- Популяции. Типы пространственной структуры популяций

Содержание
- 2. Популяция – элементарная единица эволюционного процесса и существования биоты. Толерантность к факторам среды у популяции значительно
- 3. Пространственная структура популяций Популяции различных видов размещаются разными способами по занимаемой ими территории, т.е. имеют разную
- 4. Типы пространственной структуры популяций: равномерное – каждая особь удалена от другой на определенное расстояние (антагонистические отношения)
- 5. Типы пространственной структуры популяций
- 6. Поло-возрастная структура популяций – соотношение особей, принадлежащих разным полам и находящихся на разных стадиях жизненного цикла.
- 7. Функциональная структура популяции – «разделение труда между особями». Особенно типична для организмов с общественным поведением. Молодые
- 8. Функциональная специализация с распределением функций внутри популяции термитов
- 9. Динамика популяций ПРИНЦИП ЭКСПОНЕНЦИАЛЬНОГО РОСТА ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ В БЛАГОПРИЯТНОЙ И НЕОГРАНИЧЕННОЙ СТАЦИОНАРНОЙ СРЕДЕ Это один из
- 10. Модель Томаса Мальтуса Nt = N0ert где Nt – численность во времени t N0 – исходная
- 11. О возможности геометрического роста численности организмов упоминали Ж. Бюффон и К. Линней, расчеты Т. Мальтуса оказали
- 12. Так, Чарльз Дарвин рассчитывал потенциальные возможности роста популяций разных организмов Бактерия Bacillus coli делится каждые 20
- 13. А одна инфузория (Paramecium caudatum) могла бы за несколько дней произвести такое количество протоплазмы, которая по
- 14. Модель Мальтуса теоретически отражает возможности нарастания численности любой популяции в условиях отсутствия каких либо ограничений и
- 15. Динамика популяции – процессы, протекающие в популяции за промежуток времени (рождаемость, смертность и скорость роста) Стабильный
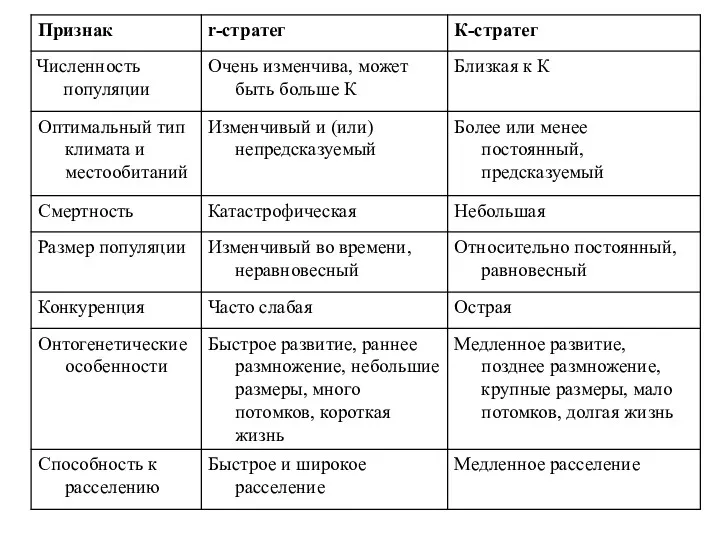
- 16. Двум типам динамики соответствуют две противоположные жизненные стратегии. (Р. Мак-Артур, Э. Вильсон) К-стратегия характерна для стабильных
- 17. В природе (и в экспериментах) динамика популяций часто хорошо соответствует так называемой логистической (сигмоидной, S-образной) кривой.
- 18. Для r-стратегов используется экспоненциальная модель. В ней рост численности не зависит от плотности, но врожденная скорость
- 20. Кривые выживания – показывают общую картину динамики численности популяций. Три основных типа кривых выживания. Тип 1
- 21. Общую оценку численности популяции в каком-то определенном местообитании в некое время t можно получить по формуле
- 22. ГИПОТЕЗА САМОРЕГУЛЯЦИИ ПОПУЛЯЦИЙ Любая популяция способна, в принципе, сама регулировать свою численность так, чтобы не подрывались
- 23. Типы регуляции численности популяции: Внутривидовая (внутрипопуляционная) конкуренция – важнейший регулятор динамики популяций. С нарастанием численности популяции
- 24. Саморегуляция осуществляется через: "механизмы стресса" (гормональные сдвиги под влиянием нервного возбуждения, тормозят деятельность половых желез, изменяются
- 25. Циклические изменения численности зайца (Lepus americanus) и рыси (Felis canadensis [Lynx lynx]) в Канадской Арктике, представленные
- 26. dN1 / dt = N1 • (r1 – b • N2) dN2 / dt = N2
- 27. ОБОБЩЕННАЯ МОДЕЛЬ Колмогорова где a(N1) – функция естественного прироста жертвы; V(N1) – трофическая функция хищника; K(N1)
- 28. УРАВНЕНИЕ (ФОРМУЛА) Ивлева Эта формула задает следующий вид трофической функции хищника от одной из жертв: V(N1)
- 30. Скачать презентацию

