- Особенности функционирования белков с четвертичной структурой

Содержание
- 2. Олигомеры и мономеры Олигомерные белки способны взаимодействовать с несколькими лигандами в центрах, удаленных от активного центра.
- 3. Миоглобин Миоглобин относят к классу гемсодержащих белков, т.е. он содержит простетическую группу - гем, довольно прочно
- 4. Гем Гем - молекула, имеющая структуру циклического тетрапиррола, где 4 пиррольных кольца соединены метиленовыми мостиками и
- 5. Строение гема
- 6. Апомиоглобин Апомиоглобин - белковая часть миоглобина; первичная структура представлена последовательностью из 153 аминокислот, которые во вторичной
- 7. Миоглобин Гем - специфический лиганд апомиоглобина, присоединяющийся к белковой части в углублении между двумя спиралями F
- 8. Миоглобин с молекулой кислорода
- 9. Трехмерная структура миоглобина
- 10. Гемоглобины Гемоглобины - родственные белки, находящиеся в эритроцитах. Эти белки выполняют 2 важные функции: Перенос О2
- 11. Гемоглобины Конформация отдельных протомеров гемоглобина удивительно напоминает конформацию миоглобина, несмотря на то, что в первичной структуре
- 12. Строение гемоглобина А
- 13. Роль гистидина E7 в функционировании миоглобина и гемоглобина Гем имеет высокое сродство к оксиду углерода (СО).
- 14. Роль гистидина E7 в функционировании миоглобина и гемоглобина Если бы сродство тема к СО не уменьшалось
- 15. Связывание гемоглобина с кислородом Олигомерная структура гемоглобина обеспечивает быстрое насыщение его кислородом в лёгких, возможность отщепления
- 16. Связывание кислорода гемом и изменение белковой части
- 17. Кооперативное изменение в молекуле гемоглобина В дезоксигемоглобине благодаря ковалентной связи с белковой частью атом Fe2+ выступает
- 18. Зависимость степени насыщения гемов различных белков от содержания кислорода
- 19. Кривая диссоциации оксимиоглобина Кривые диссоциации кислорода для миоглобина и гемоглобина в зависимости от парциального давления кислорода.
- 20. Кривая диссоциации оксигемоглобина Кривая диссоциации для гемоглобина имеет сигмоидную форму (S-образную). Это указывает на то, что
- 21. Эффект Бора Степень сродства гемоглобина к молекуле кислорода помимо кооперативного эффекта регулируется присоединением катиона водорода. Эта
- 23. Скачать презентацию

Олигомеры и мономеры
Олигомерные белки способны взаимодействовать с несколькими лигандами в центрах,
Олигомеры и мономеры
Олигомерные белки способны взаимодействовать с несколькими лигандами в центрах,

Миоглобин
Миоглобин относят к классу гемсодержащих белков, т.е. он содержит простетическую группу
Миоглобин
Миоглобин относят к классу гемсодержащих белков, т.е. он содержит простетическую группу

Гем
Гем - молекула, имеющая структуру циклического тетрапиррола, где 4 пиррольных кольца соединены
Гем
Гем - молекула, имеющая структуру циклического тетрапиррола, где 4 пиррольных кольца соединены
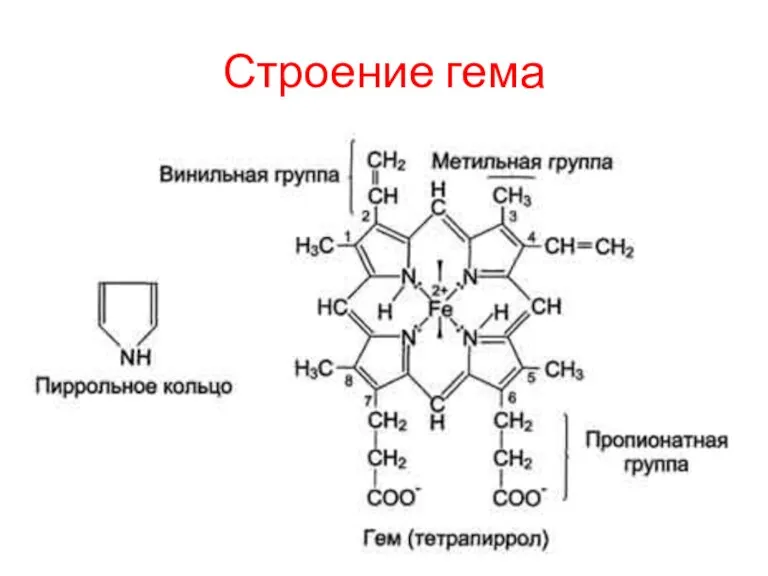
Строение гема
Строение гема

