- Основы анатомии и физиологии

Содержание
- 2. Гладкие мышцы Скелетные мышцы Сердечная мышца Типы мышечной ткани все типы – генерация силы / совершение
- 3. Уровни организации скелетной мышцы Миофибрилла Волокна (клетки) скелетных мышц очень крупные: диаметр - до 100 мкм,
- 4. Саркомер – структурно-функциональная единица миофибриллы Z-линии разных миофибрилл расположены на одном уровне 2мкм
- 5. Белки саркомера α-актинин Миозин Титин (тайтин) Актин Титин - гигантский белок (3-3,7 МДа), крупнейший из белков
- 6. Строение тонкого филамента 2 цепи, обвивающие друг друга – 1 поворот спирали = 13 G-актинам 2
- 7. Мышечная дистрофия Дюшенна (1861) 1:3500 ♂ Комплекс: Дистрофин-гликопротеины ? Взаимодействие между субсарколеммальным цитоскелетом и внеклеточным матриксом
- 8. Строение толстого филамента (миозин II) 2 тримера (1 тяжелая и 2 легких цепи) шарнирные участки 1
- 9. Актин-связывающий центр Строение головки миозина Головки миозина образуют поперечные мостики между толстыми и тонкими филаментами АТФ-связывающий
- 10. Теория скольжения нитей: во время сокращения мышцы длина толстых и тонких филаментов не изменяется, происходит их
- 11. Цикл работы головки миозина обеспечение повторения цикла энергия для движения поперечного мостика сдвиг на 11 нм
- 12. Сила сокращения скелетной мышцы зависит от длины саркомеров (степени перекрывания актиновых и миозиновых филаментов)
- 13. Расположение тропомиозина и тропонинового комплекса на актиновом филаменте Связывание Са2+ с тропонином С ослабляет взаимодействие тропонинового
- 14. Взаимодействие головок миозина с актином возможно лишь при повышении концентрации Са2+ в цитоплазме
- 15. 1. Потенциал действия 3. Связывание Са2+ c тропонином 4. Сокращение 2. Повышение внутриклеточной концентрации Ca2+ Электромеханическое
- 16. Поперечные трубочки (Т-трубочки) и саркоплазматический ретикулум (СПР)
- 17. Поперечные трубочки (Т-трубочки) и саркоплазматический ретикулум (СПР)
- 18. Поперечные трубочки (Т-трубочки) и саркоплазматический ретикулум (СПР) Три гипотезы о механизме передачи сигнала от Т-трубочек к
- 19. Гипотеза о механическом способе передачи сигнала от наружной мембраны к СПР: Chandler, Rakowski and Schneider (1976)
- 20. Соединительные ножки между мембранами Т-трубочки и СПР
- 21. «Наружная» часть ножки – ДИГИДРОПИРИДИНОВЫЙ РЕЦЕПТОР (потенциал-зависимый кальциевый канал L-типа) Цитоплазматическая петля, формирующая ножку («foot»), взаимодействующую