Популяция – элементарная единица эволюционного процесса и существования биоты.
Толерантность к факторам
Популяция – элементарная единица эволюционного процесса и существования биоты.
Толерантность к факторам

Пространственная структура популяций
Популяции различных видов размещаются разными способами по занимаемой ими
Пространственная структура популяций
Популяции различных видов размещаются разными способами по занимаемой ими

Типы пространственной структуры популяций:
равномерное – каждая особь удалена от другой на
Типы пространственной структуры популяций:
равномерное – каждая особь удалена от другой на
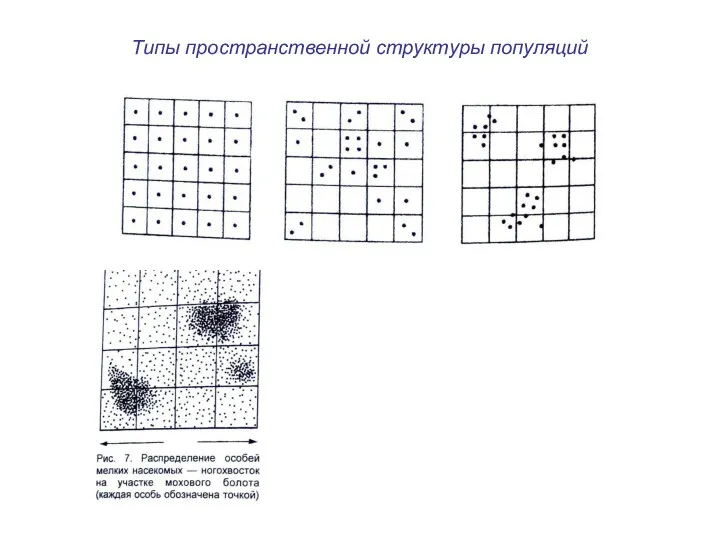
Типы пространственной структуры популяций
Типы пространственной структуры популяций
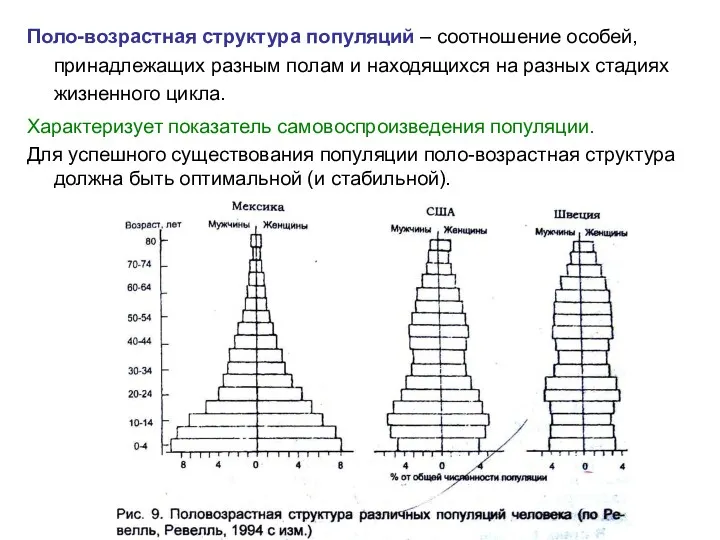
Поло-возрастная структура популяций – соотношение особей, принадлежащих разным полам и находящихся
Поло-возрастная структура популяций – соотношение особей, принадлежащих разным полам и находящихся

Функциональная структура популяции – «разделение труда между особями».
Особенно типична для
Функциональная структура популяции – «разделение труда между особями».
Особенно типична для

