- Основы молекулярной биологии

Содержание
- 2. План лекции Основные понятия и определения молекулярной биологии Строение ДНК и РНК Биосинтез белка
- 3. Молекулярная биология Термин ввел Фрэнсис Крик Это раздел биологии который раскрывает организацию и структуру молекулярных механизмов
- 4. 1869 г. – открыты нуклеиновые кислоты Иоганн Фридрих Мишер Открыл ДНК. Вначале новое вещество получило название
- 5. Нуклеиновая кислота В 1889 г. Рихард Альтман разделил нуклеин на нуклеиновую кислоту и белок и предложил
- 6. 1949г. Правила Чаргаффа (закон «эквивалентности» оснований) Эрвин Чаргафф показал, что ДНК из разных биологических источников содержит
- 7. 1953 г. открыта модель молекулы ДНК Фрэнсис Крик и Джеймс Уотсон (лауреаты Нобелевской премии) Первая демонстрационная
- 8. Физики рентгено-структурщики – Розалинда Франклин и Морис Уилкинс получили первые рентгенограммы ДНК (1950-1953гг.) Розалинда Франклин Морис
- 9. 1961 год - структура генетического кода Дж. Генрих Маттеи Маршалл Ниренберг
- 10. 1965 год - установление факта регуляции бактериального лактозного оперона у E. coli Андре Львов ЖАКОБ ФРАНСУА
- 11. 1967 год - синтез биологически-активной ДНК в системе in vitro 1968 год - впервые осуществлен химический
- 12. 1973 г. – разработан метод секвенирование Фредерик Сенгер (дважды лауреат Нобелевской премии) Секвенирование – метод установления
- 13. 1985 г. открыт метод полимеразной цепной реакции (ПЦР) Кэри Муллис (лауреат Нобелевской премии)
- 14. Современные представления Устройство генетического материала у высших организмов принципиально отличается от низших . Основные различия: 1.
- 15. 2003 г. завершен проект «Геном человека» Примеры распределения генов человека по их функциям Френсис Коллинз Количество
- 16. Нуклеотиды – органические вещества, состоящие из гетероциклического азотистого основания, моносахарида (пентозы) и остатка фосфорной кислоты. Азотистые
- 17. Полные и сокращенные обозначения нуклеозидов Названия нуклеотидов (рибо- и дезоксирибонуклеотидов) складывается из названия соответствующего нуклеозида и
- 18. Нуклеиновые кислоты являются биологическими полимерами. Существует два типа нуклеиновых кислот: ДНК (дезоксирибонуклеиновая кислота) и РНК (рибонуклеиновая
- 19. СТРУКТУРА ДНК последовательность нуклеотидов в 2-х полинуклеотидных цепях, соединенных между собой фосфодиэфирными связями в 3'-5'- направлении
- 20. Двойная спираль ДНК расположиться так, что против каждого нуклеотида одной окажется селективно взаимодействующий с ней другой
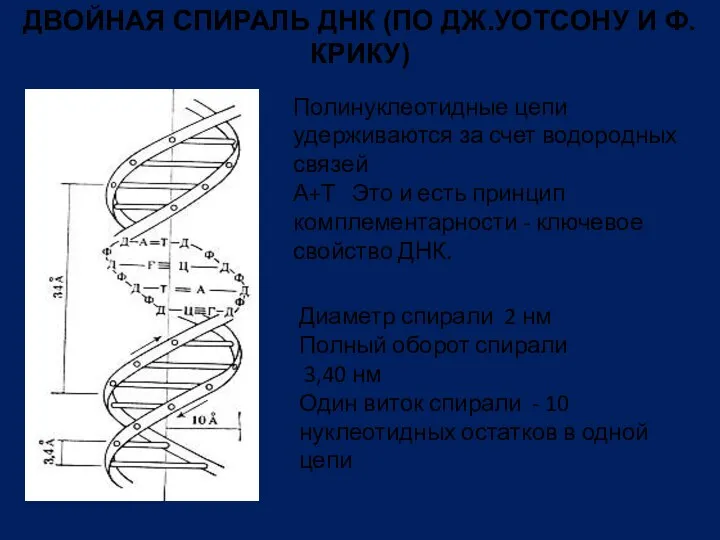
- 21. ДВОЙНАЯ СПИРАЛЬ ДНК (ПО ДЖ.УОТСОНУ И Ф. КРИКУ) Полинуклеотидные цепи удерживаются за счет водородных связей А+Т
- 22. Формы ДНК А-ДНК В-ДНК Z-ДНК Молекулы ДНК могут находятся в различных конформационных состояниях, зависящих от степени
- 23. В А-форме ДНК выполняет роль матрицы в процессе транскрипции (синтез РНК на молекуле ДНК). В В-форме
- 24. – это структура хроматина ядра и хромосом. Нуклеосома – элементарная единица структуры хроматина, компактная нуклеопротеидная частица,
- 25. Суперспирализация ДНК формирует третичную структуру с помощью разнообразных белков - гистоновых и негистоновых. Гистоны - Н2А,
- 26. Электростатическое взаимодействие При участии структурных, регуляторных белков и ферментов, участвующих в синтезе ДНК и РНК, нить
- 27. ФУНКЦИИ ДНК 1. Аутосинтетическая – способность молекулы ДНК к репликации, самоудвоение ДНК в S-период интерфазы 2.
- 28. Регуляция генетической активности ДНК Гистоны подавляют генетическую функцию ДНК. Нуклеосомы – это фрагменты «молчащего» хроматина, а
- 29. Структура РНК напоминает ДНК, отличие: в основной цепи фрагменты фосфорной кислоты чередуются с рибозой, а не
- 30. Химические свойства РНК Напоминают свойства ДНК, однако наличие дополнительных групп ОН в рибозе и меньшее (в
- 31. В цитоплазме клеток присутствуют три типа РНК транспортные РНК (тРНК), матричные РНК (мРНК) и рибосомальные РНК
- 32. Первичная структура РНК – порядок чередования рибонуклеозидмонофосфатов в полинуклеотидной цепи. Нуклеотиды в РНК связаны между собой
- 33. Транспортные РНК Пространственная структура тРНК описывается универсальной моделью «клеверного листа». Последовательность тРНК включают 70-90 нуклеотидов. Структура
- 34. Транспортные РНК Г-образная структура состоит из двух спиралей расположенных почти перпендикулярно одна относительно другой. Одну спираль
- 35. т-РНК На долю тРНК приходится около 10% от общего содержания РНК в клетке. Функции тРНК: 1)
- 36. Матричные Информационные РНК разнообразны по содержанию нуклеотидов и молекулярной массе (от 50 000 до 4 000
- 37. Рибосомные РНК содержат 3000–5000 нуклеотидов; молекулярная масса — 1 000 000–1 500 000. На долю рРНК
- 38. Рибосомальные РНК Клетки прокариот и эукариот содержат рибосомы, имеющие общий план строения. В рибосомы входят высокомолекулярные
- 39. Рибосомальные РНК Они принимают участие в образовании рибонуклеопротеинов, формирующих немембранные комплексы – рибосомы
- 40. БИОСИНТЕЗ НУКЛЕИНОВЫХ КИСЛОТ И БЕЛКОВ (МАТРИЧНЫЕ БИОСИНТЕЗЫ) При биосинтезе новых молекул нуклеиновых кислот и белков носителями
- 41. Генетическая информация программирует синтез специфических белков, определяющих в свою очередь специфичность структуры и функции клеток, органов
- 42. Биосинтез ДНК 1. Одноцепочечная – неспаренная ДНК 2. Дезоксирибонуклеозидтрифосфаты 3. Ферменты – ДНК-полимеразы, топоизомеразы (гиразы), хеликазы,
- 43. Репликация ДНК Двойная цепь ДНК служит матрицей для 2-х дочерних ДНК Синтез происходит комплементарно Три механизма
- 44. 1958г. М. Мезельсон и Ф. Сталь: реплекация ДНК у эукариотов осуществляется по полуконсервативному механизму: двойная спираль
- 45. 1963г. Ф. Жакоб, С. Бреннер, Ф. Кьюзен: Репликон – генетическая единица репликации, то есть сегмент ДНК,
- 46. Для синтеза ДНК необходимы: Матрица ДНК (I цепочечная) Субстраты - четыре dНТФ: dАТФ, dГТФ, dЦТФ, dТТФ;
- 47. Матричная цепь ДНК считывается в направлении 3'→5', вновь образующаяся цепь синтезируется антипараллельно матрице - от 5'-конца
- 48. Принцип комплементарности является ведущим не только в построении биомолекул, но и в передаче информации в живых
- 49. Репликацию можно разделить на 3 этапа: инициация (начало) – образование репликативной вилки. элонгация (продолжение) – синтез
- 50. В клетках эукариот имеются пять различных ДНК-зависимых ДНК – полимераз: α, β, δ, ε, γ они
- 51. Инициация - создание репликативной вилки Хеликаза - расплетение родительской ДНК, на разделение каждой пары оснований затрачивается
- 52. Образуется репликативный «глаз» - две репликативных «вилки», каждая имеет протяжённость до 2000 неспаренных оснований.
- 53. Каждая из разделенных цепей связывается с ДНК- связывающими белками (SSB), которые: Стабилизируют одноцепочечные участки ДНК, благодаря