- 22. Ryania speciosa Трехмерная модель рианодинового рецептора, построенная по данным электронной микроскопии Вид сбоку Вид со стороны
- 23. Взаимное расположение дигидропиридиновых (ДГП) и рианодиновых рецепторов в скелетной мышце
- 24. А.М.Рубцов Соросовский образовательный журнал 2000, том 6, №9, с.17-24 Активация рианодиновых рецепторов Типы рианодиновых рецепторов: cкелетная
- 25. А.М.Рубцов Соросовский образовательный журнал 2000, том 6, №9, с.17-24 Активация рианодиновых рецепторов Типы рианодиновых рецепторов: cкелетная
- 27. Динамика одиночного сокращения различается для разных мышц Икроножная мышца млекопитающих (m. gastrochnemius) Камбаловидная мышца млекопитающих (m.soleus)
- 28. Типы мышечных волокон Изоформы тяжелых цепей различаются по АТФ-азной активности (количеству молекул АТФ, расщепляемых в единицу
- 29. Гликолитические и оксидативные мышечные волокна различаются по размерам и активности митохондриальных ферментов
- 30. Гликолитические и оксидативные волокна различаются по устойчивости к утомлению
- 31. Свойства различных типов фазических мышечных волокон
- 32. ПД, приходящие по аксону мотонейрона вызывают только сокращение мышцы ( а не торможение сокращения) Нервно-мышечный синапс
- 33. Двигательные (моторные) единицы Двигательная единица = мотонейрон + группа иннервируемых им мышечных волокон Все мышечные волокна
- 34. Размер и количество ДЕ в мышцах человека Наружная прямая мышца глаза Икроножная
- 35. Два способа увеличения силы сокращения мышцы: Активация большего числа мотонейронов (рекрутирование ДЕ) Увеличение частоты разрядов мотонейронов:
- 36. Метаболический тип мышечного волокна определяется его иннервацией (характером разрядной активности мотонейрона) Изменение свойств мышц при перекрестной
- 37. Изменения строения мышечной системы при естественной активности мотонейронов Тренировка силы (тяжелая атлетика) Тренировка выносливости (марафон)
- 38. Источники энергии в скелетной мышце человека
- 39. Гладкие мышцы Скелетные мышцы Сердечная мышца Типы мышечной ткани сокращения (произвольные) ? мотонейроны сокращения ? ВНС,
- 40. Гладкая мышца Нет поперечной полосатости (регулярной исчерченности) Веретеновидная форма, 1 ядро Актина > чем миозина (в
- 42. Гладкая мышца Типы гладкомышечной ткани Мультиунитарная Унитарная Цилиарная мышца, мышца радужки, пилоэректоры, семявыносящий проток, крупные артерии
- 43. Фазический и тонический типы гладких мышц Фазические: сокращению предшествует ПД (матка, мочевой пузырь, вены, «трубчатые» отделы
