластов
Транскрипция. 4 гена субъединиц пластидной РНК-полимеразы (rpo)
Синтез белка. - 4
Гены хлоропластов
Транскрипция. 4 гена субъединиц пластидной РНК-полимеразы (rpo)
Синтез белка. - 4
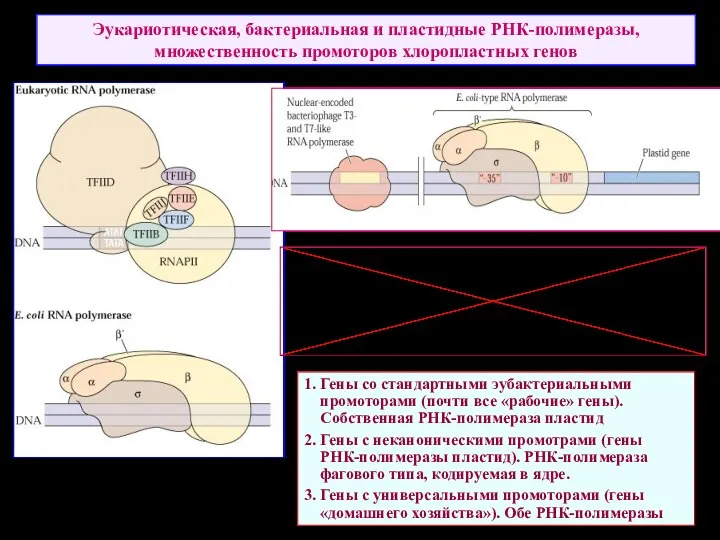
Эукариотическая, бактериальная и пластидные РНК-полимеразы, множественность промоторов хлоропластных генов
1. Гены со
Эукариотическая, бактериальная и пластидные РНК-полимеразы, множественность промоторов хлоропластных генов
1. Гены со
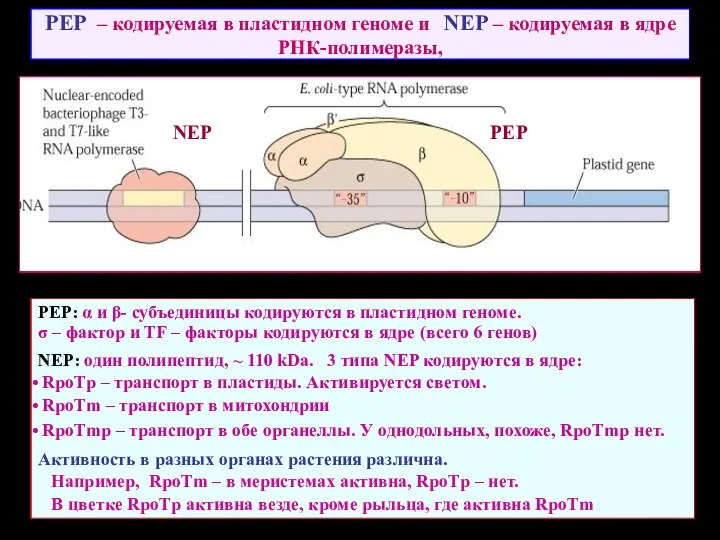
PEP – кодируемая в пластидном геноме и NEP – кодируемая в
PEP – кодируемая в пластидном геноме и NEP – кодируемая в
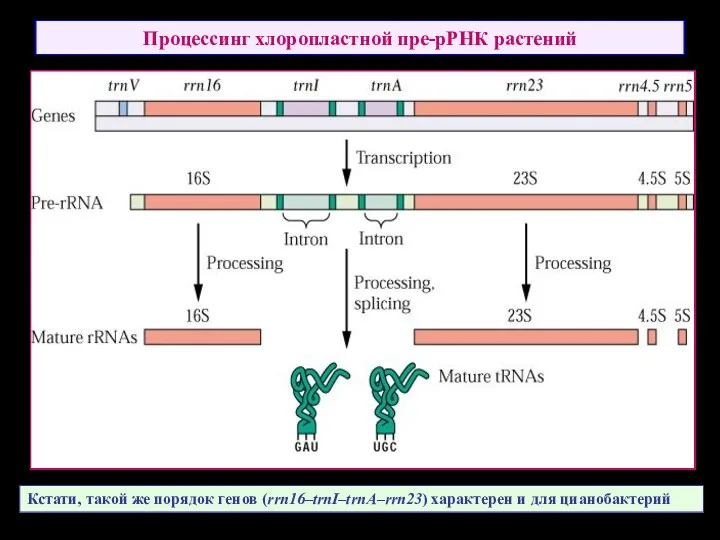
Процессинг хлоропластной пре-рРНК растений
Кстати, такой же порядок генов (rrn16–trnI–trnA–rrn23) характерен
Процессинг хлоропластной пре-рРНК растений
Кстати, такой же порядок генов (rrn16–trnI–trnA–rrn23) характерен

