- Особенности поперечно-полосатых и гладких мышц. Титин (тайтин/коннектин)

Содержание
- 2. Вопросы для мини-проверки 1. Почему актин назвали «актином»? 2. Какие изоформы или классы актинов синтезируются в
- 3. Молекула G-актина несет прочно связанную АТФ, который при переходе в F-актин медленно гидролизуется до АДФ –
- 4. Новости науки «Шесть шагов»: учёные из 153 стран объявили чрезвычайную климатическую ситуацию на планете Более 11
- 5. Главные авторы статьи — американские экологи Уильям Риппл и Кристофер Вулф из Орегонского университета. Они отметили,
- 6. Согласно данным, проанализированным учёными, за последние 10 лет массовая вырубка лесов увеличилась на 50%, выбросы углекислого
- 7. Учёные назвали шесть первоочередных задач («шесть шагов»), на решении которых необходимо сконцентрировать все силы. В своеобразную
- 8. Толстая нить. Каждая миозиновая нить состоит из 300-400 молекул миозина и С-белка. Половина молекул миозина обращена
- 9. Комплексы Ф-актина, ТНТ и ТМ (Повторение) Нити Ф-актина схема Подлубная, 1990.
- 10. Итак, уже более 60 лет известно, что в саркомерах поперечно-полосатых мышц присутствуют два типа белковых нитей:
- 11. Early electron microscope images of a formalin-fixed skeletal muscle myofibril published by Draper and Hodge (1950).
- 12. Саркомеры летательной мышцы пчелы (Trombitas & Tigyi-Sebes, 1974) Визуализация третьего типа нитей в саркомере (близ Z-диска)
- 13. Названия третьего типа нитей: «промежуточные» – видимые на микроснимках в промежутках между концами толстых и тонких
- 14. Электронные микрофотографии коннектина (Maruyama et al., 1977; Sawada et al., 1983) Увеличение 53000 Увеличение 68000 Коннектин
- 15. Электрофоретическое открытие титина (коннектина) ДСН-гель-электрофорез проведён в крупнопористых 3-4% полиакриламидных гелях Wang et al., 1979 Maruyama
- 16. Титин – «Титаник» среди белков Титин/тайтин Титаник -Титин – самый большой из известных на сегодняшний день
- 17. Domain structure of titin-isoforms and binding sites of titin ligands. A, Layout of titin-isoforms in the
- 18. служит остовом для сборки миозиновых нитей и саркомера во время миофибриллогенеза; регулирует длину миозинсодержащих нитей и
- 19. На основе связывания Т- cap с LIM белком (CsRP3), участвующим в дифференцировке миоцитов (Knoll et al.,
- 20. Предполагается, что пассивное натяжение, развиваемое титином при растяжении саркомера, регулирует ток калия в клетке (Furukawa et
- 21. Миостати́н — белок, который подавляет рост и дифференцировку мышечной ткани. Исследуются блокаторы действия миостатина через миостатиновые
- 22. M. Carrion-Vazquez et al., 2000 Развитие титином пассивного напряжения при растяжении Эластичные элементы I-области титина иммуноглобулин-подобные
- 23. Эластичные функции титина (продолжение) W. Linke, 2000 Функция эластичного саркомерного цитоскелета - Титин вносит вклад в
- 24. Однако при этом необходимо также, чтобы титин, связанный в саркомере с миозиновыми нитями, взаимодействовал и с
- 25. Участие I-области титина в процессах внутриклеточной сигнализации Связывание I-области титина с комплексом мышечных белков с анкириновыми
- 26. Функции титина в А-диске и М-линии саркомера Эта часть молекулы титина нерастяжима и, предположительно, выполняет несколько
- 27. Участие киназного домена титина в регуляции генной экспрессии и обмена белков Механическая нагрузка Активация киназы титина
- 28. Наши ЭМ исследования титина Выпрямленные молекулы титина Агрегаты титина Шкала 200 нм Электронные микрофотографии одиночных выпрямленных
- 29. Агрегаты титина скелетных мышц в растворе 0.1 М KCl, 10 мМ имидазол, рН 7.0. Шкала 200
- 30. Связывание титина с нитями миозина Контрольные нити миозина Нити миозина в присутствии титина Вихлянцев, 2005
- 31. Взаимовие паракристаллов Ф-актина (картинка слева) с титином скелетных мышц кролика в растворе 50 мМ MgCl2, 5
- 33. Скачать презентацию