- 44. Потенциал покоя и потенциал действия гладкомышечных клеток Потенциал покоя ниже, чем в скелетной и сердечной мышцах
- 45. Потенциал покоя и потенциал действия гладкомышечных клеток Некоторые другие ионные каналы, которые важны для работы гладкой
- 46. Два механизма запуска сокращения гладкой мышцы Что такое «рецептор-управляемые кальциевые каналы»? Вход Са2+ приводит к деполяризации
- 47. TRP-каналы в сосудах Store-operated channels (депо-управляемые каналы) Receptor-operated channels (рецептор-управляемые каналы) Stretch-activated channels (каналы, активируемые растяжением)
- 48. Основные пути активации актомиозинового взаимодействия Актин = «рельсы» Миозиновый мотор КПД 50-80% (у двигателей внутреннего сгорания
- 49. Сигнальные пути, регулирующие взаимодействие актина и миозина в гладкомышечных клетках В отличие от скелетной и сердечной
- 50. Сигнальные пути, регулирующие взаимодействие актина и миозина в гладкомышечных клетках В отличие от скелетной и сердечной
- 51. Сравнение динамики сокращения скелетной, сердечной и гладкой мышц Низкая АТФазная активность миозина
- 52. Главные механизмы, влияющие на внутриклеточную концентрацию кальция в ГМК
- 55. Скачать презентацию
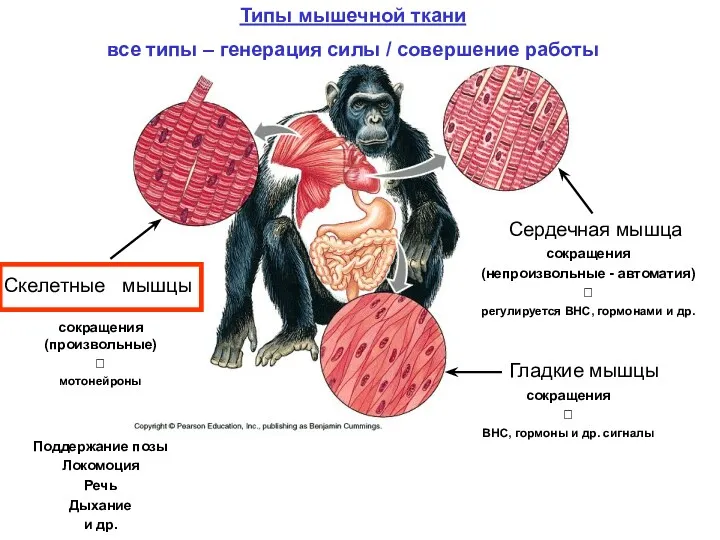
Гладкие мышцы
Скелетные мышцы
Сердечная мышца
Типы мышечной ткани
все типы – генерация силы /
Гладкие мышцы
Скелетные мышцы
Сердечная мышца
Типы мышечной ткани
все типы – генерация силы /
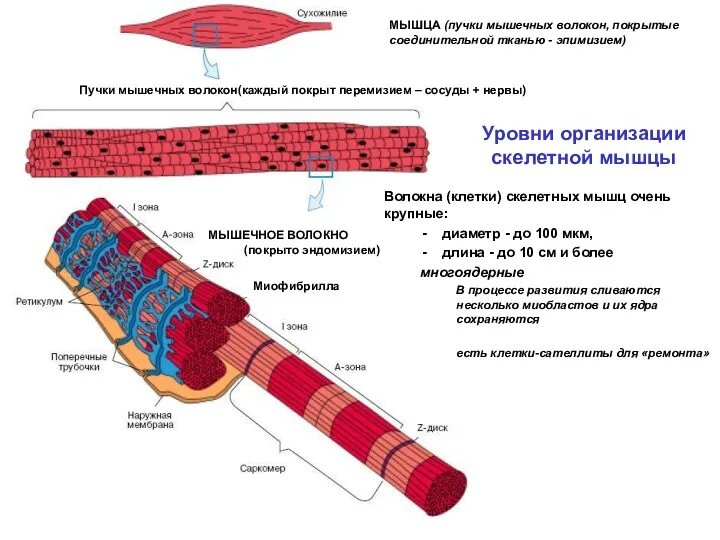
Уровни организации
скелетной мышцы
Миофибрилла
Волокна (клетки) скелетных мышц очень крупные:
диаметр - до
Уровни организации
скелетной мышцы
Миофибрилла
Волокна (клетки) скелетных мышц очень крупные:
диаметр - до
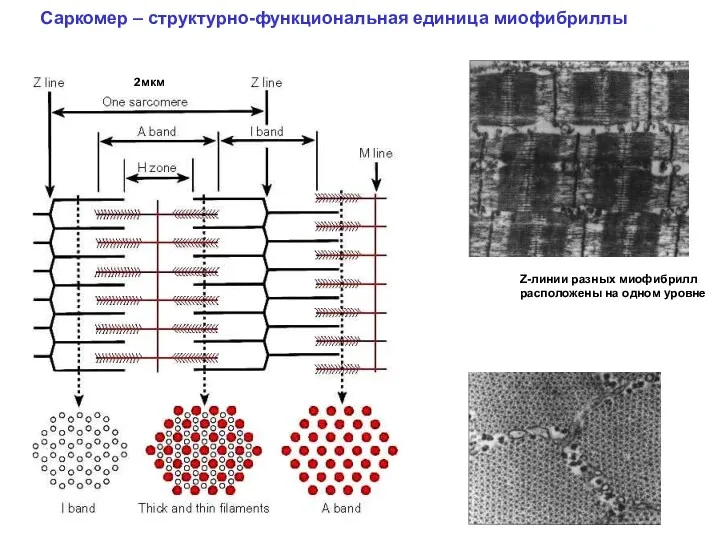
Саркомер – структурно-функциональная единица миофибриллы
Z-линии разных миофибрилл расположены на одном
Саркомер – структурно-функциональная единица миофибриллы
Z-линии разных миофибрилл расположены на одном
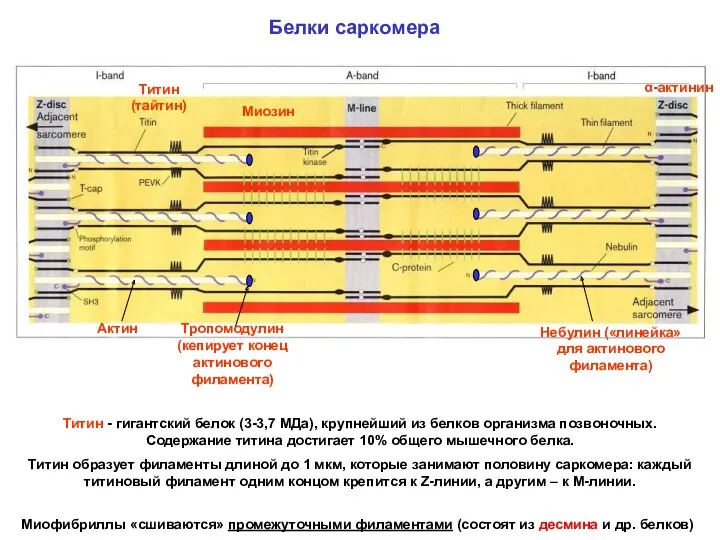
Белки саркомера
α-актинин
Миозин
Титин (тайтин)
Актин
Титин - гигантский белок (3-3,7 МДа), крупнейший из белков
Белки саркомера
α-актинин
Миозин
Титин (тайтин)
Актин
Титин - гигантский белок (3-3,7 МДа), крупнейший из белков
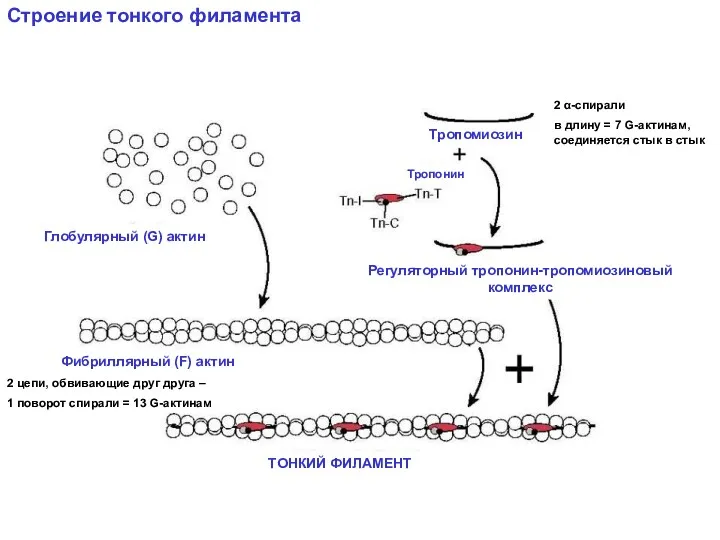
Строение тонкого филамента
2 цепи, обвивающие друг друга –
1 поворот
Строение тонкого филамента
2 цепи, обвивающие друг друга –
1 поворот
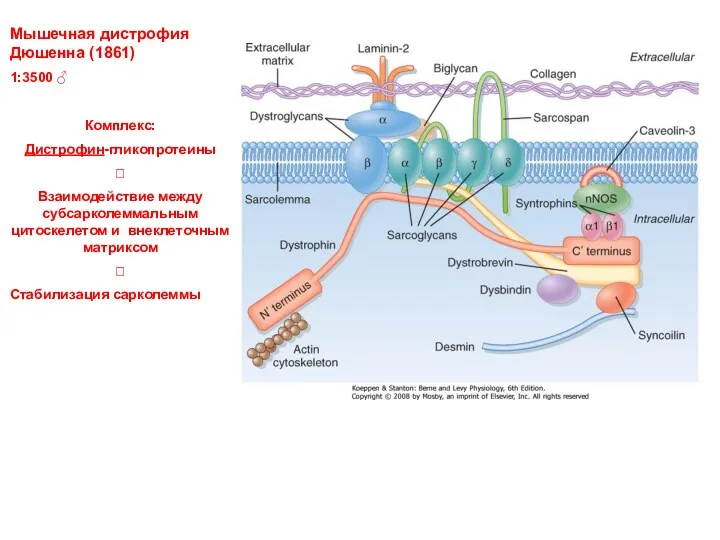