- 54. ДНК-зависимая РНК - роl (праймаза) комплекса ДНК-полимераза α - праймаза синтезирует РНК - «затравки» размером в
- 55. ДНК человека: Содержит 150 млн пар нуклеотидов Скорость синтеза - 50 нуклеотидов в 1 мин. Инициация
- 56. Начало репликации происходит на специфических репликаторах - в точках ori, которые у млекопитающих отстоят на расстоянии
- 57. Ориджины (от англ. origin- происхождение) - сайты инициации репликации Репликон – единица репликации – последовательность ДНК,
- 58. Факторами, влияющими на клеточное деление, являются: а) факторы роста (пептиды, активирующие деление определённого типа клеток); б)
- 59. Известно 100 факторов роста и около 40 факторов, подавляющих клеточное деление Механизмы действия факторов роста на
- 60. ЭЛОНГАЦИЯ а) Синтез лидирующей цепи осуществляет ДНК-полимераза δ, а отстающей ДНК-полимераза ε. Они удлиняют «затравку», присоединяя
- 61. Репликация у эукариот идет на уровне нуклеосом
- 62. Элонгация в) 5'-3' экзонуклеаза и РНК-аза Н1 удаляют РНК-затравки (праймер). г) ДНК-полимераза заполняет бреши. д) ДНК-лигаза
- 63. Потери генетической информации не происходит: укорочение идет за счет теломер –многократно повторяющейся на каждом конце хромосомы
- 64. Механизм действия теломеразы: а) с помощью РНК фермент комплементарно прикрепляется к 5' – концу недостроенной дочерней
- 65. Наибольшая активность теломеразы обнаружена в клетках с высокой скоростью обновления: Лимфоциты Стволовые клетки костного мозга Клетки
- 66. В соматических клетках теломераза неактивна Соматическая клетка имеет длину теломерной ДНК, достаточную для времени жизни клетки
- 67. Терминация репликации Продвижение репликативной вилки прекращается: а) при столкновении с другой вилкой, движущейся в противоположном направлении,
- 68. Точность репликации ДНК 1 ошибка на 10 трансферазных реакций, однако подобная ошибка обычно легко исправляется за
- 69. Открытие обратной транскриптазы указывает на возможность передачи наследственной информации от РНК на ДНК, не подчиняясь основному
- 70. Экспрессия генов Процесс транскрипции – биосинтез РНК на матрице ДНК. Начальная стадия реализации генетической информации. В
- 71. передача (переписывание) информации, содержащейся в последовательности нуклеотидных остатков в ДНК-матрице, в последовательность нуклеиновых звеньев в молекуле
- 72. Транскриптон – часть молекулы ДНК транскрибируется в процессе биосинтеза РНК на ней. ограниченная промотором (точка начала
- 73. Независимая транскрипция гена – ген обладает собственным промотором и терминатором транскрипции. Координированная транскрипция генов – группа
- 74. Ген эукариот является мозаичной структурой, содержащей наряду с кодирующими (экзонами) также некодирующие (интроны) последовательности Экзоны (от
- 75. Соотношение информативной и неинформативной частей в транскриптонах Эукариотов – 1:9 Прокариотов – 9:1
- 76. Соседние транскриптоны могут быть отделены друг от друга нетранскрибируемыми участками Биологический смысл: разделение ДНК на множество
- 77. Ферменты транскрипции В 1960 г. открыт в двух лабораториях США (Дж. Хервиц и С. Вейс) ДНК
- 78. Фермент РНК-полимераза катализирует транскрипцию как экзонов, так и интронов Для синтеза РНК, катализируемого РНК - pol
- 79. Кор-фермент без субъединицы σ, начинает работать только после ее присоединения, узнает промоторные сайты, с которых начинается
- 80. Транскрипция Инициация Элонгация Терминация -35 -10 +1 ----TTGACA ----------------TATAAT-------------CAT----- ----AACTGT----------------ATATTA-------------GTA---- 16-19 bp 4-8 bp Промотор –
- 81. Факторы транскрипции регуляторные белки обеспечивающие у эукариот работу РНК-pol ТАТА- связывающий белок (ТСБ, англ. ТАТА-Вох Binding
- 82. РНК-pol связывается с промотором, происходит локальное раскручивание двойной спирали ДНК (примерно 18 п.н.) и образование открытого
- 83. Копирование смысловой (ДНК+) цепи Направление 5’-3’ Первый нуклеотид всегда АТФ или ГТФ, затем образуется первая фосфодиэфирная
- 84. Факторы элонгации (Е,Н,F) повышают активность РНК-pol и облегчают расхождение цепей. Скорость работы РНК-pol 50 нуклеотидов в
- 85. Завершение синтеза РНК Происходит в сайтах терминации (транскрипционные терминаторы – GC – богатые участки) – строго
- 86. Факторы транскрипции объединены вместе с РНК-pol в единый транскрипционный комплекс.
- 87. Транскрипция сходна с репликацей В основе механизма процесса лежит принцип комплементарности (А=У, Г=Ц) и антипараллельности: синтез
- 88. Транскрипция отличается от репликации рядом особенностей: 1. Не требует синтеза праймера. 2. Использует не всю молекулу
- 89. Регуляторные элементы, стимулирующие транскрипцию связанных с ними генов, называются энхансерами (усилителями, от англ. enhancer). подавляющие транскрипцию
- 90. Транскрипционные факторы (Транс- действующие факторы) это белки, т. е. продукты других, независимых генов, поэтому их называют
- 91. Процессинг первичных транскриптов РНК Между транскрипцией и трансляцией существует важное звено – процессинг или созревание мРНК.
- 92. Схема сплайсинга РНК Катализируется протеиновым комплексом, сплайсосомой Состав: белки, мяРНК (рибозимы – катализируют расщепление фосфодиэфирных связей),
- 93. Регуляция генной экспрессии Не все белки синтезируются постоянно Некоторые белки синтезируются только при определенных условиях Синтезируемые
- 94. Гипотеза оперона (1961г. Ф. Жакоб и Ж. Моно) ген-регулятор промотор – точки инициации для синтеза мРНК
- 95. Организация хроматина в дифференцированных клетках многоклеточного организма Гетерохроматин (стабильно репрессированные участки)- ДНК упакована компактно и недоступна
- 96. В разных типах клеток в область эухроматина попадают разные гены, а это означает, что в разных
- 97. Регуляторные зоны ДНК Энхансеры и сайленсоры различны по числу и расположению на молекуле ДНК для различных
- 98. Это способ записи информации об аминоксилотной последовательности белков с помощью последовательности нуклеотидов в мРНК Генетический код
- 99. СВОЙСТВА ГЕНЕТИЧЕСКОГО КОДА Триплетность - кодовое число = 3. Три нуклеотидных остатка (триплет) кодируют одну аминокислоту.
- 100. Код непрерывен, отсутствие «знаков препинания», т.е. сигналов, указывающих на конец одного кодона и начало другого. Другими
- 101. Вторичная структура тРНК Третичная структура тРНК
- 102. Значение активирования АК: аатРНК - это транспортная форма АК Каждая АК получила свой шифр в виде
- 103. Рост белковой цепи идет с N- конца, а С-конец – закреплен на рибосоме
- 104. Полисома – комплекс, состоящий из рибосом, расположенных на мРНК с интервалом 100 нуклеотидов
- 105. Функции субъединиц рибосомы в процессе трансляции: Малая субъединица присоединяет мРНК и декодирует информацию с помощью тРНК
- 107. Скачать презентацию