Вопросы для мини-проверки
1. Почему актин назвали «актином»?
2. Какие изоформы или
Вопросы для мини-проверки
1. Почему актин назвали «актином»?
2. Какие изоформы или

Молекула G-актина несет прочно связанную АТФ, который при
переходе в F-актин медленно
Молекула G-актина несет прочно связанную АТФ, который при переходе в F-актин медленно

Новости науки
«Шесть шагов»: учёные из 153 стран объявили чрезвычайную климатическую ситуацию
Новости науки
«Шесть шагов»: учёные из 153 стран объявили чрезвычайную климатическую ситуацию

Главные авторы статьи — американские экологи Уильям Риппл и Кристофер Вулф
Главные авторы статьи — американские экологи Уильям Риппл и Кристофер Вулф

Согласно данным, проанализированным учёными, за последние 10 лет массовая вырубка лесов
Согласно данным, проанализированным учёными, за последние 10 лет массовая вырубка лесов

Учёные назвали шесть первоочередных задач («шесть шагов»), на решении которых необходимо
Учёные назвали шесть первоочередных задач («шесть шагов»), на решении которых необходимо
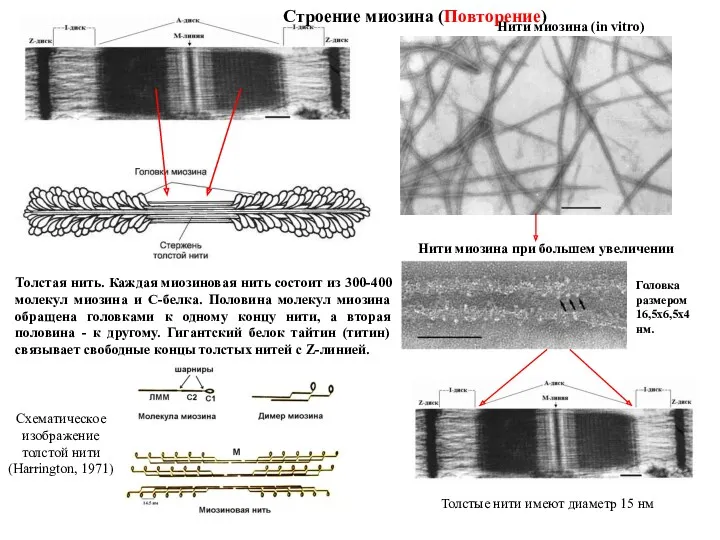
Толстая нить. Каждая миозиновая нить состоит из 300-400 молекул миозина и
Толстая нить. Каждая миозиновая нить состоит из 300-400 молекул миозина и
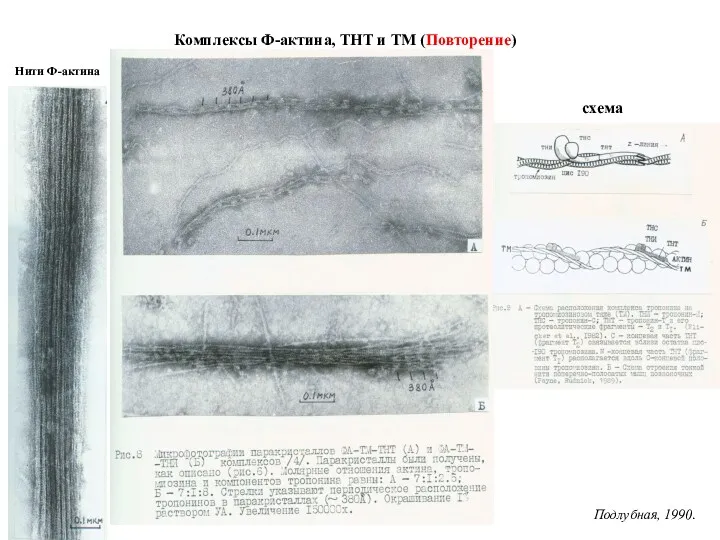
Комплексы Ф-актина, ТНТ и ТМ (Повторение)
Нити Ф-актина
схема
Подлубная, 1990.
Комплексы Ф-актина, ТНТ и ТМ (Повторение)
Нити Ф-актина
схема
Подлубная, 1990.