Структуры зрелой пластидной и ядерной иРНК.
Полиаденилирование выполняет для них функции с
Структуры зрелой пластидной и ядерной иРНК. Полиаденилирование выполняет для них функции с

Функции пластид
Фотосинтез – NB
Синтез: все жирные кислоты, многие аминокислоты, синтез пуринов
Функции пластид
Фотосинтез – NB
Синтез: все жирные кислоты, многие аминокислоты, синтез пуринов
Растительные митохондрии имеют разнообразный размер и форму
Растительные митохондрии имеют разнообразный размер и форму
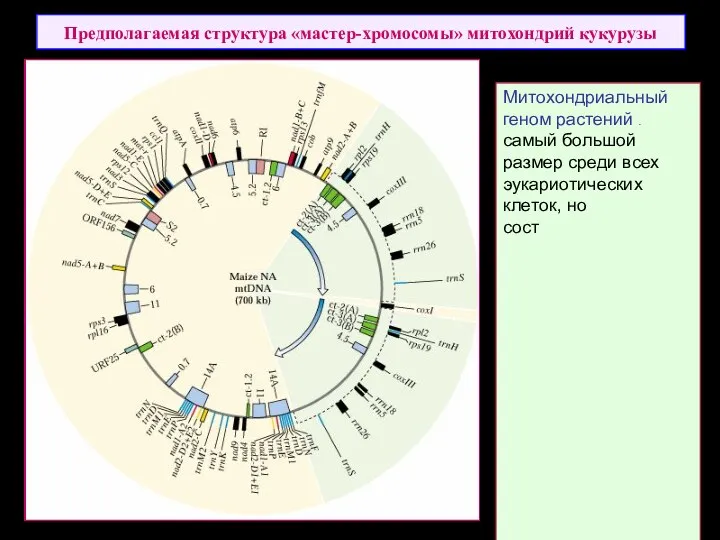
Предполагаемая структура «мастер-хромосомы» митохондрий кукурузы
Митохондриальный
геном растений имеет самый большой размер
Предполагаемая структура «мастер-хромосомы» митохондрий кукурузы
Митохондриальный
геном растений имеет самый большой размер




 Регуляция пищеварения. Заболевания органов пищеварения и их предупреждение. Биология 8 кл (Пасечник)
Регуляция пищеварения. Заболевания органов пищеварения и их предупреждение. Биология 8 кл (Пасечник) Уровни организации жизни
Уровни организации жизни 20231130_8_klass_ptitsy._obshchie_priznaki._osobennosti_stroeniya._1
20231130_8_klass_ptitsy._obshchie_priznaki._osobennosti_stroeniya._1 Папоротеподібні. Загальна характеристика
Папоротеподібні. Загальна характеристика Тип Кольчатые черви
Тип Кольчатые черви Безусловные рефлексы
Безусловные рефлексы Зоология беспозвоночных
Зоология беспозвоночных Химический состав клетки
Химический состав клетки Немембранные и двумембранные органоиды
Немембранные и двумембранные органоиды Фармацевтична ботаніка. Аналіз буклету 2015 року
Фармацевтична ботаніка. Аналіз буклету 2015 року Проектування і створення заповідних територій
Проектування і створення заповідних територій Мутационная изменчивость
Мутационная изменчивость Неорганические вещества, входящие в состав клетки
Неорганические вещества, входящие в состав клетки Понятие евгеники
Понятие евгеники Молекулярные основы наследственности. Биосинтез белка. Транскрипция
Молекулярные основы наследственности. Биосинтез белка. Транскрипция Фізіологічні особливості прояву рухових якостей в спортивному і оздоровчому тренуванні. Витривалість. (Лекція 7)
Фізіологічні особливості прояву рухових якостей в спортивному і оздоровчому тренуванні. Витривалість. (Лекція 7) Транспортные системы организма человека
Транспортные системы организма человека Систематика основных тканей растений
Систематика основных тканей растений Адаптаційні зміни в організмі людини, як основа розвитку тренувальних ефектів при заняттях фізичними вправами
Адаптаційні зміни в організмі людини, як основа розвитку тренувальних ефектів при заняттях фізичними вправами Выделение. Органы выделения
Выделение. Органы выделения Возрастная анатомия и физиология
Возрастная анатомия и физиология Презентация к уроку биологии 5 кл.
Презентация к уроку биологии 5 кл. Приспособленность растений к условиям жизни в лесном биогеоценозе.Лабораторная работа.
Приспособленность растений к условиям жизни в лесном биогеоценозе.Лабораторная работа. Тип Хордовые (chordata)
Тип Хордовые (chordata) Энергообразующие органоиды (лекция 7)
Энергообразующие органоиды (лекция 7) Дыхание растений
Дыхание растений Общая характеристика транспортных процессов в клетке. Пассивный транспорт
Общая характеристика транспортных процессов в клетке. Пассивный транспорт Двойное оплодотворение у цветковых растений
Двойное оплодотворение у цветковых растений