План лекции
Основные понятия и определения молекулярной биологии
Строение ДНК и РНК
Биосинтез белка
План лекции
Основные понятия и определения молекулярной биологии
Строение ДНК и РНК
Биосинтез белка

Молекулярная биология
Термин ввел Фрэнсис Крик
Это раздел биологии который раскрывает организацию и
Молекулярная биология
Термин ввел Фрэнсис Крик
Это раздел биологии который раскрывает организацию и

1869 г. – открыты нуклеиновые кислоты
Иоганн Фридрих
Мишер
Открыл ДНК. Вначале
1869 г. – открыты нуклеиновые кислоты
Иоганн Фридрих
Мишер
Открыл ДНК. Вначале

Нуклеиновая кислота
В 1889 г. Рихард Альтман разделил нуклеин на нуклеиновую
Нуклеиновая кислота
В 1889 г. Рихард Альтман разделил нуклеин на нуклеиновую

1949г. Правила Чаргаффа (закон «эквивалентности» оснований)
Эрвин Чаргафф показал, что ДНК из
1949г. Правила Чаргаффа (закон «эквивалентности» оснований)
Эрвин Чаргафф показал, что ДНК из

1953 г. открыта модель молекулы ДНК
Фрэнсис Крик и Джеймс Уотсон
1953 г. открыта модель молекулы ДНК
Фрэнсис Крик и Джеймс Уотсон

Физики рентгено-структурщики – Розалинда Франклин и Морис Уилкинс получили первые рентгенограммы
Физики рентгено-структурщики – Розалинда Франклин и Морис Уилкинс получили первые рентгенограммы

1961 год - структура генетического кода
Дж. Генрих Маттеи
Маршалл Ниренберг
1961 год - структура генетического кода
Дж. Генрих Маттеи
Маршалл Ниренберг

1965 год - установление факта регуляции бактериального лактозного оперона у E.
1965 год - установление факта регуляции бактериального лактозного оперона у E.

1967 год - синтез биологически-активной ДНК в системе in vitro
1968 год
1967 год - синтез биологически-активной ДНК в системе in vitro
1968 год
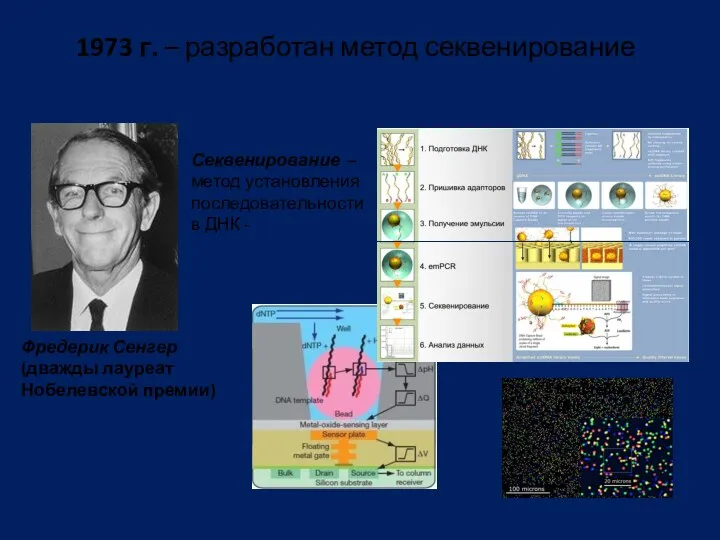
1973 г. – разработан метод секвенирование
Фредерик Сенгер
(дважды лауреат
Нобелевской
1973 г. – разработан метод секвенирование
Фредерик Сенгер
(дважды лауреат
Нобелевской

1985 г. открыт метод полимеразной цепной реакции (ПЦР)
Кэри Муллис
(лауреат Нобелевской
1985 г. открыт метод полимеразной цепной реакции (ПЦР)
Кэри Муллис
(лауреат Нобелевской

Современные представления
Устройство генетического материала у высших организмов принципиально отличается от
Современные представления
Устройство генетического материала у высших организмов принципиально отличается от

2003 г. завершен проект «Геном человека»
Примеры распределения генов человека
2003 г. завершен проект «Геном человека»
Примеры распределения генов человека
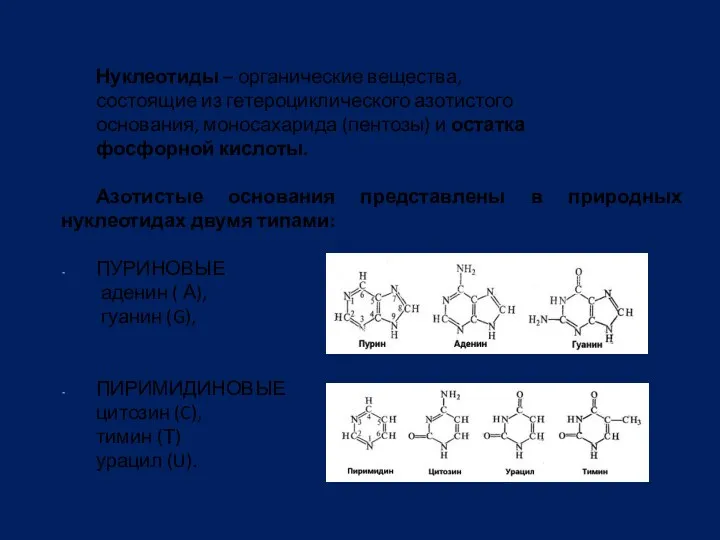
Нуклеотиды – органические вещества,
состоящие из гетероциклического азотистого
основания, моносахарида (пентозы)
Нуклеотиды – органические вещества,
состоящие из гетероциклического азотистого
основания, моносахарида (пентозы)
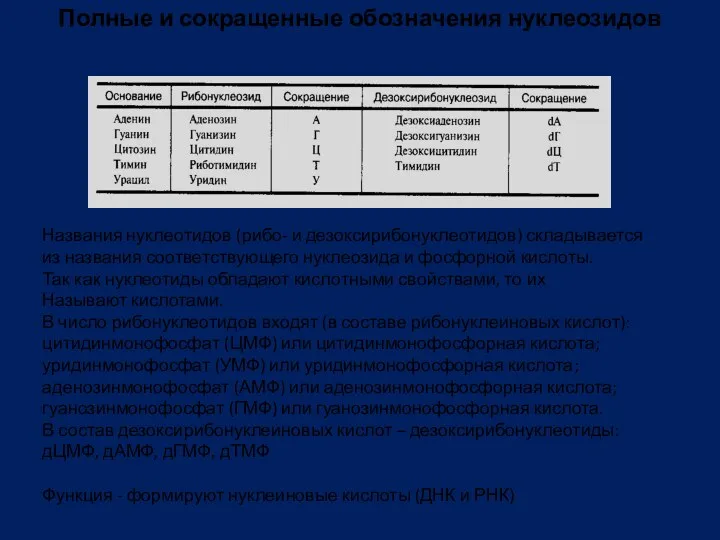
Полные и сокращенные обозначения нуклеозидов
Названия нуклеотидов (рибо- и дезоксирибонуклеотидов) складывается
из
Полные и сокращенные обозначения нуклеозидов
Названия нуклеотидов (рибо- и дезоксирибонуклеотидов) складывается
из

Нуклеиновые кислоты являются биологическими полимерами.
Существует два типа нуклеиновых кислот:
Нуклеиновые кислоты являются биологическими полимерами.
Существует два типа нуклеиновых кислот:
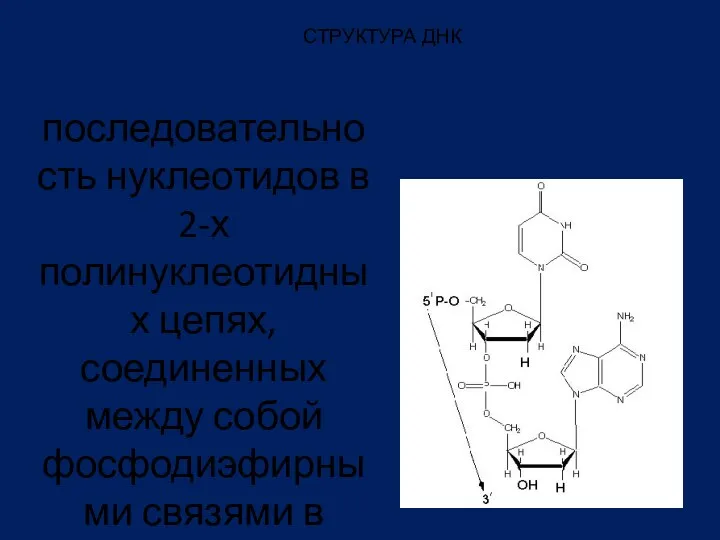
СТРУКТУРА ДНК
последовательность нуклеотидов в 2-х полинуклеотидных цепях, соединенных между собой фосфодиэфирными
СТРУКТУРА ДНК
последовательность нуклеотидов в 2-х полинуклеотидных цепях, соединенных между собой фосфодиэфирными