Итак, уже более 60 лет известно, что в саркомерах поперечно-полосатых мышц
Итак, уже более 60 лет известно, что в саркомерах поперечно-полосатых мышц

Early electron microscope
images of a formalin-fixed
skeletal muscle myofibril
published by Draper and
Early electron microscope
images of a formalin-fixed
skeletal muscle myofibril
published by Draper and

Саркомеры летательной мышцы пчелы (Trombitas & Tigyi-Sebes, 1974)
Визуализация третьего типа
Саркомеры летательной мышцы пчелы (Trombitas & Tigyi-Sebes, 1974)
Визуализация третьего типа

Названия третьего типа нитей:
«промежуточные» – видимые на микроснимках в промежутках
Названия третьего типа нитей:
«промежуточные» – видимые на микроснимках в промежутках
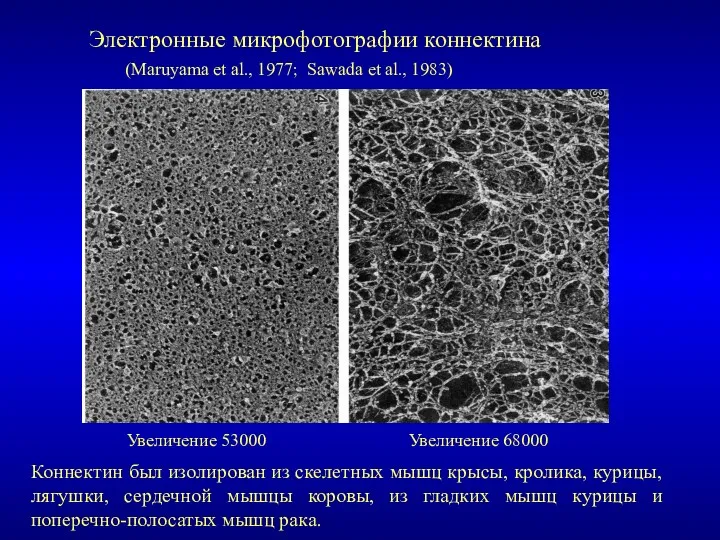
Электронные микрофотографии коннектина (Maruyama et al., 1977; Sawada et al., 1983)
Электронные микрофотографии коннектина (Maruyama et al., 1977; Sawada et al., 1983)

Электрофоретическое открытие титина (коннектина)
ДСН-гель-электрофорез проведён в крупнопористых 3-4% полиакриламидных гелях
Wang et
Электрофоретическое открытие титина (коннектина)
ДСН-гель-электрофорез проведён в крупнопористых 3-4% полиакриламидных гелях
Wang et