Апомиоглобин
Апомиоглобин - белковая часть миоглобина; первичная структура представлена последовательностью из 153 аминокислот,
Апомиоглобин
Апомиоглобин - белковая часть миоглобина; первичная структура представлена последовательностью из 153 аминокислот,

Миоглобин
Гем - специфический лиганд апомиоглобина, присоединяющийся к белковой части в углублении
Миоглобин
Гем - специфический лиганд апомиоглобина, присоединяющийся к белковой части в углублении
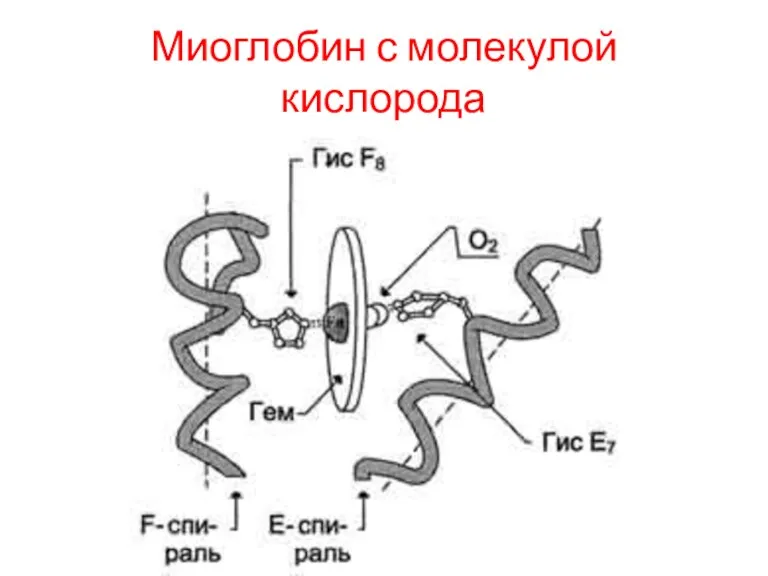
Миоглобин с молекулой кислорода
Миоглобин с молекулой кислорода
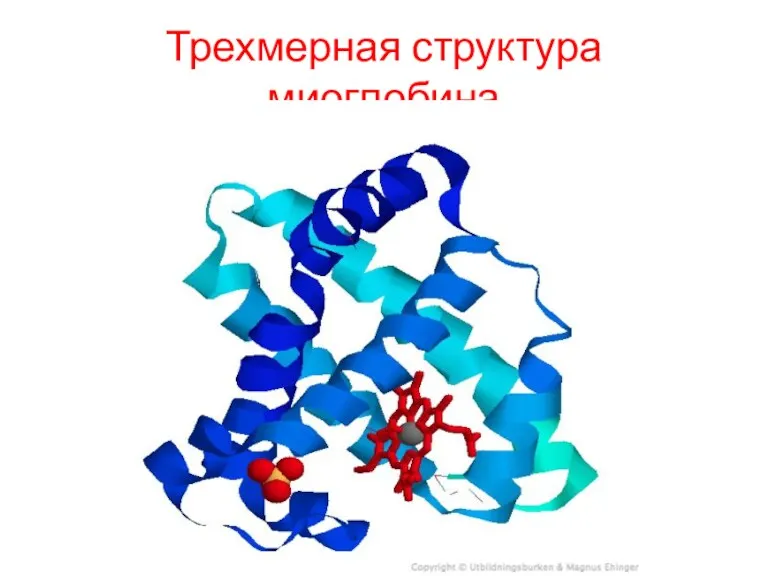
Трехмерная структура миоглобина
Трехмерная структура миоглобина

Гемоглобины
Гемоглобины - родственные белки, находящиеся в эритроцитах. Эти белки выполняют 2
Гемоглобины
Гемоглобины - родственные белки, находящиеся в эритроцитах. Эти белки выполняют 2

Гемоглобины
Конформация отдельных протомеров гемоглобина удивительно напоминает конформацию миоглобина, несмотря на то,
Гемоглобины
Конформация отдельных протомеров гемоглобина удивительно напоминает конформацию миоглобина, несмотря на то,
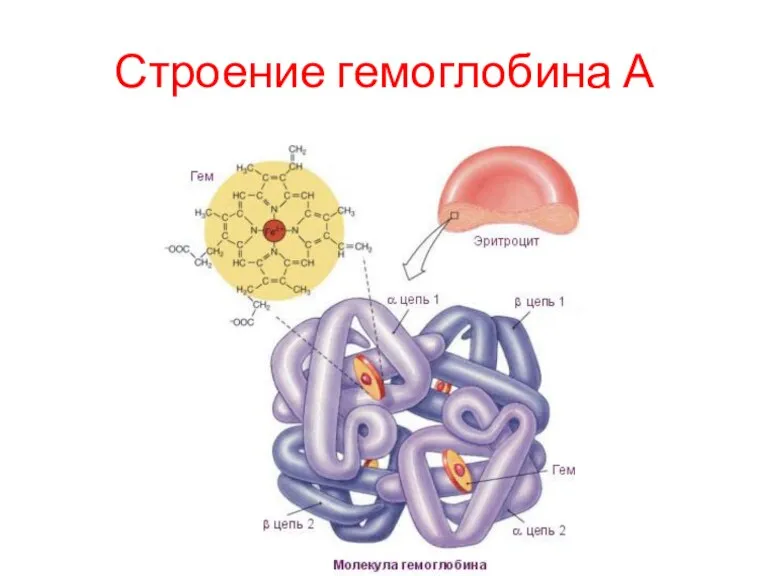
Строение гемоглобина А
Строение гемоглобина А