Двойная спираль ДНК
расположиться так, что против каждого нуклеотида одной окажется селективно
Двойная спираль ДНК
расположиться так, что против каждого нуклеотида одной окажется селективно
ДВОЙНАЯ СПИРАЛЬ ДНК (ПО ДЖ.УОТСОНУ И Ф. КРИКУ)
Полинуклеотидные цепи удерживаются
ДВОЙНАЯ СПИРАЛЬ ДНК (ПО ДЖ.УОТСОНУ И Ф. КРИКУ)
Полинуклеотидные цепи удерживаются

Формы ДНК
А-ДНК
В-ДНК
Z-ДНК
Молекулы ДНК могут находятся в различных конформационных состояниях, зависящих от
Формы ДНК
А-ДНК
В-ДНК
Z-ДНК
Молекулы ДНК могут находятся в различных конформационных состояниях, зависящих от

В А-форме ДНК выполняет роль матрицы в процессе транскрипции (синтез РНК

– это структура хроматина ядра и хромосом.
Нуклеосома – элементарная единица
– это структура хроматина ядра и хромосом.
Нуклеосома – элементарная единица

Суперспирализация ДНК формирует третичную структуру с помощью
разнообразных белков - гистоновых
Суперспирализация ДНК формирует третичную структуру с помощью
разнообразных белков - гистоновых
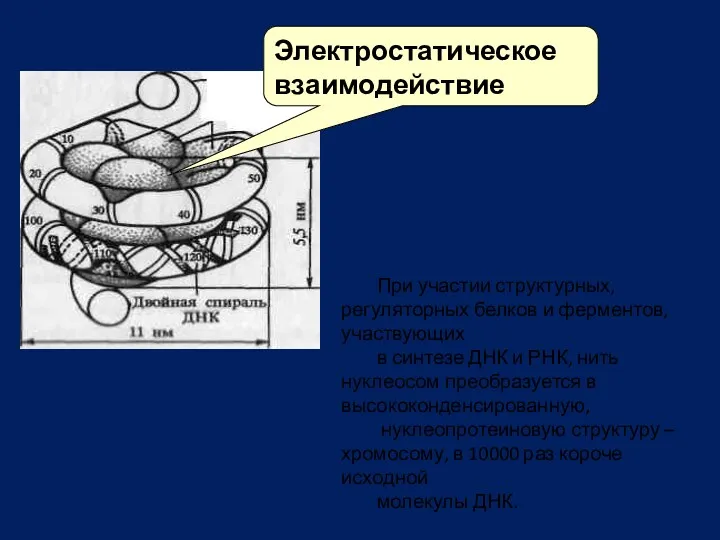
Электростатическое взаимодействие
При участии структурных, регуляторных белков и ферментов, участвующих
в синтезе
Электростатическое взаимодействие
При участии структурных, регуляторных белков и ферментов, участвующих
в синтезе
ФУНКЦИИ ДНК
1. Аутосинтетическая – способность молекулы ДНК к репликации, самоудвоение ДНК
ФУНКЦИИ ДНК
1. Аутосинтетическая – способность молекулы ДНК к репликации, самоудвоение ДНК
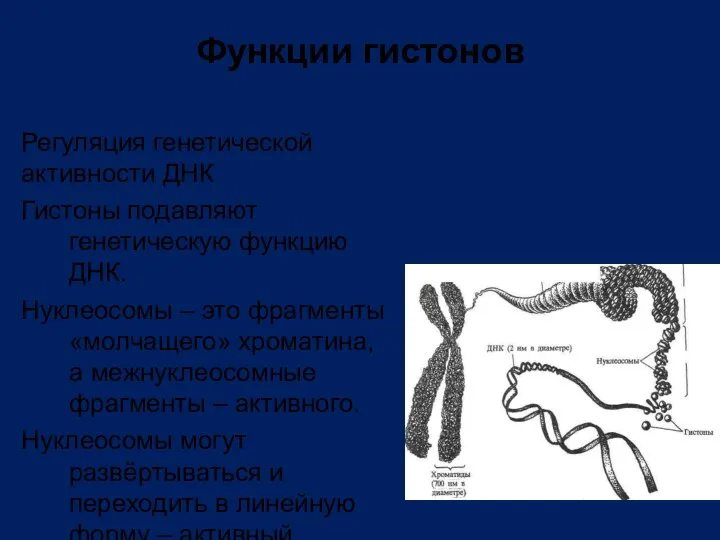
Регуляция генетической активности ДНК
Гистоны подавляют генетическую функцию ДНК.
Нуклеосомы – это
Регуляция генетической активности ДНК
Гистоны подавляют генетическую функцию ДНК.
Нуклеосомы – это
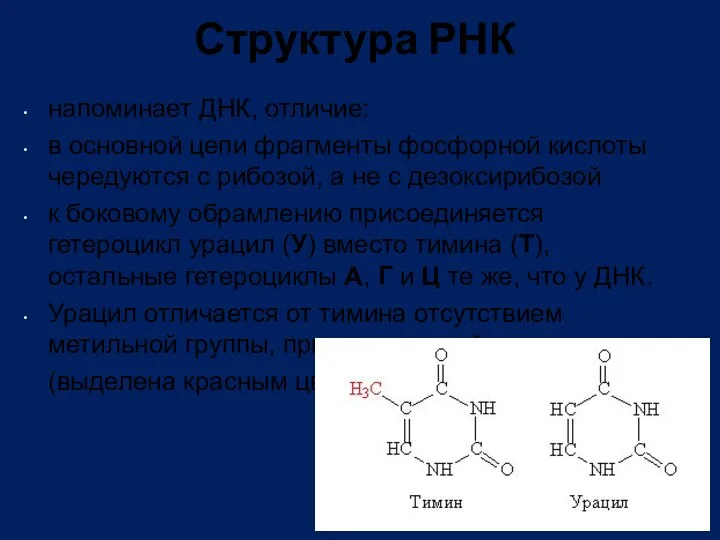
Структура РНК
напоминает ДНК, отличие:
в основной цепи фрагменты фосфорной кислоты чередуются с
Структура РНК
напоминает ДНК, отличие:
в основной цепи фрагменты фосфорной кислоты чередуются с

Химические свойства РНК
Напоминают свойства ДНК, однако наличие дополнительных групп ОН в
Химические свойства РНК
Напоминают свойства ДНК, однако наличие дополнительных групп ОН в

В цитоплазме клеток присутствуют три типа РНК
транспортные РНК (тРНК), матричные
В цитоплазме клеток присутствуют три типа РНК
транспортные РНК (тРНК), матричные

Первичная структура РНК – порядок чередования рибонуклеозидмонофосфатов в полинуклеотидной цепи.
Нуклеотиды
Первичная структура РНК – порядок чередования рибонуклеозидмонофосфатов в полинуклеотидной цепи.
Нуклеотиды

Транспортные РНК
Пространственная структура тРНК описывается универсальной моделью «клеверного листа».
Последовательность тРНК
Транспортные РНК
Пространственная структура тРНК описывается универсальной моделью «клеверного листа».
Последовательность тРНК

Транспортные РНК
Г-образная структура состоит из двух спиралей расположенных почти перпендикулярно одна
Транспортные РНК
Г-образная структура состоит из двух спиралей расположенных почти перпендикулярно одна

т-РНК
На долю тРНК приходится около 10% от общего содержания РНК
т-РНК
На долю тРНК приходится около 10% от общего содержания РНК

Матричные Информационные РНК разнообразны по содержанию нуклеотидов и молекулярной массе (от
Матричные Информационные РНК разнообразны по содержанию нуклеотидов и молекулярной массе (от

Рибосомные РНК содержат 3000–5000 нуклеотидов; молекулярная масса — 1 000 000–1 500 000.
На долю
Рибосомные РНК содержат 3000–5000 нуклеотидов; молекулярная масса — 1 000 000–1 500 000.
На долю