Мышечная дистрофия Дюшенна (1861)
1:3500 ♂
Комплекс:
Дистрофин-гликопротеины
?
Взаимодействие между субсарколеммальным цитоскелетом и
Мышечная дистрофия Дюшенна (1861)
1:3500 ♂
Комплекс:
Дистрофин-гликопротеины
?
Взаимодействие между субсарколеммальным цитоскелетом и
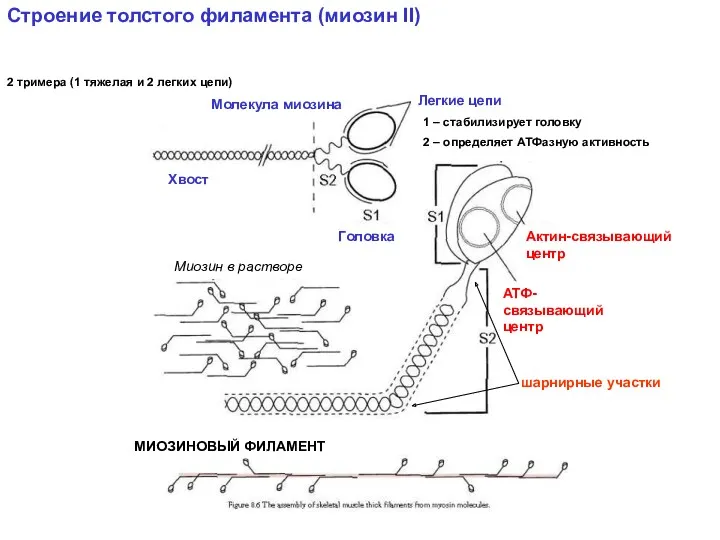
Строение толстого филамента (миозин II)
2 тримера (1 тяжелая и 2
Строение толстого филамента (миозин II)
2 тримера (1 тяжелая и 2
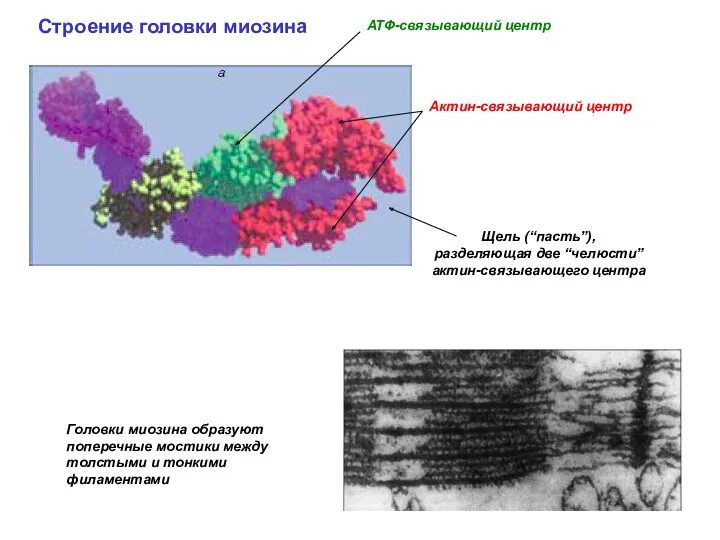
Актин-связывающий центр
Строение головки миозина
Головки миозина образуют поперечные мостики между толстыми и
Актин-связывающий центр
Строение головки миозина
Головки миозина образуют поперечные мостики между толстыми и

Теория скольжения нитей:
во время сокращения мышцы длина толстых и тонких
Теория скольжения нитей:
во время сокращения мышцы длина толстых и тонких
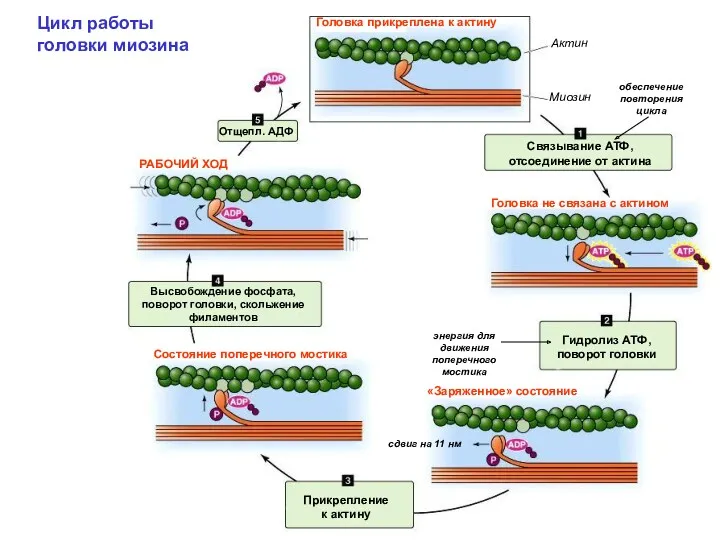
Цикл работы головки миозина
обеспечение повторения цикла
энергия для движения поперечного мостика
сдвиг на
Цикл работы головки миозина
обеспечение повторения цикла
энергия для движения поперечного мостика
сдвиг на
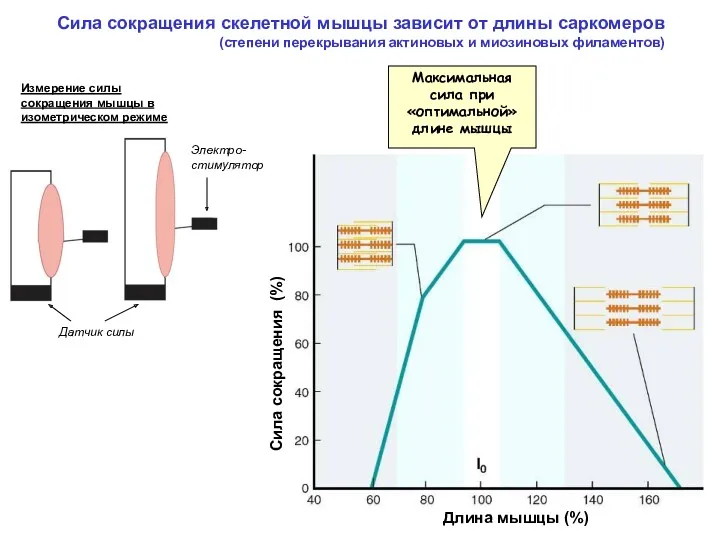
Сила сокращения скелетной мышцы зависит от длины саркомеров
(степени перекрывания актиновых
Сила сокращения скелетной мышцы зависит от длины саркомеров (степени перекрывания актиновых
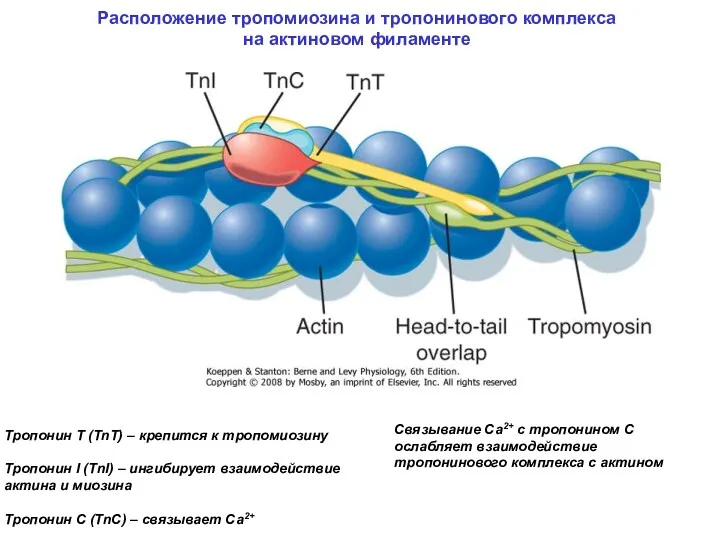
Расположение тропомиозина и тропонинового комплекса на актиновом филаменте
Связывание Са2+ с тропонином
Расположение тропомиозина и тропонинового комплекса на актиновом филаменте
Связывание Са2+ с тропонином
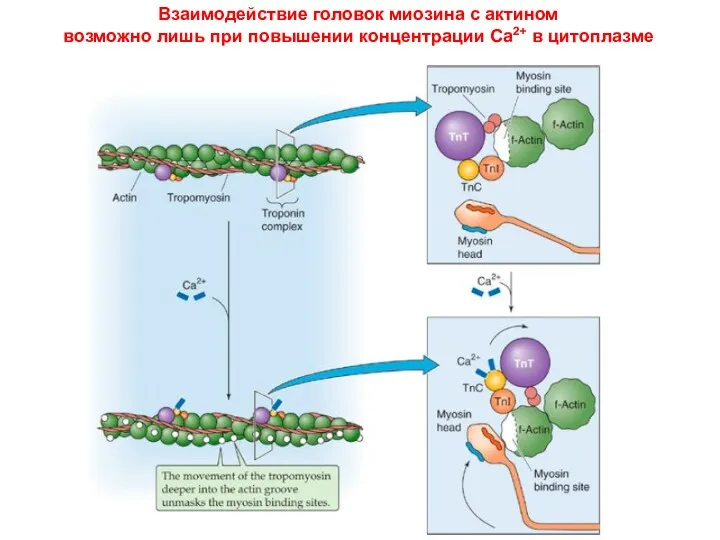
Взаимодействие головок миозина с актином
возможно лишь при повышении концентрации Са2+
Взаимодействие головок миозина с актином возможно лишь при повышении концентрации Са2+
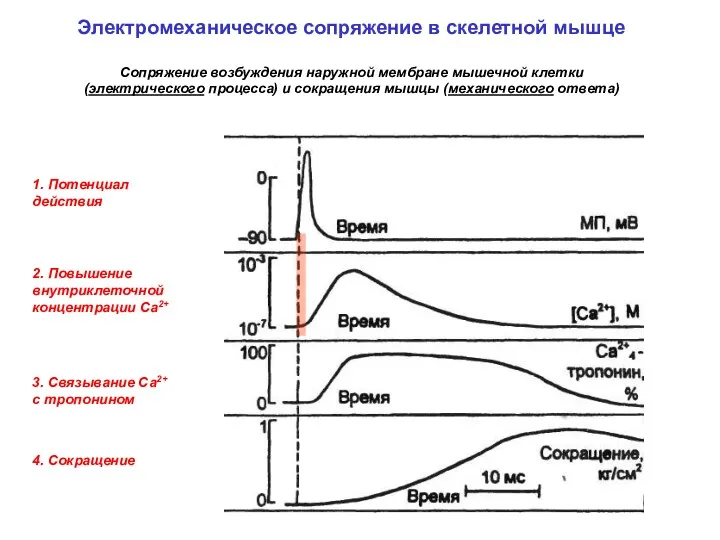
1. Потенциал действия
3. Связывание Са2+
c тропонином
4. Сокращение
2. Повышение внутриклеточной концентрации
1. Потенциал действия
3. Связывание Са2+
c тропонином
4. Сокращение
2. Повышение внутриклеточной концентрации
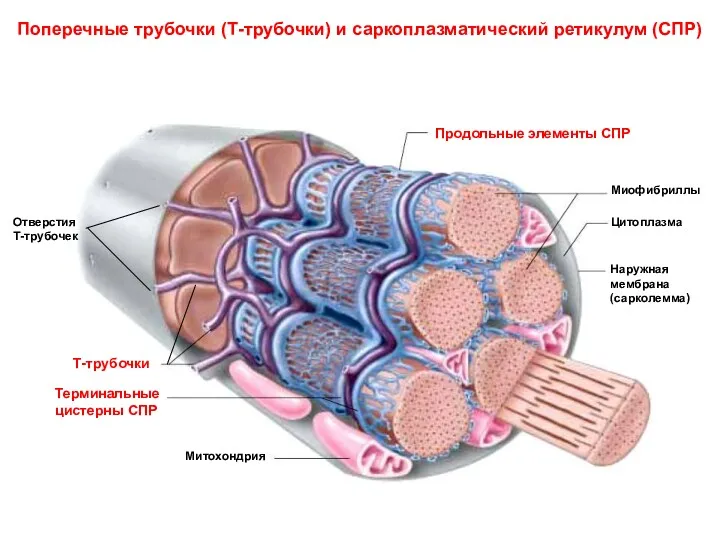
Поперечные трубочки (Т-трубочки) и саркоплазматический ретикулум (СПР)
Поперечные трубочки (Т-трубочки) и саркоплазматический ретикулум (СПР)
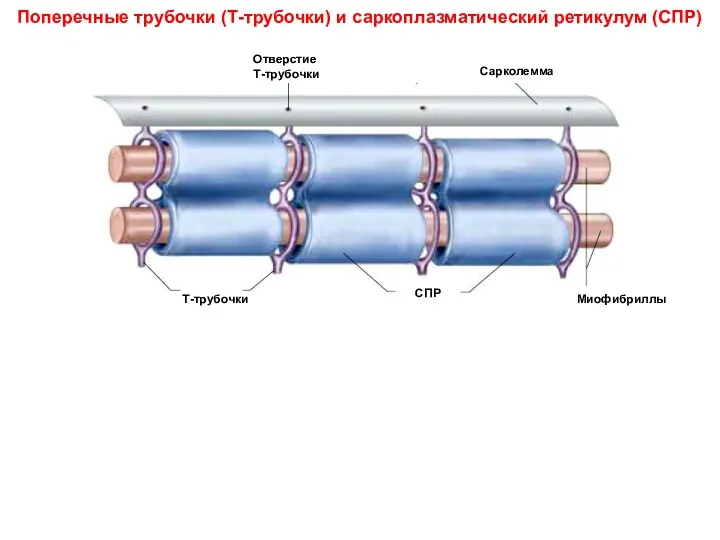
Поперечные трубочки (Т-трубочки) и саркоплазматический ретикулум (СПР)
Поперечные трубочки (Т-трубочки) и саркоплазматический ретикулум (СПР)
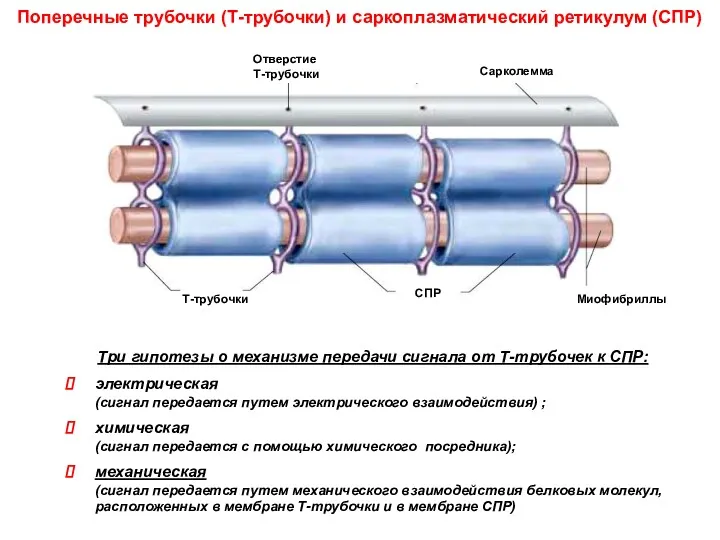