Роль гистидина E7 в функционировании миоглобина и гемоглобина
Гем имеет высокое сродство к
Роль гистидина E7 в функционировании миоглобина и гемоглобина
Гем имеет высокое сродство к
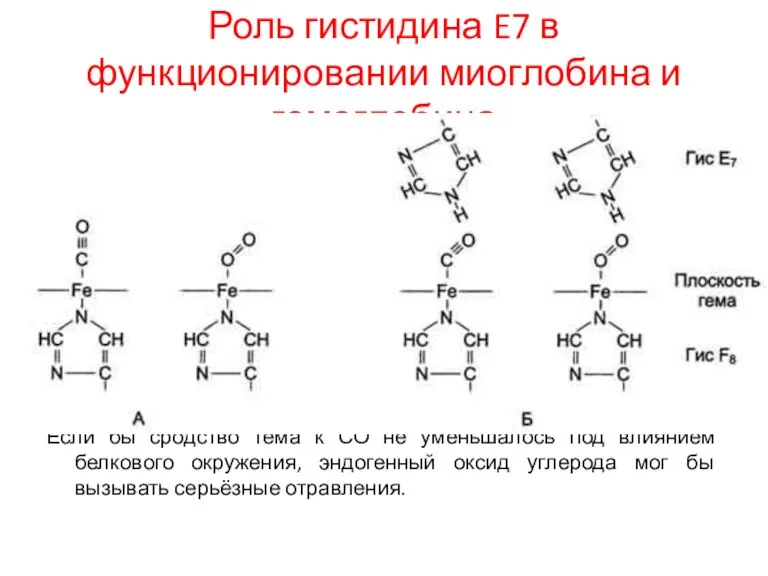
Роль гистидина E7 в функционировании миоглобина и гемоглобина
Если бы сродство тема к
Роль гистидина E7 в функционировании миоглобина и гемоглобина
Если бы сродство тема к

Связывание гемоглобина с кислородом
Олигомерная структура гемоглобина обеспечивает быстрое насыщение его кислородом
Связывание гемоглобина с кислородом
Олигомерная структура гемоглобина обеспечивает быстрое насыщение его кислородом
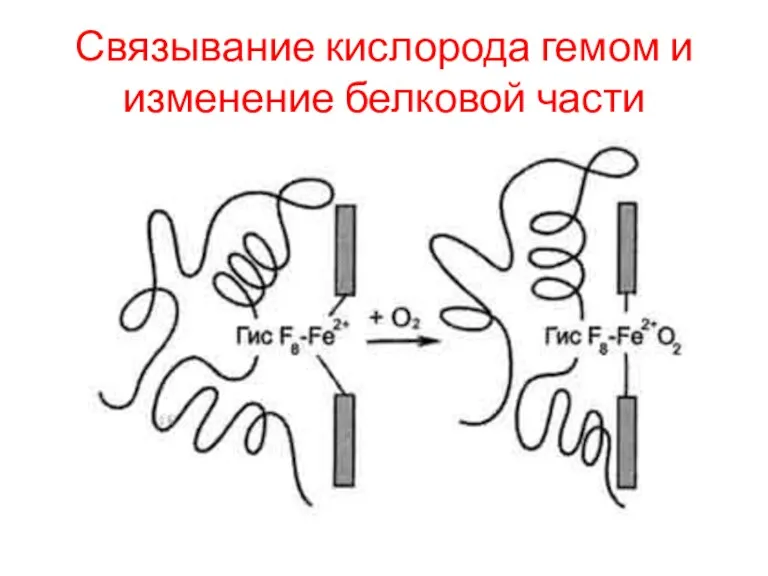
Связывание кислорода гемом и изменение белковой части
Связывание кислорода гемом и изменение белковой части