Рибосомальные РНК
Клетки прокариот и эукариот содержат рибосомы, имеющие общий план строения.
Рибосомальные РНК
Клетки прокариот и эукариот содержат рибосомы, имеющие общий план строения.
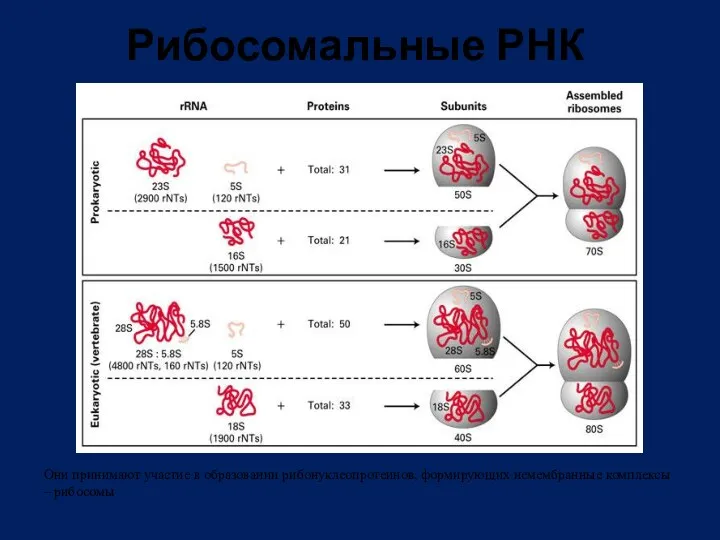
Рибосомальные РНК
Они принимают участие в образовании рибонуклеопротеинов, формирующих немембранные комплексы –
Рибосомальные РНК
Они принимают участие в образовании рибонуклеопротеинов, формирующих немембранные комплексы –

БИОСИНТЕЗ НУКЛЕИНОВЫХ КИСЛОТ И БЕЛКОВ (МАТРИЧНЫЕ БИОСИНТЕЗЫ)
При биосинтезе новых молекул нуклеиновых
БИОСИНТЕЗ НУКЛЕИНОВЫХ КИСЛОТ И БЕЛКОВ (МАТРИЧНЫЕ БИОСИНТЕЗЫ)
При биосинтезе новых молекул нуклеиновых

Генетическая информация программирует синтез специфических белков, определяющих в свою очередь специфичность
Генетическая информация программирует синтез специфических белков, определяющих в свою очередь специфичность

Биосинтез ДНК
1. Одноцепочечная – неспаренная ДНК
2. Дезоксирибонуклеозидтрифосфаты
3. Ферменты – ДНК-полимеразы, топоизомеразы
Биосинтез ДНК
1. Одноцепочечная – неспаренная ДНК
2. Дезоксирибонуклеозидтрифосфаты
3. Ферменты – ДНК-полимеразы, топоизомеразы

Репликация ДНК
Двойная цепь ДНК служит матрицей для 2-х дочерних ДНК
Синтез происходит
Репликация ДНК
Двойная цепь ДНК служит матрицей для 2-х дочерних ДНК
Синтез происходит

1958г. М. Мезельсон и Ф. Сталь:
реплекация ДНК у эукариотов осуществляется по
1958г. М. Мезельсон и Ф. Сталь: реплекация ДНК у эукариотов осуществляется по

1963г. Ф. Жакоб, С. Бреннер, Ф. Кьюзен:
Репликон – генетическая единица репликации,
1963г. Ф. Жакоб, С. Бреннер, Ф. Кьюзен: Репликон – генетическая единица репликации,

Для синтеза ДНК необходимы:
Матрица ДНК (I цепочечная)
Субстраты - четыре dНТФ:
Для синтеза ДНК необходимы:
Матрица ДНК (I цепочечная)
Субстраты - четыре dНТФ:
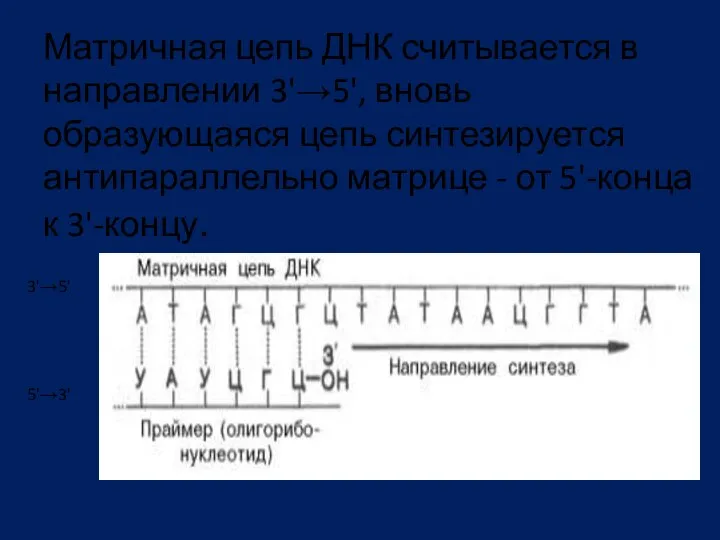
Матричная цепь ДНК считывается в направлении 3'→5', вновь образующаяся цепь синтезируется
Матричная цепь ДНК считывается в направлении 3'→5', вновь образующаяся цепь синтезируется
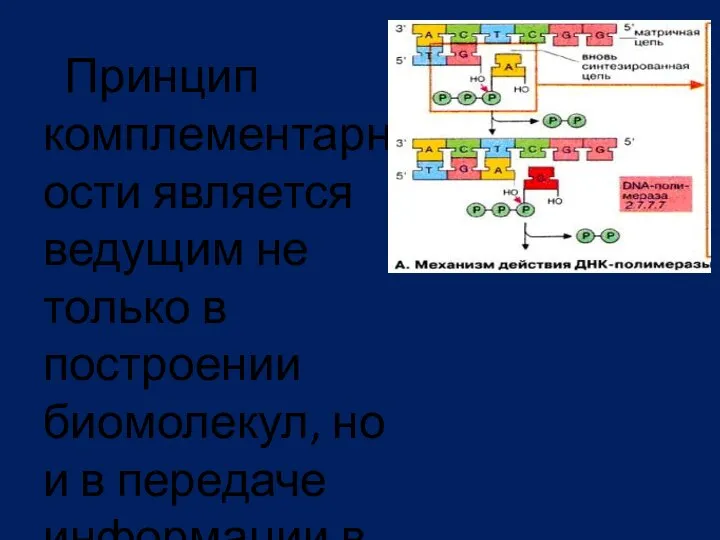
Принцип комплементарности является ведущим не только в построении биомолекул, но
Принцип комплементарности является ведущим не только в построении биомолекул, но

Репликацию можно разделить на 3 этапа:
инициация (начало) – образование репликативной вилки.
элонгация
Репликацию можно разделить на 3 этапа:
инициация (начало) – образование репликативной вилки.
элонгация

В клетках эукариот имеются пять различных ДНК-зависимых ДНК – полимераз: α,
В клетках эукариот имеются пять различных ДНК-зависимых ДНК – полимераз: α,

Инициация - создание репликативной вилки
Хеликаза - расплетение родительской ДНК, на разделение
Инициация - создание репликативной вилки
Хеликаза - расплетение родительской ДНК, на разделение

Образуется репликативный «глаз» - две репликативных «вилки», каждая имеет протяжённость до
Образуется репликативный «глаз» - две репликативных «вилки», каждая имеет протяжённость до

Каждая из разделенных цепей связывается с ДНК- связывающими белками (SSB), которые:
Стабилизируют
Каждая из разделенных цепей связывается с ДНК- связывающими белками (SSB), которые:
Стабилизируют

ДНК-зависимая РНК - роl (праймаза)
комплекса ДНК-полимераза α - праймаза синтезирует РНК
ДНК-зависимая РНК - роl (праймаза)
комплекса ДНК-полимераза α - праймаза синтезирует РНК