Поперечные трубочки (Т-трубочки) и саркоплазматический ретикулум (СПР)
Три гипотезы о механизме
Поперечные трубочки (Т-трубочки) и саркоплазматический ретикулум (СПР)
Три гипотезы о механизме
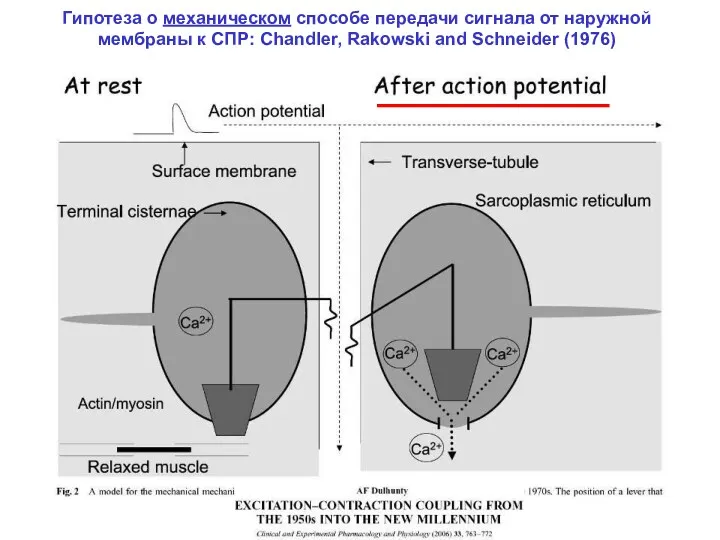
Гипотеза о механическом способе передачи сигнала от наружной мембраны к СПР:
Гипотеза о механическом способе передачи сигнала от наружной мембраны к СПР:
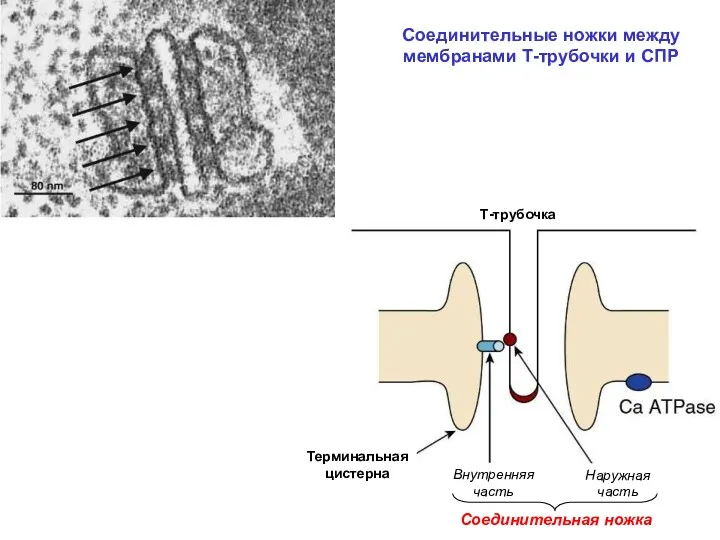
Соединительные ножки между мембранами Т-трубочки и СПР
Соединительные ножки между мембранами Т-трубочки и СПР
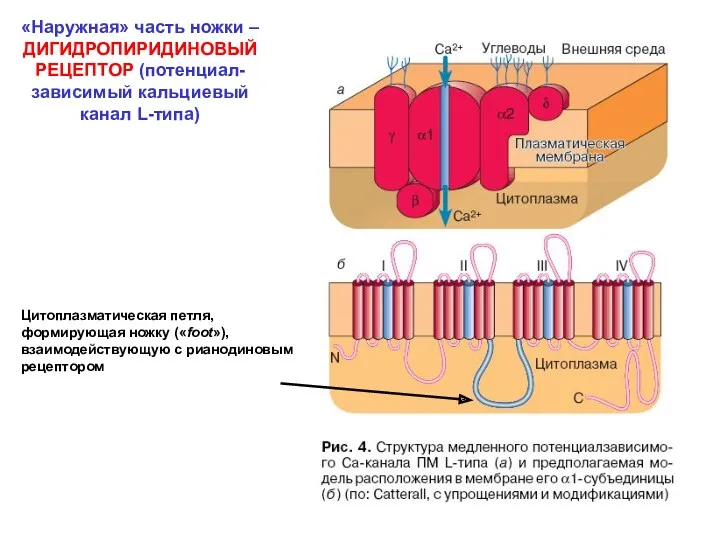
«Наружная» часть ножки – ДИГИДРОПИРИДИНОВЫЙ РЕЦЕПТОР (потенциал-зависимый кальциевый канал L-типа)
Цитоплазматическая петля,
«Наружная» часть ножки – ДИГИДРОПИРИДИНОВЫЙ РЕЦЕПТОР (потенциал-зависимый кальциевый канал L-типа)
Цитоплазматическая петля,
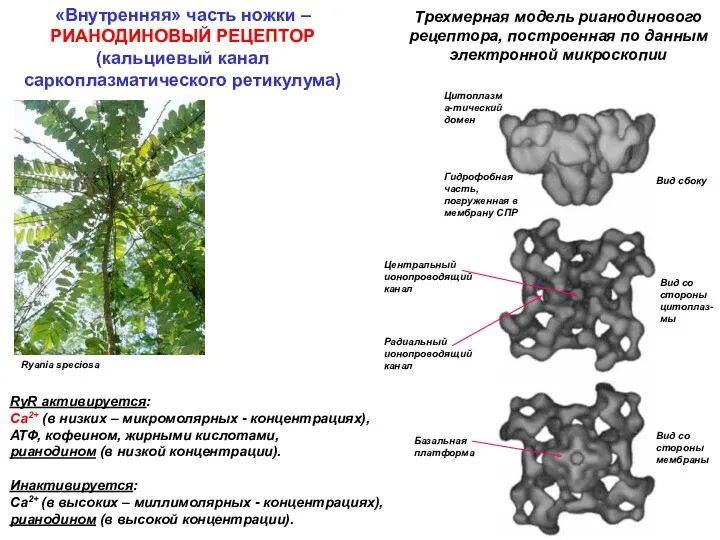
Ryania speciosa
Трехмерная модель рианодинового рецептора, построенная по данным электронной микроскопии
Вид сбоку
Вид
Ryania speciosa
Трехмерная модель рианодинового рецептора, построенная по данным электронной микроскопии
Вид сбоку
Вид
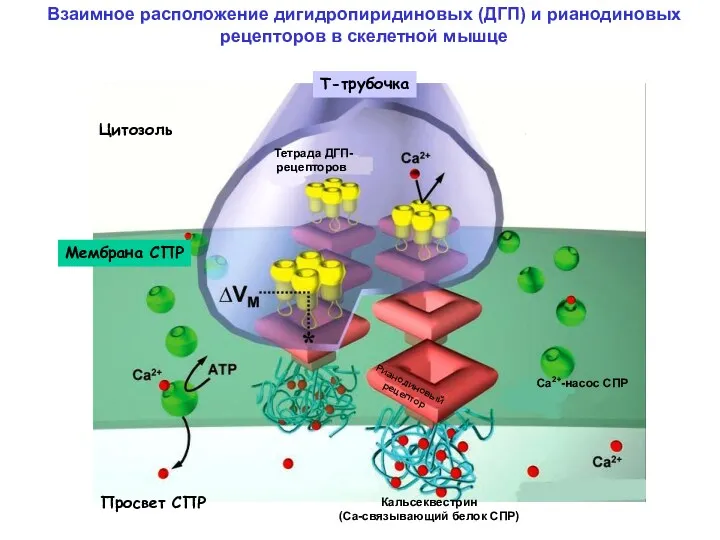
Взаимное расположение дигидропиридиновых (ДГП) и рианодиновых рецепторов в скелетной мышце
Взаимное расположение дигидропиридиновых (ДГП) и рианодиновых рецепторов в скелетной мышце
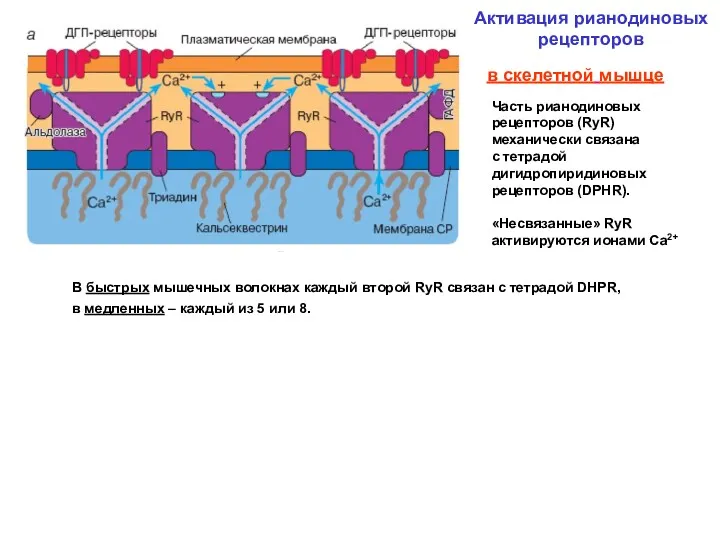
А.М.Рубцов Соросовский образовательный журнал 2000, том 6, №9, с.17-24
Активация рианодиновых рецепторов
Типы
А.М.Рубцов Соросовский образовательный журнал 2000, том 6, №9, с.17-24
Активация рианодиновых рецепторов
Типы
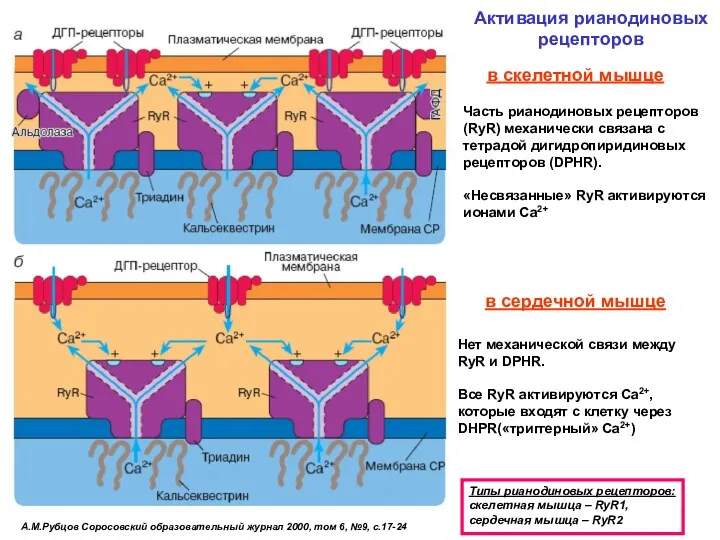
А.М.Рубцов Соросовский образовательный журнал 2000, том 6, №9, с.17-24
Активация рианодиновых рецепторов
Типы
А.М.Рубцов Соросовский образовательный журнал 2000, том 6, №9, с.17-24
Активация рианодиновых рецепторов
Типы
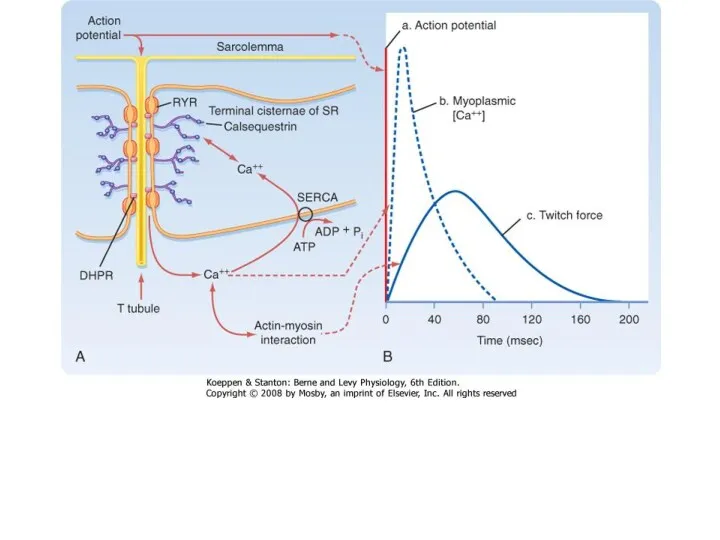
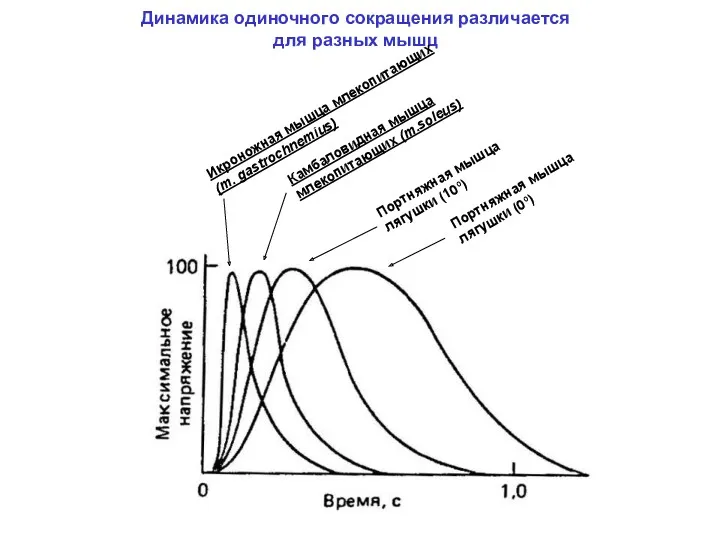
Динамика одиночного сокращения различается
для разных мышц
Икроножная мышца млекопитающих (m. gastrochnemius)
Камбаловидная
Динамика одиночного сокращения различается
для разных мышц
Икроножная мышца млекопитающих (m. gastrochnemius)
Камбаловидная

Типы мышечных волокон
Изоформы тяжелых цепей различаются по АТФ-азной активности (количеству молекул
Типы мышечных волокон
Изоформы тяжелых цепей различаются по АТФ-азной активности (количеству молекул

Гликолитические и оксидативные мышечные волокна различаются