Функциональная специализация с распределением функций внутри популяции термитов
Функциональная специализация с распределением функций внутри популяции термитов

Динамика популяций
ПРИНЦИП ЭКСПОНЕНЦИАЛЬНОГО РОСТА
ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ В БЛАГОПРИЯТНОЙ И НЕОГРАНИЧЕННОЙ СТАЦИОНАРНОЙ
Динамика популяций ПРИНЦИП ЭКСПОНЕНЦИАЛЬНОГО РОСТА ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ В БЛАГОПРИЯТНОЙ И НЕОГРАНИЧЕННОЙ СТАЦИОНАРНОЙ
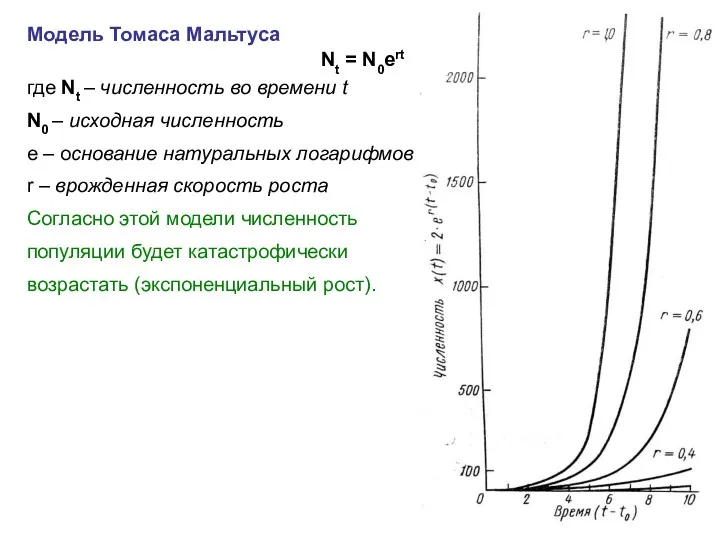
Модель Томаса Мальтуса
Nt = N0ert
где Nt – численность во времени t
N0
Модель Томаса Мальтуса
Nt = N0ert
где Nt – численность во времени t
N0

О возможности геометрического роста численности организмов упоминали Ж. Бюффон и К.
О возможности геометрического роста численности организмов упоминали Ж. Бюффон и К.

Так, Чарльз Дарвин рассчитывал потенциальные возможности роста популяций разных организмов
Бактерия Bacillus
Так, Чарльз Дарвин рассчитывал потенциальные возможности роста популяций разных организмов
Бактерия Bacillus

А одна инфузория (Paramecium caudatum) могла бы за несколько дней произвести
А одна инфузория (Paramecium caudatum) могла бы за несколько дней произвести

Модель Мальтуса теоретически отражает возможности нарастания численности любой популяции в условиях
Модель Мальтуса теоретически отражает возможности нарастания численности любой популяции в условиях

Динамика популяции – процессы, протекающие в популяции за промежуток времени (рождаемость,
Динамика популяции – процессы, протекающие в популяции за промежуток времени (рождаемость,