Титин – «Титаник» среди белков
Титин/тайтин
Титаник
-Титин – самый большой из
известных на
Титин – «Титаник» среди белков
Титин/тайтин
Титаник
-Титин – самый большой из
известных на
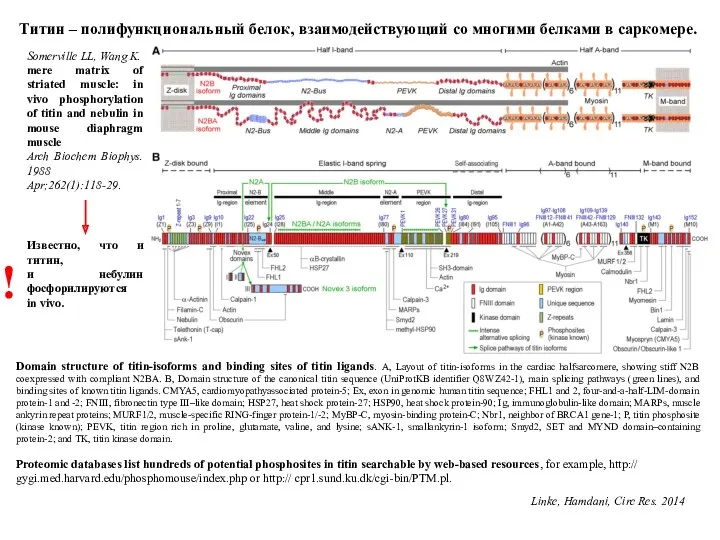
Domain structure of titin-isoforms and binding sites of titin ligands. A,
Domain structure of titin-isoforms and binding sites of titin ligands. A,
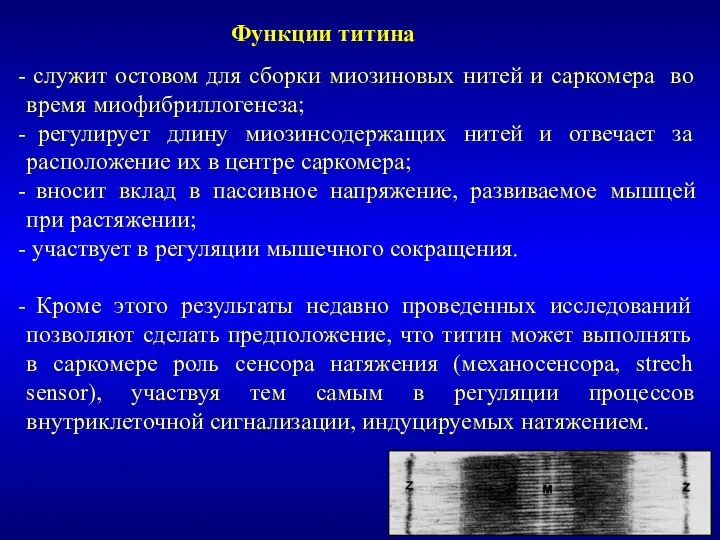
служит остовом для сборки миозиновых нитей и саркомера во время
служит остовом для сборки миозиновых нитей и саркомера во время

На основе связывания Т- cap с LIM белком (CsRP3), участвующим
На основе связывания Т- cap с LIM белком (CsRP3), участвующим

Предполагается, что пассивное натяжение, развиваемое титином при растяжении саркомера, регулирует ток
Предполагается, что пассивное натяжение, развиваемое титином при растяжении саркомера, регулирует ток