по размерам и активности митохондриальных
Гликолитические и оксидативные мышечные волокна различаются по размерам и активности митохондриальных
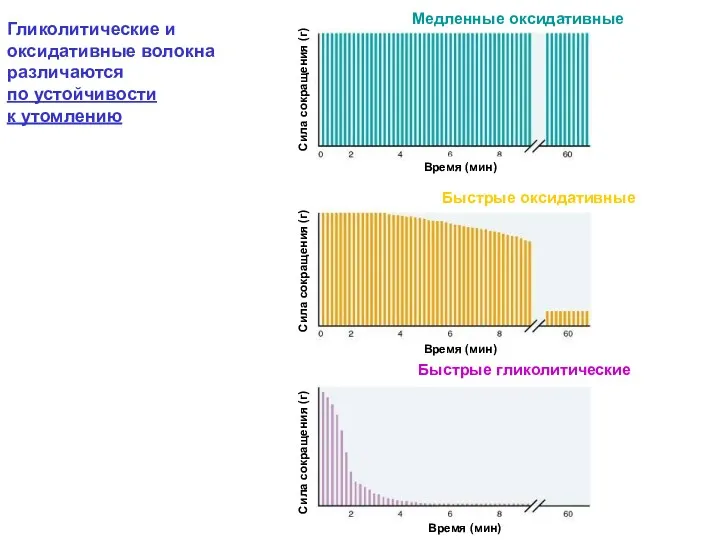
Гликолитические и оксидативные волокна различаются
по устойчивости
к утомлению
Гликолитические и оксидативные волокна различаются
по устойчивости
к утомлению
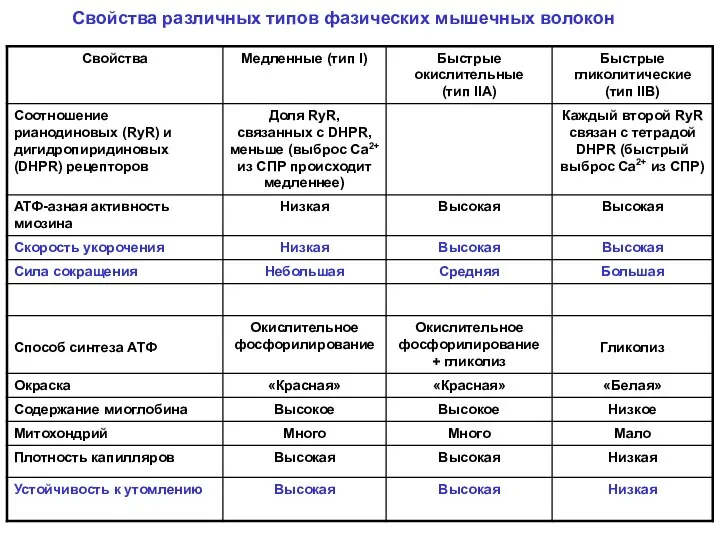
Свойства различных типов фазических мышечных волокон
Свойства различных типов фазических мышечных волокон
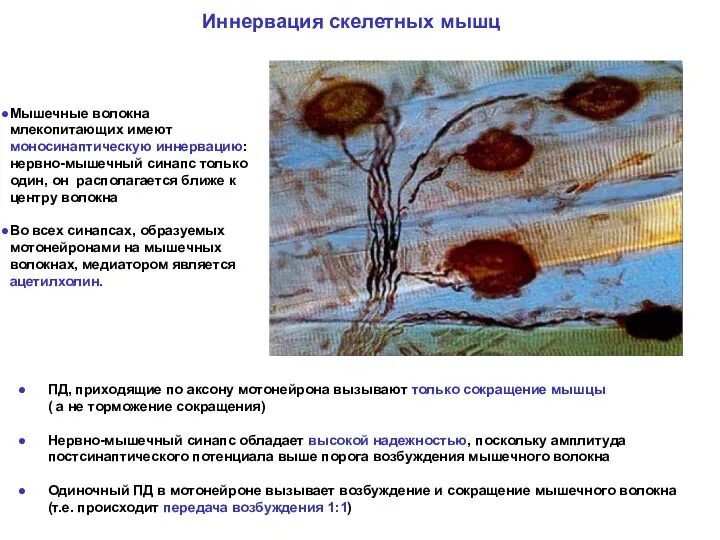
ПД, приходящие по аксону мотонейрона вызывают только сокращение мышцы
( а
ПД, приходящие по аксону мотонейрона вызывают только сокращение мышцы ( а

Двигательные (моторные) единицы
Двигательная единица = мотонейрон + группа иннервируемых им
мышечных
Двигательные (моторные) единицы
Двигательная единица = мотонейрон + группа иннервируемых им мышечных

Размер и количество ДЕ в мышцах человека
Наружная прямая мышца глаза
Икроножная
Размер и количество ДЕ в мышцах человека
Наружная прямая мышца глаза
Икроножная
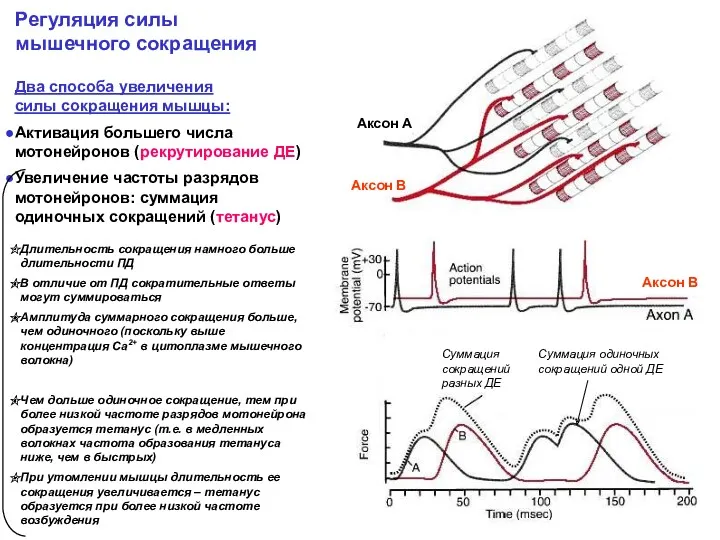
Два способа увеличения
силы сокращения мышцы:
Активация большего числа мотонейронов (рекрутирование ДЕ)
Увеличение
Два способа увеличения
силы сокращения мышцы:
Активация большего числа мотонейронов (рекрутирование ДЕ)
Увеличение
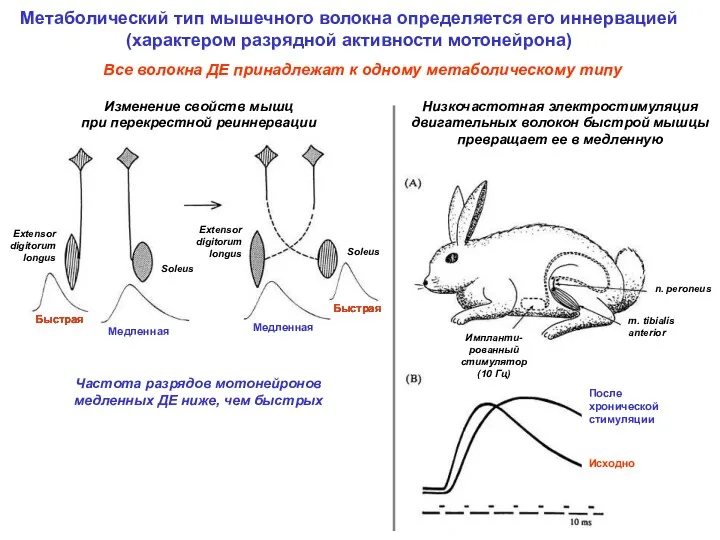
Метаболический тип мышечного волокна определяется его иннервацией
(характером разрядной активности мотонейрона)
Изменение
Метаболический тип мышечного волокна определяется его иннервацией
(характером разрядной активности мотонейрона)
Изменение
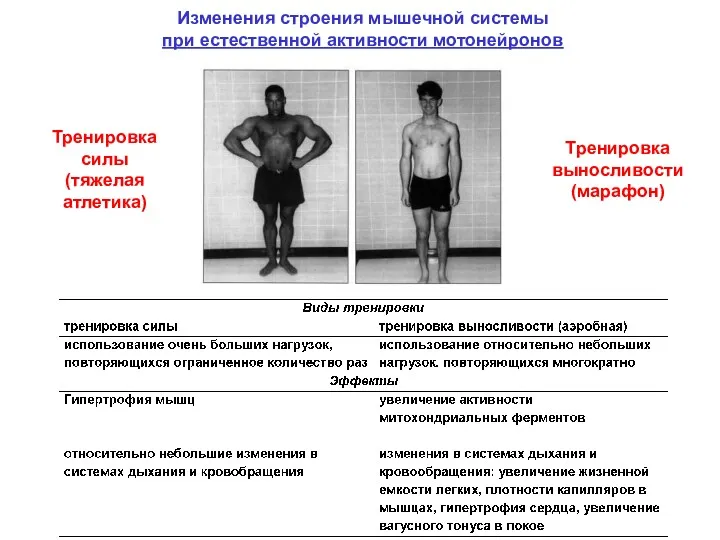
Изменения строения мышечной системы
при естественной активности мотонейронов
Тренировка силы
(тяжелая атлетика)
Тренировка выносливости
(марафон)
Изменения строения мышечной системы
при естественной активности мотонейронов
Тренировка силы
(тяжелая атлетика)
Тренировка выносливости
(марафон)
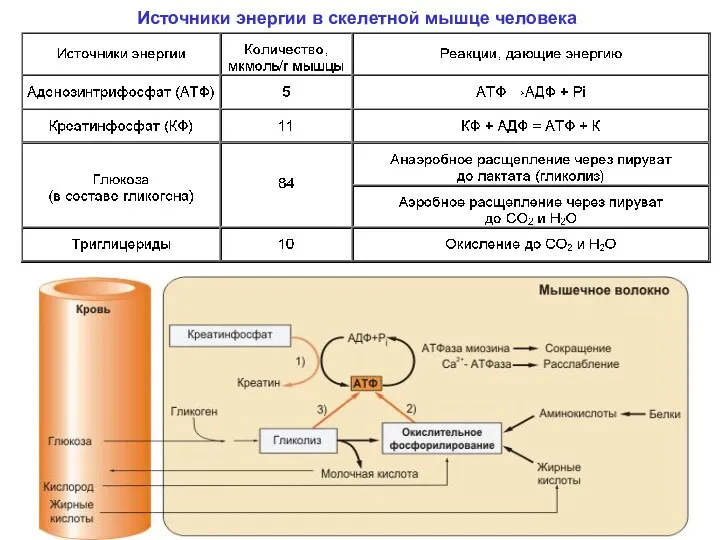
Источники энергии в скелетной мышце человека
Источники энергии в скелетной мышце человека
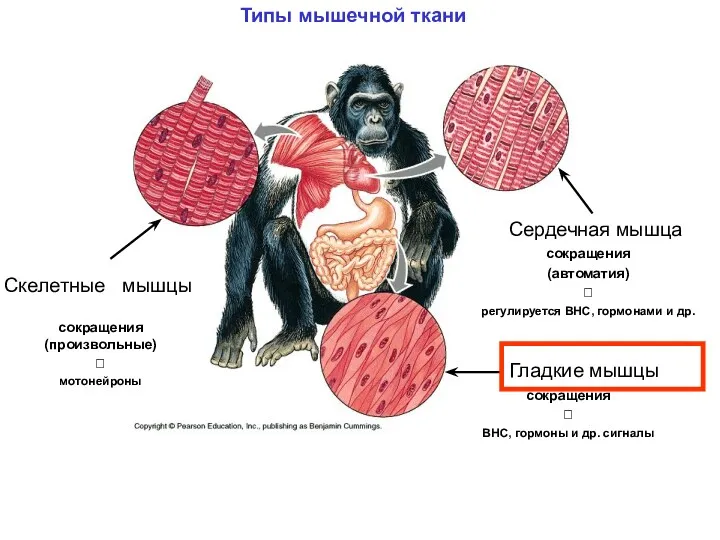
Гладкие мышцы
Скелетные мышцы
Сердечная мышца
Типы мышечной ткани
сокращения (произвольные)
?
мотонейроны
сокращения
?
ВНС, гормоны и др. сигналы
сокращения
(автоматия)
?
регулируется
Гладкие мышцы
Скелетные мышцы
Сердечная мышца
Типы мышечной ткани
сокращения (произвольные)
?
мотонейроны
сокращения
?
ВНС, гормоны и др. сигналы
сокращения
(автоматия)
?