ДНК человека:
Содержит 150 млн пар нуклеотидов
Скорость синтеза - 50 нуклеотидов в
ДНК человека: Содержит 150 млн пар нуклеотидов Скорость синтеза - 50 нуклеотидов в
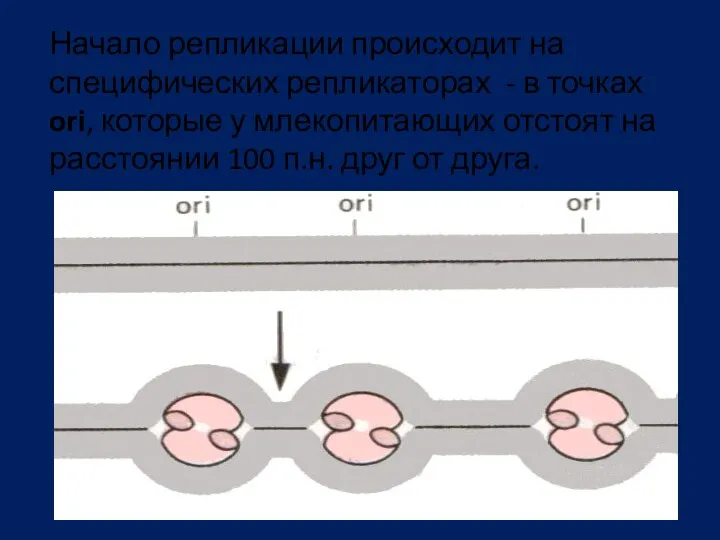
Начало репликации происходит на специфических репликаторах - в точках ori, которые
Начало репликации происходит на специфических репликаторах - в точках ori, которые
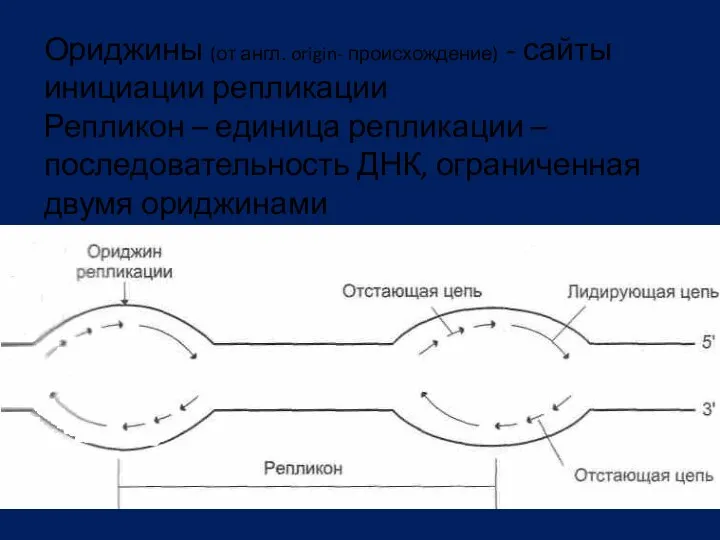
Ориджины (от англ. origin- происхождение) - сайты инициации репликации
Репликон –
Ориджины (от англ. origin- происхождение) - сайты инициации репликации Репликон –

Факторами, влияющими на клеточное деление, являются:
а) факторы роста (пептиды, активирующие деление
Факторами, влияющими на клеточное деление, являются:
а) факторы роста (пептиды, активирующие деление

Известно 100 факторов роста и около 40 факторов, подавляющих клеточное деление
Механизмы
Известно 100 факторов роста и около 40 факторов, подавляющих клеточное деление
Механизмы

ЭЛОНГАЦИЯ
а) Синтез лидирующей цепи осуществляет ДНК-полимераза δ, а отстающей ДНК-полимераза ε.
ЭЛОНГАЦИЯ
а) Синтез лидирующей цепи осуществляет ДНК-полимераза δ, а отстающей ДНК-полимераза ε.
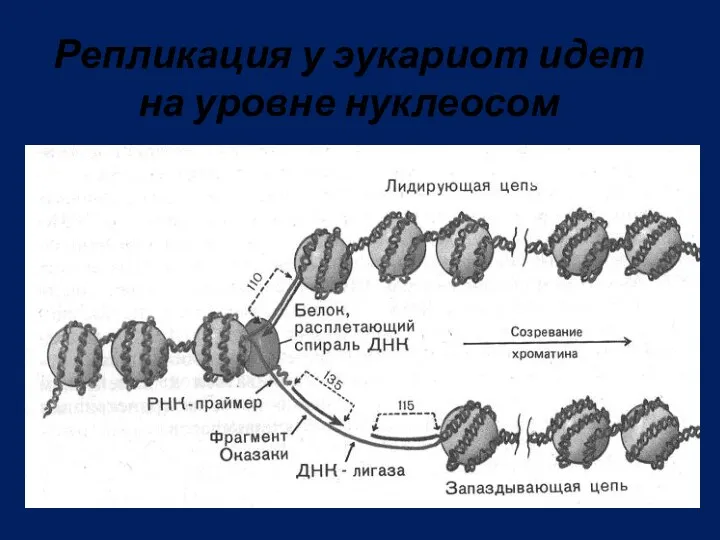
Репликация у эукариот идет на уровне нуклеосом
Репликация у эукариот идет на уровне нуклеосом

Элонгация
в) 5'-3' экзонуклеаза и РНК-аза Н1 удаляют РНК-затравки (праймер).
г) ДНК-полимераза заполняет
Элонгация
в) 5'-3' экзонуклеаза и РНК-аза Н1 удаляют РНК-затравки (праймер).
г) ДНК-полимераза заполняет

Потери генетической информации не происходит:
укорочение идет за счет теломер –многократно повторяющейся
Потери генетической информации не происходит:
укорочение идет за счет теломер –многократно повторяющейся

Механизм действия теломеразы:
а) с помощью РНК фермент комплементарно прикрепляется к
Механизм действия теломеразы:
а) с помощью РНК фермент комплементарно прикрепляется к

Наибольшая активность теломеразы обнаружена в клетках с высокой скоростью обновления:
Лимфоциты
Стволовые клетки
Наибольшая активность теломеразы обнаружена в клетках с высокой скоростью обновления:
Лимфоциты
Стволовые клетки

В соматических клетках теломераза неактивна
Соматическая клетка имеет длину теломерной ДНК, достаточную
В соматических клетках теломераза неактивна
Соматическая клетка имеет длину теломерной ДНК, достаточную

Терминация репликации
Продвижение репликативной вилки прекращается:
а) при столкновении с другой вилкой, движущейся
Терминация репликации
Продвижение репликативной вилки прекращается:
а) при столкновении с другой вилкой, движущейся

Точность репликации ДНК 1 ошибка на 10 трансферазных реакций, однако подобная
Точность репликации ДНК 1 ошибка на 10 трансферазных реакций, однако подобная
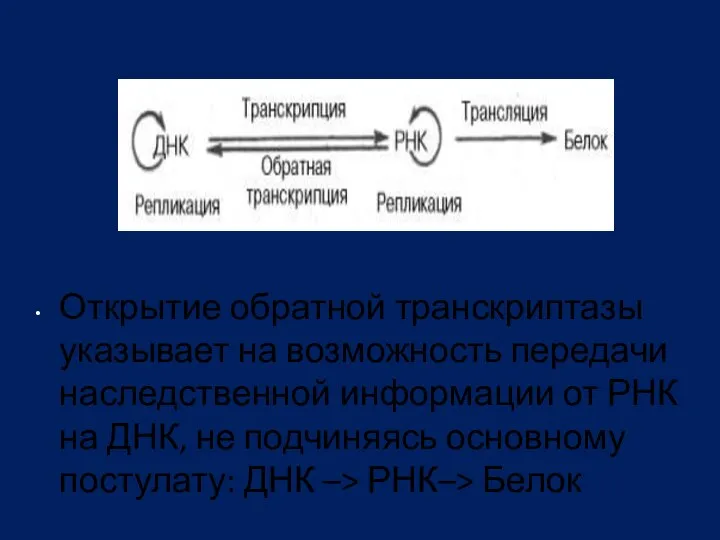
Открытие обратной транскриптазы указывает на возможность передачи наследственной информации от РНК
Открытие обратной транскриптазы указывает на возможность передачи наследственной информации от РНК

Экспрессия генов
Процесс транскрипции – биосинтез РНК на матрице ДНК. Начальная
Экспрессия генов
Процесс транскрипции – биосинтез РНК на матрице ДНК. Начальная

передача (переписывание) информации, содержащейся в последовательности нуклеотидных остатков в ДНК-матрице,
передача (переписывание) информации, содержащейся в последовательности нуклеотидных остатков в ДНК-матрице,

Транскриптон – часть молекулы ДНК
транскрибируется в процессе биосинтеза РНК на
Транскриптон – часть молекулы ДНК
транскрибируется в процессе биосинтеза РНК на

Независимая транскрипция гена – ген обладает собственным промотором и терминатором транскрипции.
Независимая транскрипция гена – ген обладает собственным промотором и терминатором транскрипции.