Миостати́н — белок, который подавляет рост и дифференцировку мышечной ткани.
Исследуются
Миостати́н — белок, который подавляет рост и дифференцировку мышечной ткани.
Исследуются
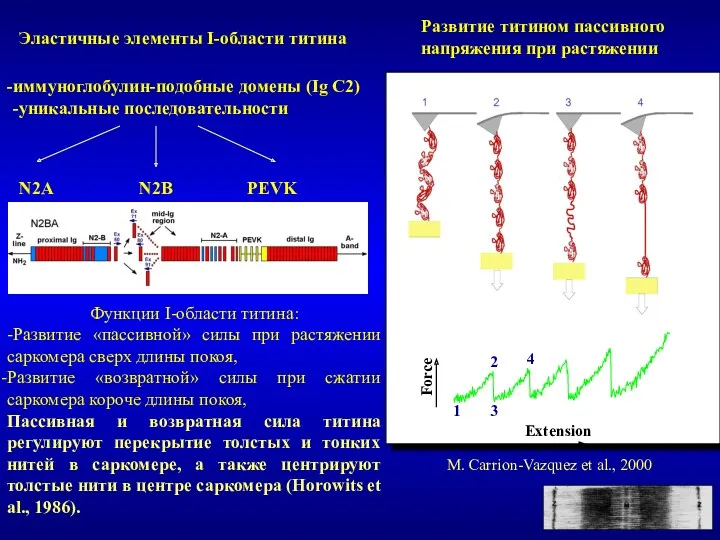
M. Carrion-Vazquez et al., 2000
Развитие титином пассивного
напряжения при растяжении
Эластичные элементы
M. Carrion-Vazquez et al., 2000
Развитие титином пассивного
напряжения при растяжении
Эластичные элементы
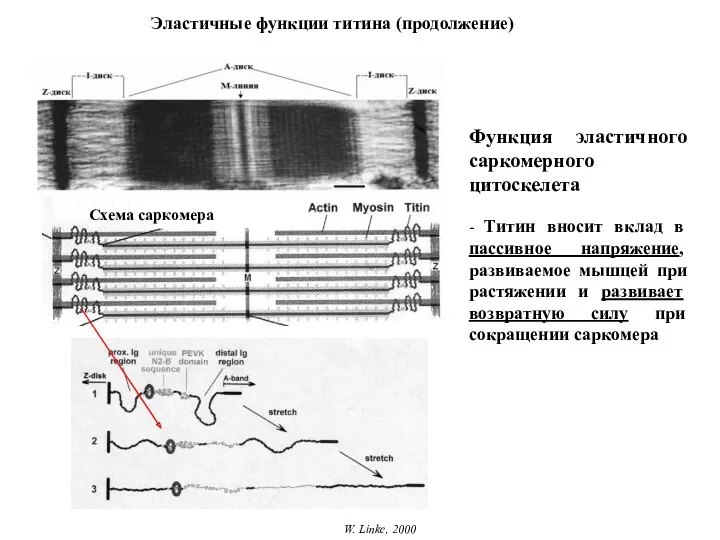
Эластичные функции титина (продолжение)
W. Linke, 2000
Функция эластичного саркомерного цитоскелета
- Титин вносит
Эластичные функции титина (продолжение)
W. Linke, 2000
Функция эластичного саркомерного цитоскелета
- Титин вносит
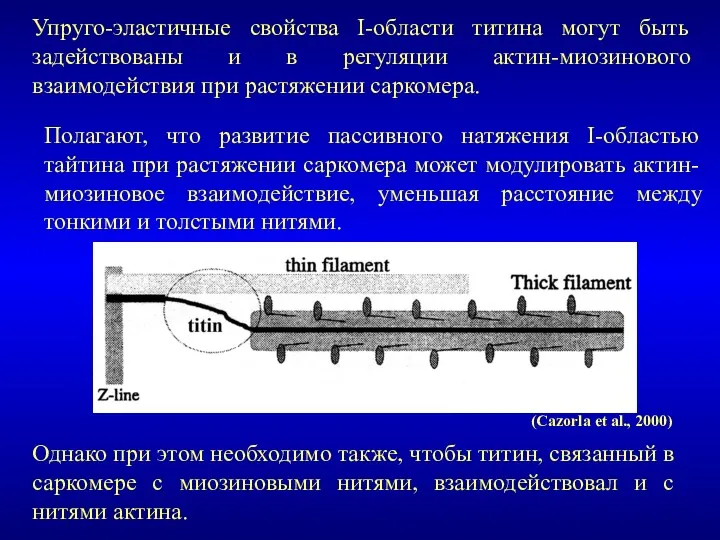
Однако при этом необходимо также, чтобы титин, связанный в саркомере с
Однако при этом необходимо также, чтобы титин, связанный в саркомере с