регулируется
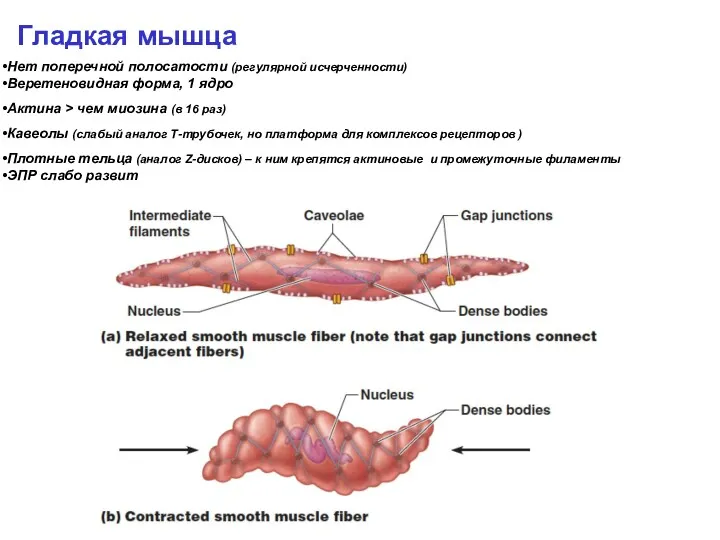
Гладкая мышца
Нет поперечной полосатости (регулярной исчерченности)
Веретеновидная форма, 1 ядро
Актина > чем
Гладкая мышца
Нет поперечной полосатости (регулярной исчерченности)
Веретеновидная форма, 1 ядро
Актина > чем

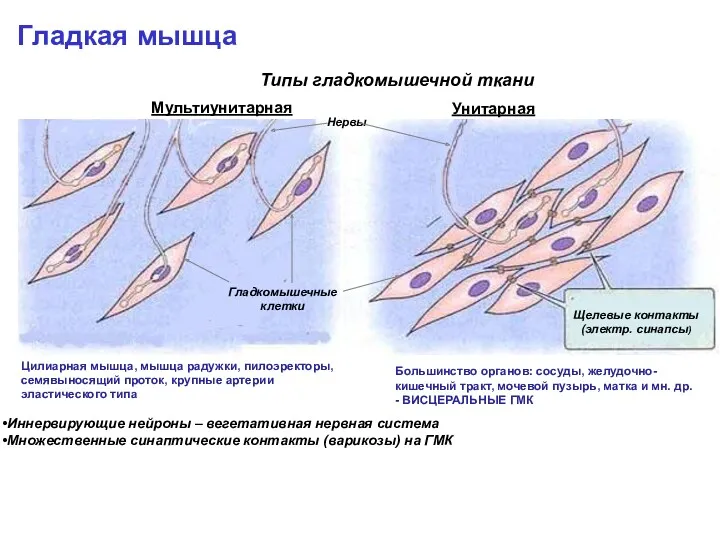
Гладкая мышца
Типы гладкомышечной ткани
Мультиунитарная
Унитарная
Цилиарная мышца, мышца радужки, пилоэректоры, семявыносящий проток, крупные
Гладкая мышца
Типы гладкомышечной ткани
Мультиунитарная
Унитарная
Цилиарная мышца, мышца радужки, пилоэректоры, семявыносящий проток, крупные
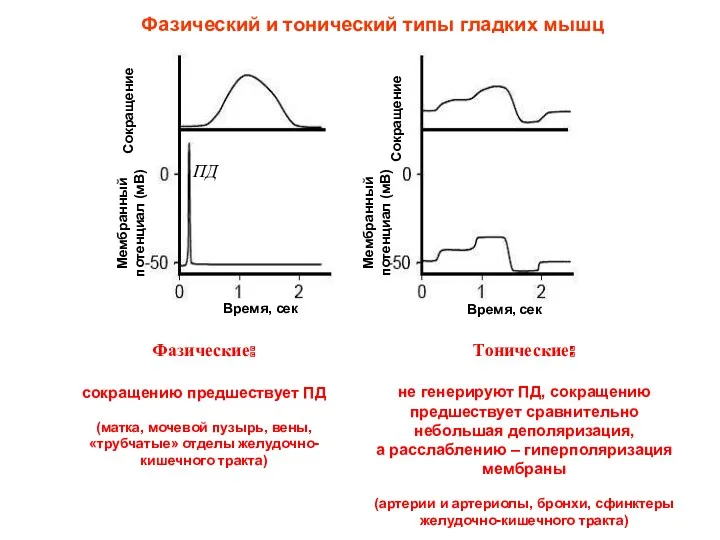
Фазический и тонический типы гладких мышц
Фазические:
сокращению предшествует ПД
(матка, мочевой
Фазический и тонический типы гладких мышц
Фазические:
сокращению предшествует ПД
(матка, мочевой
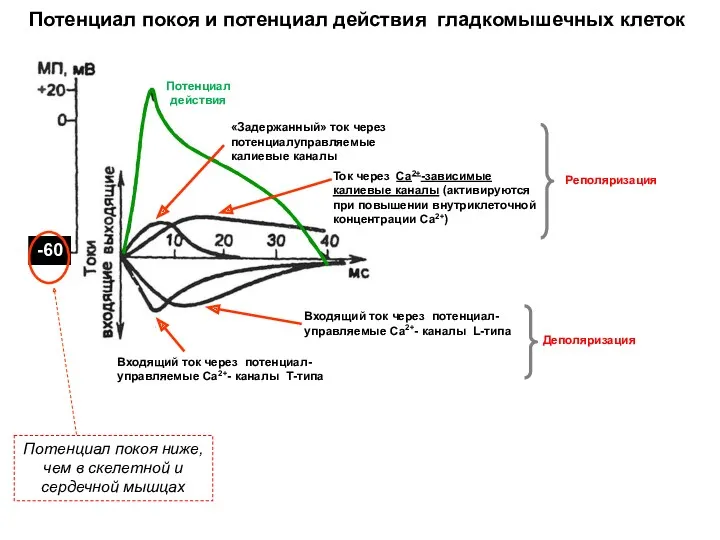
Потенциал покоя и потенциал действия гладкомышечных клеток
Потенциал покоя ниже,
чем в
Потенциал покоя и потенциал действия гладкомышечных клеток
Потенциал покоя ниже, чем в
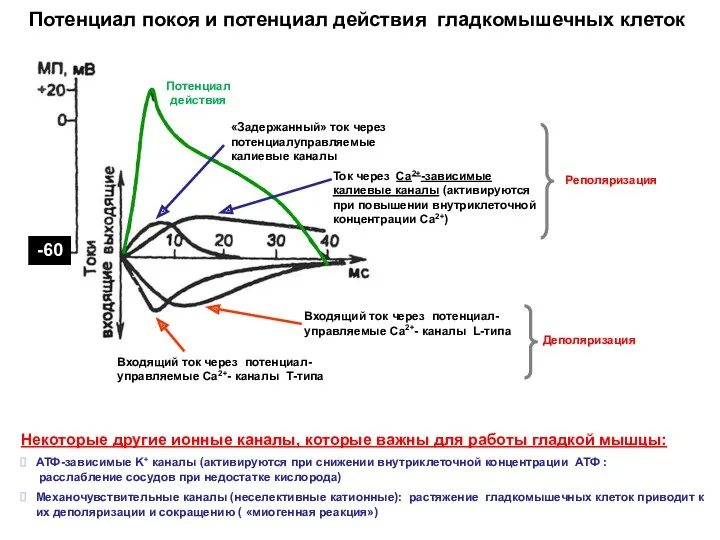
Потенциал покоя и потенциал действия гладкомышечных клеток
Некоторые другие ионные каналы, которые
Потенциал покоя и потенциал действия гладкомышечных клеток
Некоторые другие ионные каналы, которые
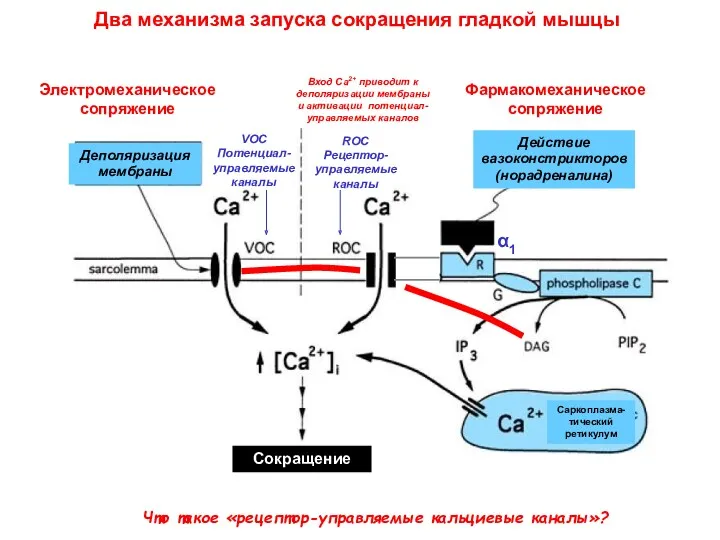
Два механизма запуска сокращения гладкой мышцы
Что такое «рецептор-управляемые кальциевые каналы»?
Вход Са2+
Два механизма запуска сокращения гладкой мышцы
Что такое «рецептор-управляемые кальциевые каналы»?