Ген эукариот является мозаичной структурой, содержащей наряду с кодирующими (экзонами) также
Ген эукариот является мозаичной структурой, содержащей наряду с кодирующими (экзонами) также

Соотношение информативной и неинформативной частей в транскриптонах
Эукариотов – 1:9
Прокариотов – 9:1
Соотношение информативной и неинформативной частей в транскриптонах
Эукариотов – 1:9
Прокариотов – 9:1

Соседние транскриптоны могут быть отделены друг от друга нетранскрибируемыми участками
Биологический смысл:
Соседние транскриптоны могут быть отделены друг от друга нетранскрибируемыми участками
Биологический смысл:

Ферменты транскрипции
В 1960 г. открыт в двух лабораториях США (Дж. Хервиц
Ферменты транскрипции
В 1960 г. открыт в двух лабораториях США (Дж. Хервиц

Фермент РНК-полимераза катализирует транскрипцию как экзонов, так и интронов
Для синтеза РНК,
Фермент РНК-полимераза катализирует транскрипцию как экзонов, так и интронов
Для синтеза РНК,
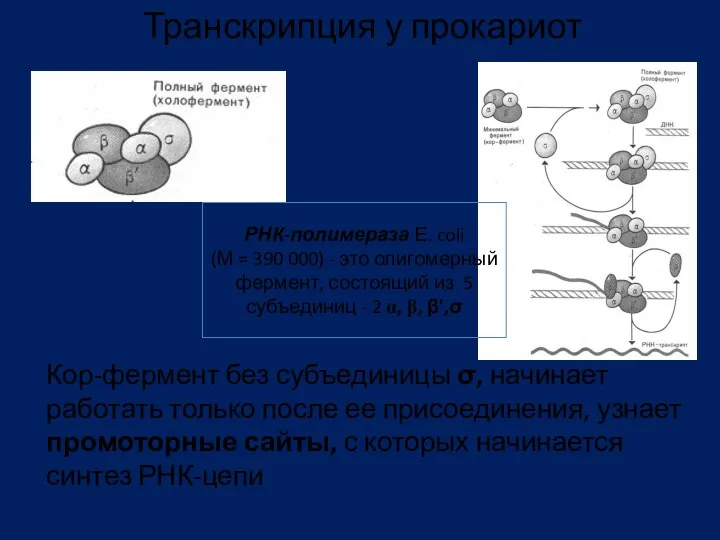
Кор-фермент без субъединицы σ, начинает работать только после ее присоединения, узнает
Кор-фермент без субъединицы σ, начинает работать только после ее присоединения, узнает

Транскрипция
Инициация
Элонгация
Терминация
-35 -10 +1
----TTGACA ----------------TATAAT-------------CAT-----
----AACTGT----------------ATATTA-------------GTA----
16-19 bp 4-8
Транскрипция
Инициация
Элонгация
Терминация
-35 -10 +1
----TTGACA ----------------TATAAT-------------CAT-----
----AACTGT----------------ATATTA-------------GTA----
16-19 bp 4-8

Факторы транскрипции регуляторные белки обеспечивающие у эукариот работу РНК-pol
ТАТА-
Факторы транскрипции регуляторные белки обеспечивающие у эукариот работу РНК-pol
ТАТА-

РНК-pol связывается с промотором, происходит локальное раскручивание двойной спирали ДНК (примерно
РНК-pol связывается с промотором, происходит локальное раскручивание двойной спирали ДНК (примерно

Копирование смысловой (ДНК+) цепи
Направление 5’-3’
Первый нуклеотид всегда АТФ или ГТФ, затем
Копирование смысловой (ДНК+) цепи
Направление 5’-3’
Первый нуклеотид всегда АТФ или ГТФ, затем

Факторы элонгации (Е,Н,F) повышают активность РНК-pol и облегчают расхождение цепей. Скорость
Факторы элонгации (Е,Н,F) повышают активность РНК-pol и облегчают расхождение цепей. Скорость

Завершение синтеза РНК
Происходит в сайтах терминации (транскрипционные терминаторы – GC –
Завершение синтеза РНК
Происходит в сайтах терминации (транскрипционные терминаторы – GC –
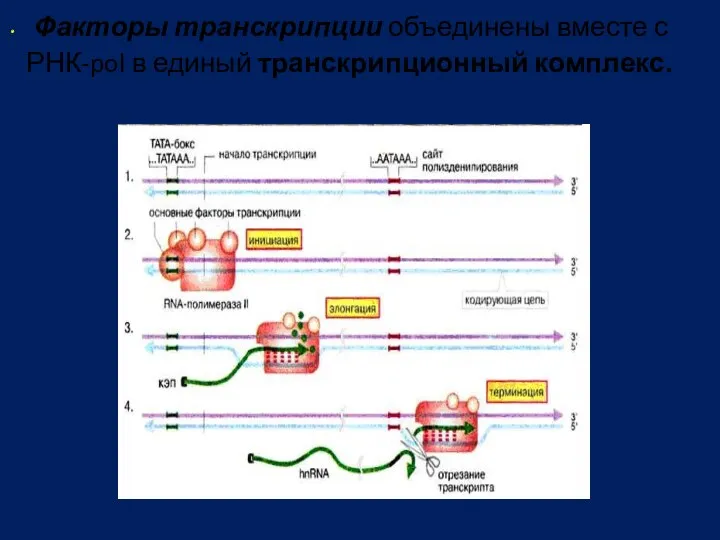
Факторы транскрипции объединены вместе с
РНК-pol в единый транскрипционный комплекс.
Факторы транскрипции объединены вместе с
РНК-pol в единый транскрипционный комплекс.

Транскрипция сходна с репликацей
В основе механизма процесса лежит принцип комплементарности (А=У,
Транскрипция сходна с репликацей
В основе механизма процесса лежит принцип комплементарности (А=У,

Транскрипция отличается от репликации рядом особенностей:
1. Не требует синтеза праймера.
2. Использует
Транскрипция отличается от репликации рядом особенностей:
1. Не требует синтеза праймера.
2. Использует

Регуляторные элементы, стимулирующие транскрипцию связанных с ними генов, называются энхансерами (усилителями,
Регуляторные элементы, стимулирующие транскрипцию связанных с ними генов, называются энхансерами (усилителями,

Транскрипционные факторы
(Транс- действующие факторы)
это белки, т. е. продукты других, независимых генов,
Транскрипционные факторы
(Транс- действующие факторы)
это белки, т. е. продукты других, независимых генов,

Процессинг первичных
транскриптов РНК
Между транскрипцией и трансляцией существует важное звено –
Процессинг первичных
транскриптов РНК
Между транскрипцией и трансляцией существует важное звено –
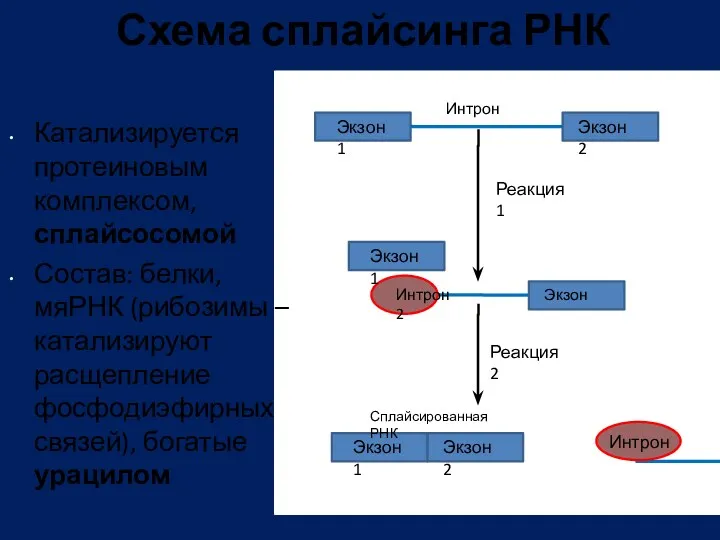
Схема сплайсинга РНК
Катализируется протеиновым комплексом, сплайсосомой
Состав: белки, мяРНК (рибозимы – катализируют
Схема сплайсинга РНК
Катализируется протеиновым комплексом, сплайсосомой
Состав: белки, мяРНК (рибозимы – катализируют

Регуляция генной экспрессии
Не все белки синтезируются постоянно
Некоторые белки синтезируются только при
Регуляция генной экспрессии
Не все белки синтезируются постоянно
Некоторые белки синтезируются только при
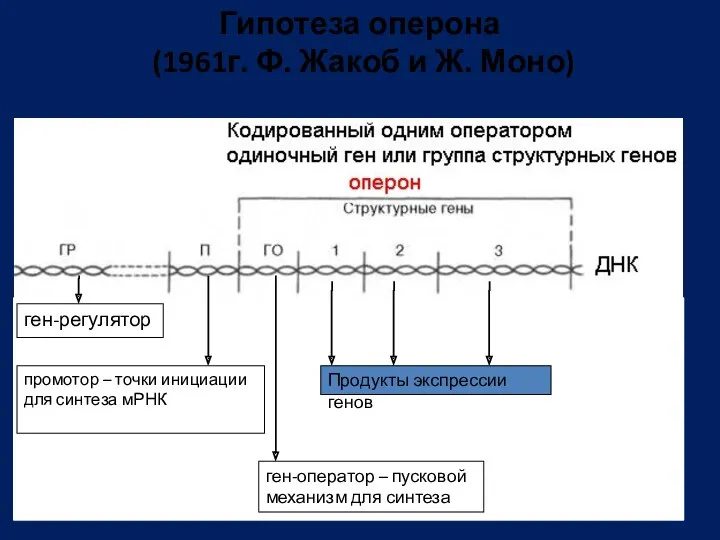
Гипотеза оперона (1961г. Ф. Жакоб и Ж. Моно)
ген-регулятор
промотор – точки инициации
Гипотеза оперона (1961г. Ф. Жакоб и Ж. Моно)
ген-регулятор
промотор – точки инициации