Участие I-области титина в процессах внутриклеточной сигнализации
Связывание I-области титина
с комплексом мышечных
Участие I-области титина в процессах внутриклеточной сигнализации
Связывание I-области титина
с комплексом мышечных

Функции титина в А-диске и М-линии саркомера
Эта часть молекулы титина нерастяжима
Функции титина в А-диске и М-линии саркомера
Эта часть молекулы титина нерастяжима

Участие киназного домена титина в регуляции генной экспрессии и обмена белков
Механическая
Участие киназного домена титина в регуляции генной экспрессии и обмена белков
Механическая

Наши ЭМ исследования титина
Выпрямленные молекулы титина
Агрегаты титина
Шкала 200 нм
Электронные микрофотографии
Наши ЭМ исследования титина
Выпрямленные молекулы титина
Агрегаты титина
Шкала 200 нм
Электронные микрофотографии
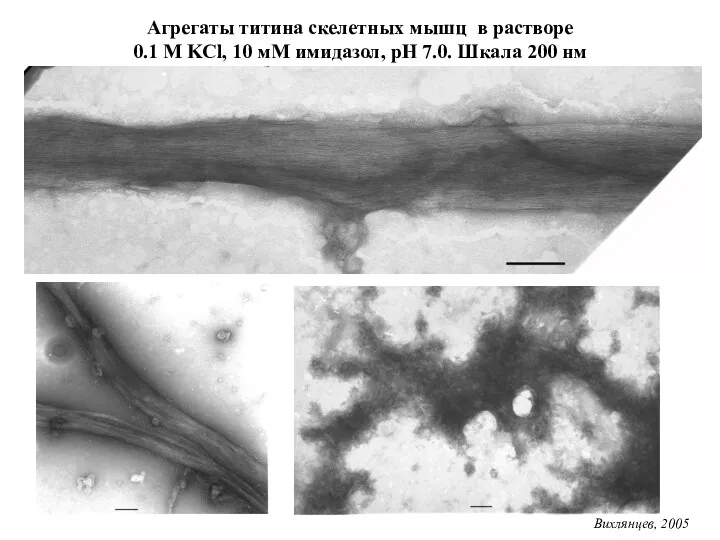
Агрегаты титина скелетных мышц в растворе
0.1 М KCl, 10 мМ
Агрегаты титина скелетных мышц в растворе 0.1 М KCl, 10 мМ
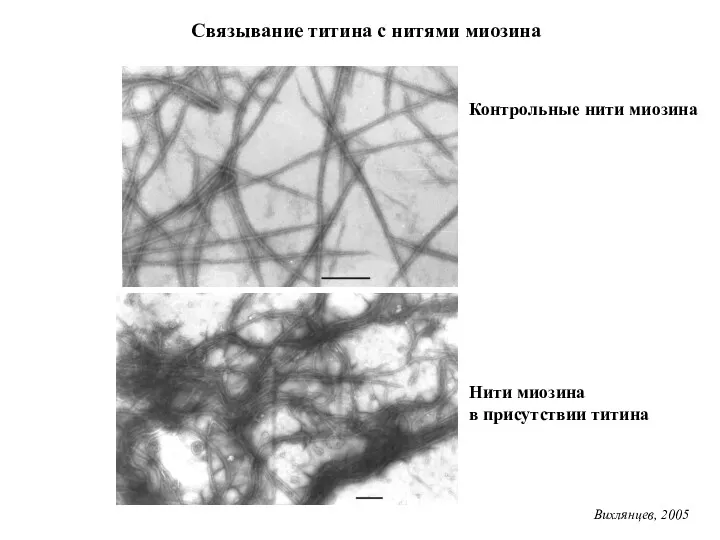
Связывание титина с нитями миозина
Контрольные нити миозина
Нити миозина
в присутствии титина
Вихлянцев,
Связывание титина с нитями миозина
Контрольные нити миозина
Нити миозина
в присутствии титина
Вихлянцев,
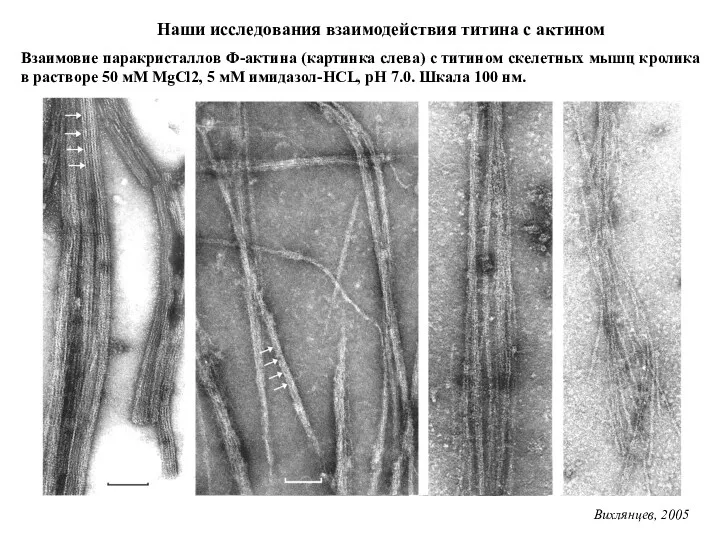
Взаимовие паракристаллов Ф-актина (картинка слева) с титином скелетных мышц кролика в
Взаимовие паракристаллов Ф-актина (картинка слева) с титином скелетных мышц кролика в
 Широколиственный лес
Широколиственный лес Царство грибов
Царство грибов Многообразие живой природы
Многообразие живой природы конспект урока по биологии 8 класс с мультимедийной презентацией Пищеварительная система
конспект урока по биологии 8 класс с мультимедийной презентацией Пищеварительная система Проект учащихся 5 класса ВЛИЯНИЕ СВЕТА НА ПРОРАСТАНИЕ СЕМЯН ПОДСОЛНЕЧНИКА