Кооперативное изменение в молекуле гемоглобина
В дезоксигемоглобине благодаря ковалентной связи с белковой
Кооперативное изменение в молекуле гемоглобина
В дезоксигемоглобине благодаря ковалентной связи с белковой
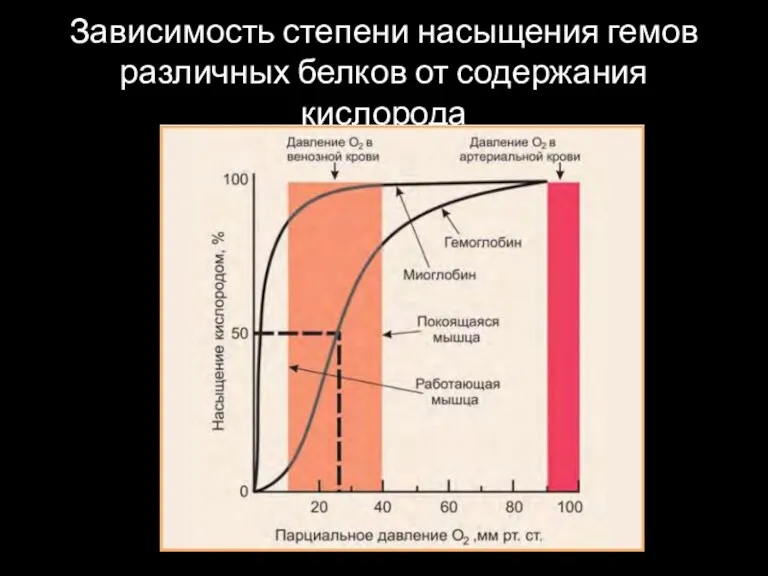
Зависимость степени насыщения гемов различных белков от содержания кислорода
Зависимость степени насыщения гемов различных белков от содержания кислорода

Кривая диссоциации оксимиоглобина
Кривые диссоциации кислорода для миоглобина и гемоглобина в зависимости
Кривая диссоциации оксимиоглобина
Кривые диссоциации кислорода для миоглобина и гемоглобина в зависимости

Кривая диссоциации оксигемоглобина
Кривая диссоциации для гемоглобина имеет сигмоидную форму (S-образную). Это
Кривая диссоциации оксигемоглобина
Кривая диссоциации для гемоглобина имеет сигмоидную форму (S-образную). Это

Эффект Бора
Степень сродства гемоглобина к молекуле кислорода помимо кооперативного эффекта регулируется
Эффект Бора
Степень сродства гемоглобина к молекуле кислорода помимо кооперативного эффекта регулируется
 Размножение и развитие птиц
Размножение и развитие птиц Рэспубліканскі конкурс Юны натуралiст. Намінацыя Калекцыя кветак
Рэспубліканскі конкурс Юны натуралiст. Намінацыя Калекцыя кветак Адаптация и здоровье.Психотипы человека и их склонность к возникновению заболеваний
Адаптация и здоровье.Психотипы человека и их склонность к возникновению заболеваний Паразиты.Подразделения
Паразиты.Подразделения Красная книга. 1 класс УМК Гармония
Красная книга. 1 класс УМК Гармония Селекция и её достижения
Селекция и её достижения Работа мышц
Работа мышц Самый большой отряд насекомых
Самый большой отряд насекомых Роль слюнных желёз в пищеварении
Роль слюнных желёз в пищеварении Вегетативная (автономная) нервная система
Вегетативная (автономная) нервная система Путешествие в удивительный мир растений
Путешествие в удивительный мир растений Многообразие простейших
Многообразие простейших Решение задач по генетике
Решение задач по генетике Вегетативные органы растений: корень
Вегетативные органы растений: корень Презентация к уроку биология 10 класс Питание клетки.
Презентация к уроку биология 10 класс Питание клетки. Қан құрамындағы газдардың тұтқырлығын қамтамасыз ететін жастық ерекшеліктер
Қан құрамындағы газдардың тұтқырлығын қамтамасыз ететін жастық ерекшеліктер Вітаміни. Види
Вітаміни. Види Генетика как наука. Основные понятия дисциплины и ее связь с другими науками. История развития науки
Генетика как наука. Основные понятия дисциплины и ее связь с другими науками. История развития науки Тип черви
Тип черви Строение растений. Цветы, соцветия и плоды
Строение растений. Цветы, соцветия и плоды Формы размножения организмов
Формы размножения организмов Удивительный мир животных. Интерактивная экскурсия
Удивительный мир животных. Интерактивная экскурсия Кәдімгі цикорий-Сichorium intybus l
Кәдімгі цикорий-Сichorium intybus l Тағамдық микронутриенттер. Микронутриенттердің жеткіліксіздігі. Микронутриенттер тапшылығының алдын-алу стратегиясы
Тағамдық микронутриенттер. Микронутриенттердің жеткіліксіздігі. Микронутриенттер тапшылығының алдын-алу стратегиясы Общая физиология нервной системы и возбудимых тканей
Общая физиология нервной системы и возбудимых тканей Cells
Cells Строение глаза
Строение глаза Урок-презентация на тему Тип Членистоногие
Урок-презентация на тему Тип Членистоногие