Организация хроматина в дифференцированных клетках многоклеточного организма
Гетерохроматин (стабильно репрессированные участки)- ДНК
Организация хроматина в дифференцированных клетках многоклеточного организма
Гетерохроматин (стабильно репрессированные участки)- ДНК

В разных типах клеток в область эухроматина попадают разные гены, а
В разных типах клеток в область эухроматина попадают разные гены, а

Регуляторные зоны ДНК
Энхансеры и сайленсоры
различны по числу и расположению на
Регуляторные зоны ДНК
Энхансеры и сайленсоры
различны по числу и расположению на

Это способ записи информации об
аминоксилотной последовательности белков с помощью
Это способ записи информации об
аминоксилотной последовательности белков с помощью

СВОЙСТВА ГЕНЕТИЧЕСКОГО КОДА
Триплетность - кодовое число = 3. Три нуклеотидных остатка
СВОЙСТВА ГЕНЕТИЧЕСКОГО КОДА
Триплетность - кодовое число = 3. Три нуклеотидных остатка

Код непрерывен, отсутствие «знаков препинания», т.е. сигналов, указывающих на конец одного
Код непрерывен, отсутствие «знаков препинания», т.е. сигналов, указывающих на конец одного
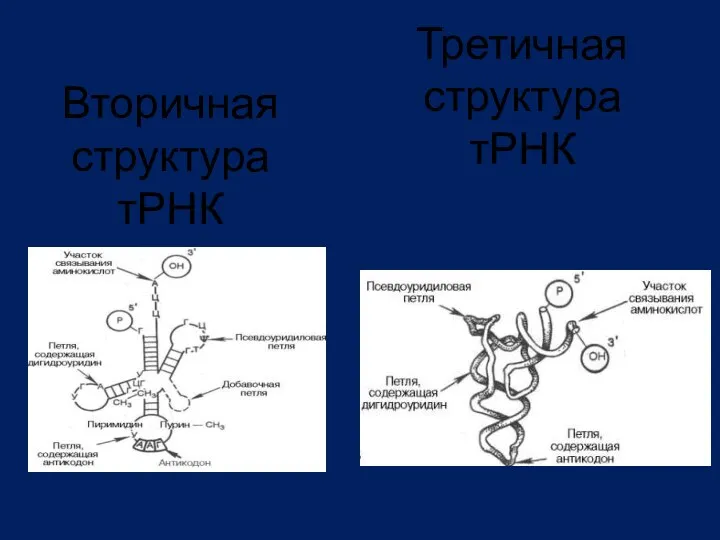
Вторичная структура тРНК
Третичная структура тРНК
Вторичная структура тРНК
Третичная структура тРНК

Значение активирования АК:
аатРНК - это транспортная форма АК
Каждая АК получила свой
Значение активирования АК:
аатРНК - это транспортная форма АК
Каждая АК получила свой
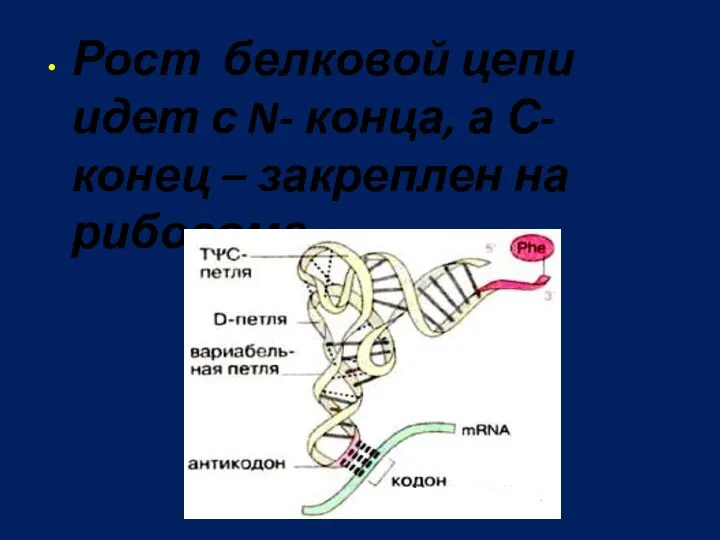
Рост белковой цепи идет с N- конца, а С-конец – закреплен
Рост белковой цепи идет с N- конца, а С-конец – закреплен
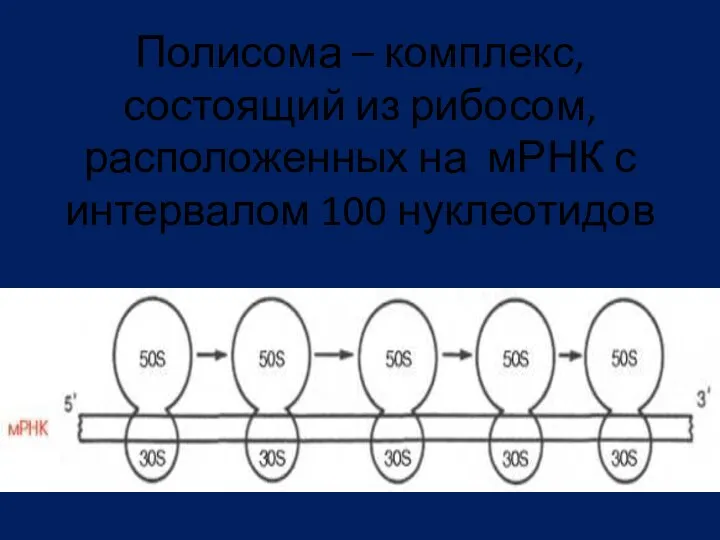
Полисома – комплекс, состоящий из рибосом, расположенных на мРНК с интервалом
Полисома – комплекс, состоящий из рибосом, расположенных на мРНК с интервалом

Функции субъединиц рибосомы в процессе трансляции:
Малая субъединица присоединяет мРНК и
Функции субъединиц рибосомы в процессе трансляции:
Малая субъединица присоединяет мРНК и
 Красная книга Казахстана
Красная книга Казахстана Бионика. Бионическая архитектура
Бионика. Бионическая архитектура Семейство сельдевых, тресковых, камбаловых и ставридовых рыб
Семейство сельдевых, тресковых, камбаловых и ставридовых рыб Бауырдың биохимиясы
Бауырдың биохимиясы Кісткові і Хрящові риби. 7 клас
Кісткові і Хрящові риби. 7 клас Лікарські рослини, які містять полісахариди
Лікарські рослини, які містять полісахариди Сезонные изменения в природе и жизнедеятельности организмов
Сезонные изменения в природе и жизнедеятельности организмов презентация по биологии 7 класс Плоские черви
презентация по биологии 7 класс Плоские черви Объекты биотехнологии. Биообъекты, применяемые на практике
Объекты биотехнологии. Биообъекты, применяемые на практике Лисичка желтая
Лисичка желтая Семейство розоцветных.
Семейство розоцветных. Эндокринная система
Эндокринная система Физиология сенсорных систем
Физиология сенсорных систем Биоэнергетика. Биологическое окисление. Биологические виды энергии
Биоэнергетика. Биологическое окисление. Биологические виды энергии Реакции электрофильного замещения в ароматическом и гетероциклическом рядах SE. (Лекция 5)
Реакции электрофильного замещения в ароматическом и гетероциклическом рядах SE. (Лекция 5) Exotic animals
Exotic animals Строение семян
Строение семян Определение соотношения размеров хвоинок и шишек хвойных деревьев. Выявление наиболее полезных хвойных деревьев
Определение соотношения размеров хвоинок и шишек хвойных деревьев. Выявление наиболее полезных хвойных деревьев Carbohydrates
Carbohydrates Метод селекции породы собак сиба-ину
Метод селекции породы собак сиба-ину Строение. Функции. Значение кожи
Строение. Функции. Значение кожи Царство Растения
Царство Растения Молекулярный уровень
Молекулярный уровень Основные компоненты клетки
Основные компоненты клетки Ознаки живих організмів
Ознаки живих організмів Моногибридное скрещивание
Моногибридное скрещивание Состояние и перспективы развития пчеловодства в Республике Башкортостан. История развития, ведущие ученые
Состояние и перспективы развития пчеловодства в Республике Башкортостан. История развития, ведущие ученые о тиграх
о тиграх