Проект учащихся 5 класса ВЛИЯНИЕ СВЕТА НА ПРОРАСТАНИЕ СЕМЯН ПОДСОЛНЕЧНИКА Органическое вещество почвы
Органическое вещество почвы Круговорот веществ в природе
Круговорот веществ в природе Цветок. Плод. Семя
Цветок. Плод. Семя Семейства растений. Семейства крестоцветные и розоцветные
Семейства растений. Семейства крестоцветные и розоцветные Плаунтәрізді өсімдіктер
Плаунтәрізді өсімдіктер Общая характеристика грибов
Общая характеристика грибов Биоценозы. Концепция биоценоза
Биоценозы. Концепция биоценоза Органические вещества в составе клетки
Органические вещества в составе клетки Уровни организации живых организмов
Уровни организации живых организмов Лесные ягоды. Грибы
Лесные ягоды. Грибы Отдел зеленые водоросли. Класс харовые
Отдел зеленые водоросли. Класс харовые Методы изучения биологии
Методы изучения биологии Рослини Миколаївщини
Рослини Миколаївщини Презентация по биологии Внутренняя среда организма. Кровь
Презентация по биологии Внутренняя среда организма. Кровь Определение пола и наследование, сцепленное с полом
Определение пола и наследование, сцепленное с полом Гормоны строение, классификация, функции, биосинтез, транспорт. (Лекция 10)
Гормоны строение, классификация, функции, биосинтез, транспорт. (Лекция 10) презент плесн.грибы
презент плесн.грибы Хрящевые рыбы
Хрящевые рыбы Покровные и проводящие растительные ткани
Покровные и проводящие растительные ткани Червона книга України
Червона книга України Формирование УУД на уроках биологии и химии.
Формирование УУД на уроках биологии и химии. Атлас млекопитающих Москвы и Подмосковья
Атлас млекопитающих Москвы и Подмосковья Презентация Вид. Критерии вида
Презентация Вид. Критерии вида