Вход Са2+
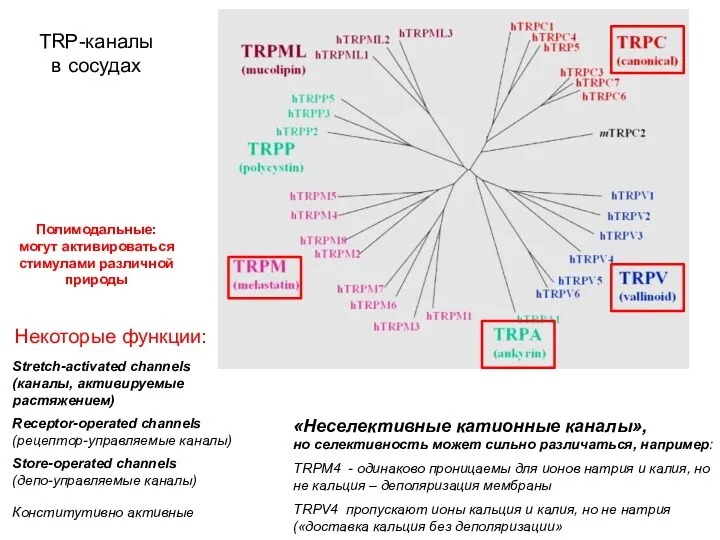
TRP-каналы
в сосудах
Store-operated channels
(депо-управляемые каналы)
Receptor-operated channels
(рецептор-управляемые каналы)
Stretch-activated channels
(каналы,
TRP-каналы
в сосудах
Store-operated channels
(депо-управляемые каналы)
Receptor-operated channels
(рецептор-управляемые каналы)
Stretch-activated channels (каналы,
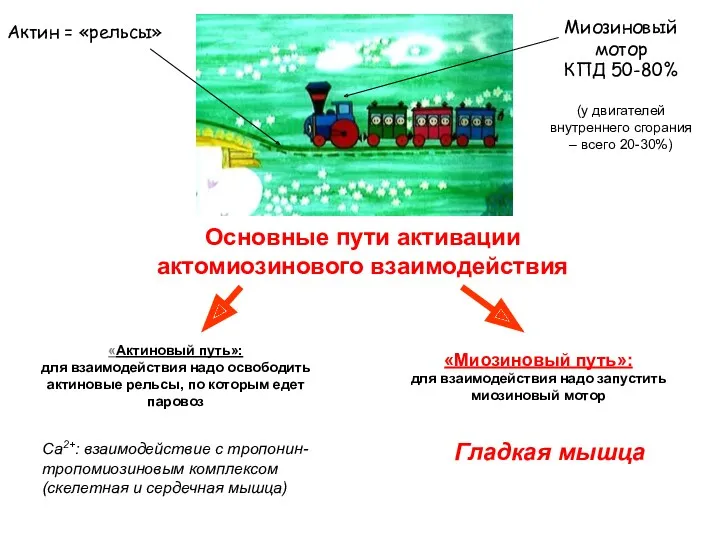
Основные пути активации актомиозинового взаимодействия
Актин = «рельсы»
Миозиновый мотор
КПД 50-80%
(у двигателей
Основные пути активации актомиозинового взаимодействия
Актин = «рельсы»
Миозиновый мотор
КПД 50-80%
(у двигателей
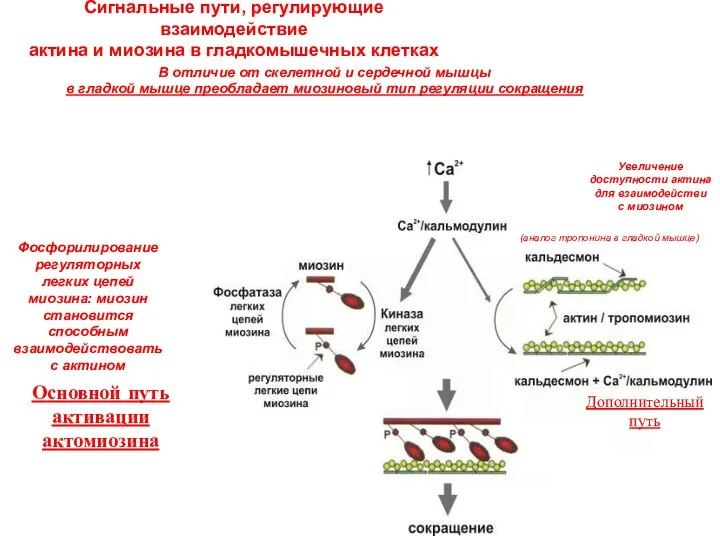
Сигнальные пути, регулирующие взаимодействие
актина и миозина в гладкомышечных клетках
В отличие
Сигнальные пути, регулирующие взаимодействие
актина и миозина в гладкомышечных клетках
В отличие
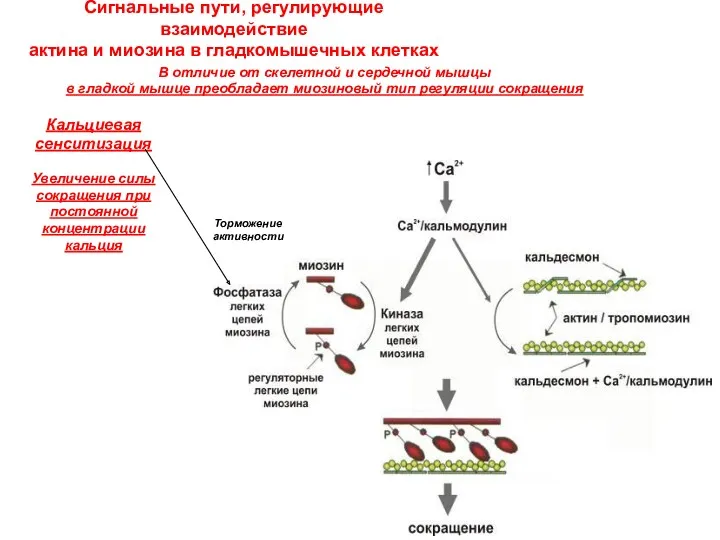
Сигнальные пути, регулирующие взаимодействие
актина и миозина в гладкомышечных клетках
В отличие
Сигнальные пути, регулирующие взаимодействие
актина и миозина в гладкомышечных клетках
В отличие
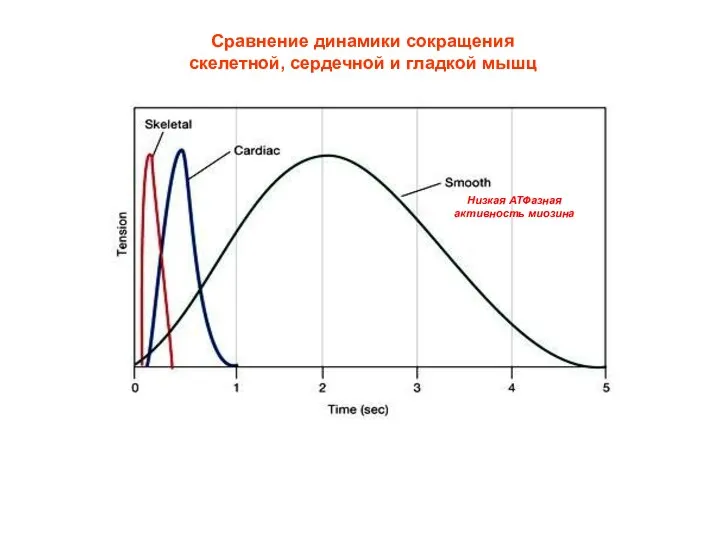
Сравнение динамики сокращения скелетной, сердечной и гладкой мышц
Низкая АТФазная активность
Сравнение динамики сокращения скелетной, сердечной и гладкой мышц
Низкая АТФазная активность
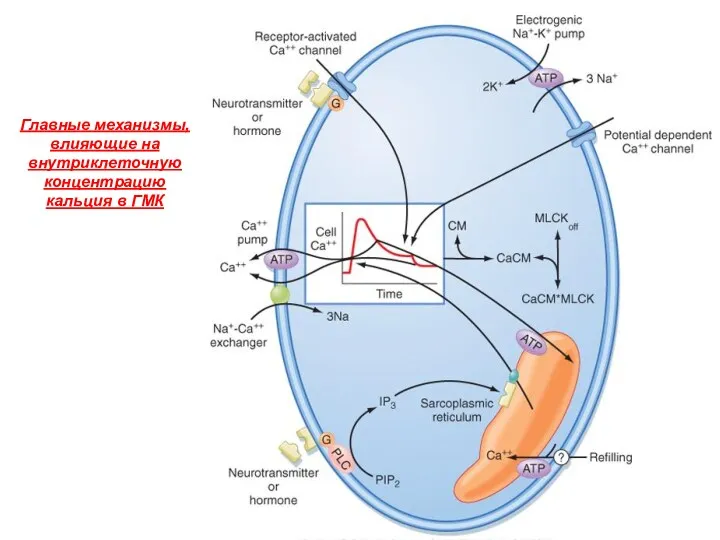
Главные механизмы, влияющие на внутриклеточную концентрацию кальция в ГМК
Главные механизмы, влияющие на внутриклеточную концентрацию кальция в ГМК

 Лимфатическая система
Лимфатическая система The requirement of growing tomatoes in the greenhouse
The requirement of growing tomatoes in the greenhouse Экологические сообщества. Экосистема, биогеоценоз
Экологические сообщества. Экосистема, биогеоценоз Взаимодействие генов
Взаимодействие генов Презентация Как растения показывают погоду и время.
Презентация Как растения показывают погоду и время. Разнообразие цветов. Строение цветка
Разнообразие цветов. Строение цветка Животные жарких стран
Животные жарких стран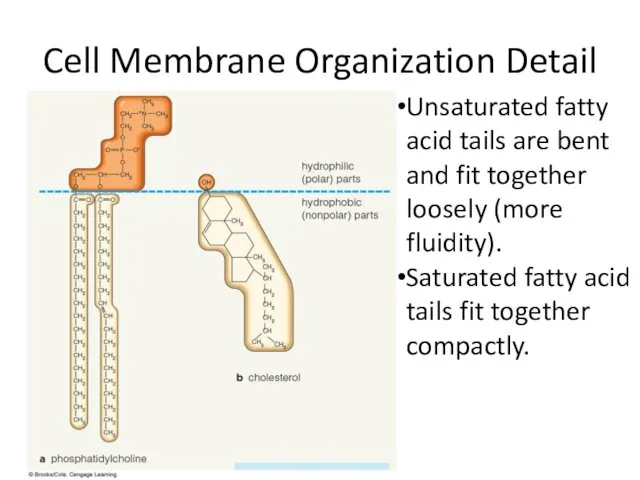 Cell Membrane Organization Detail
Cell Membrane Organization Detail ТН ВЭД: Группа 08. Съедобные фрукты и орехи; кожура цитрусовых плодов или корки дынь
ТН ВЭД: Группа 08. Съедобные фрукты и орехи; кожура цитрусовых плодов или корки дынь Дикие животные
Дикие животные Взгляды, гипотезы и теории о происхождении жизни
Взгляды, гипотезы и теории о происхождении жизни Кактусы. Об этих интереснейших растениях
Кактусы. Об этих интереснейших растениях Влияние шума на организм человека
Влияние шума на организм человека Пентозо-фосфатный путь окисления глюкозы
Пентозо-фосфатный путь окисления глюкозы Эксперимент Шпемана – путь к клонированию
Эксперимент Шпемана – путь к клонированию Биотехнология растений. Культуры растительных клеток и тканей in vitro
Биотехнология растений. Культуры растительных клеток и тканей in vitro Mitosis. Asexual reproduction
Mitosis. Asexual reproduction Мы из Красной книги. Викторина
Мы из Красной книги. Викторина Современные представления об эволюции орган мира(9 класс)
Современные представления об эволюции орган мира(9 класс) Влияние экстрасола на всхожесть и биометрические параметры проростков кукурузы
Влияние экстрасола на всхожесть и биометрические параметры проростков кукурузы Эндемик өсімдіктер: Кауфман қызғалдағы
Эндемик өсімдіктер: Кауфман қызғалдағы Разнообразие грибов: сапрофиты, фитопатогенные, зоопатогенные
Разнообразие грибов: сапрофиты, фитопатогенные, зоопатогенные Зимующие птицы
Зимующие птицы Влажность как экологический фактор
Влажность как экологический фактор Презентация к уроку биологии 6 кл.
Презентация к уроку биологии 6 кл. Химический круговорот веществ в природе. 6 класс
Химический круговорот веществ в природе. 6 класс Биологиялық белсенді затттардың сапалық құрамы мен сандық мөлшеріне әсер ететін факторлар
Биологиялық белсенді затттардың сапалық құрамы мен сандық мөлшеріне әсер ететін факторлар Происхождение жизни на Земле
Происхождение жизни на Земле