Двум типам динамики соответствуют две противоположные жизненные стратегии. (Р. Мак-Артур, Э.
Двум типам динамики соответствуют две противоположные жизненные стратегии. (Р. Мак-Артур, Э.
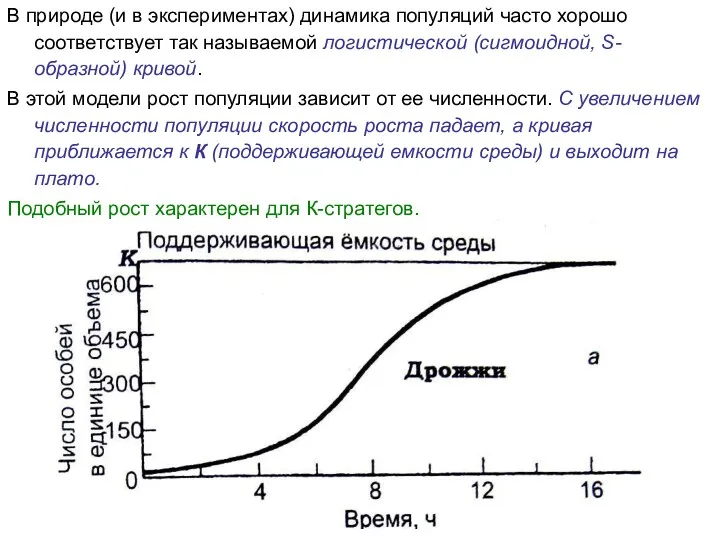
В природе (и в экспериментах) динамика популяций часто хорошо соответствует так
В природе (и в экспериментах) динамика популяций часто хорошо соответствует так
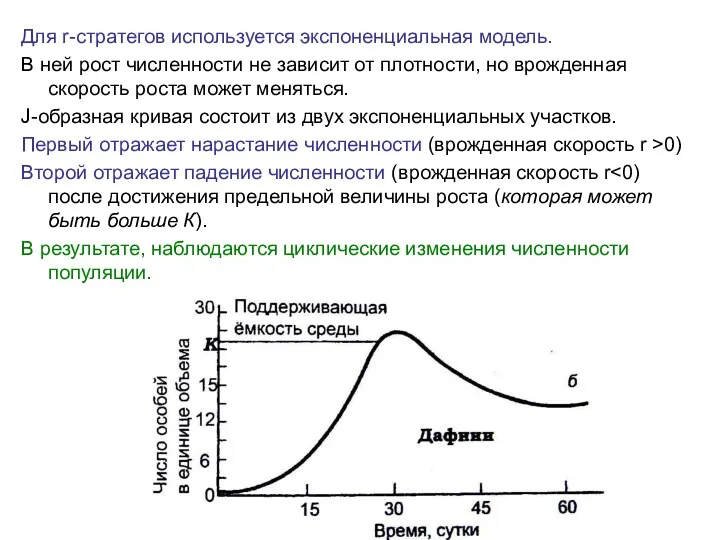
Для r-стратегов используется экспоненциальная модель.
В ней рост численности не зависит
Для r-стратегов используется экспоненциальная модель.
В ней рост численности не зависит
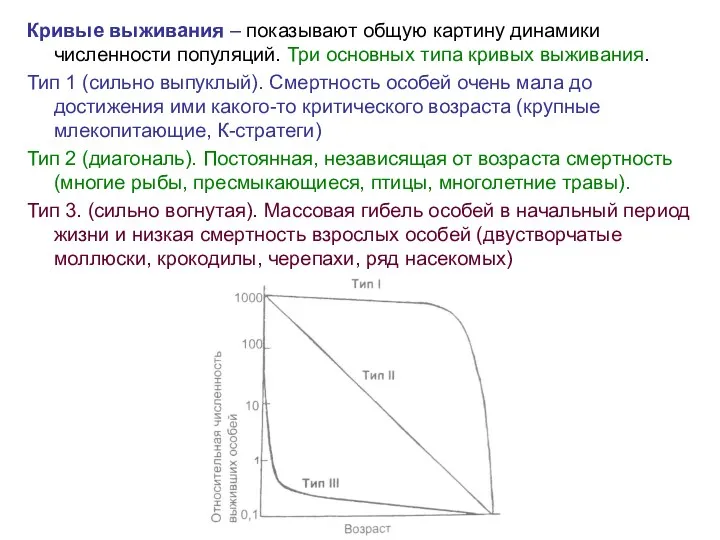
Кривые выживания – показывают общую картину динамики численности популяций. Три основных
Кривые выживания – показывают общую картину динамики численности популяций. Три основных

Общую оценку численности популяции в каком-то определенном местообитании в некое время
Общую оценку численности популяции в каком-то определенном местообитании в некое время

ГИПОТЕЗА САМОРЕГУЛЯЦИИ ПОПУЛЯЦИЙ
Любая популяция способна, в принципе, сама регулировать свою
ГИПОТЕЗА САМОРЕГУЛЯЦИИ ПОПУЛЯЦИЙ
Любая популяция способна, в принципе, сама регулировать свою

Типы регуляции численности популяции:
Внутривидовая (внутрипопуляционная) конкуренция – важнейший регулятор динамики популяций.
Типы регуляции численности популяции:
Внутривидовая (внутрипопуляционная) конкуренция – важнейший регулятор динамики популяций.

Саморегуляция осуществляется через:
"механизмы стресса" (гормональные сдвиги под влиянием нервного возбуждения, тормозят
Саморегуляция осуществляется через:
"механизмы стресса" (гормональные сдвиги под влиянием нервного возбуждения, тормозят
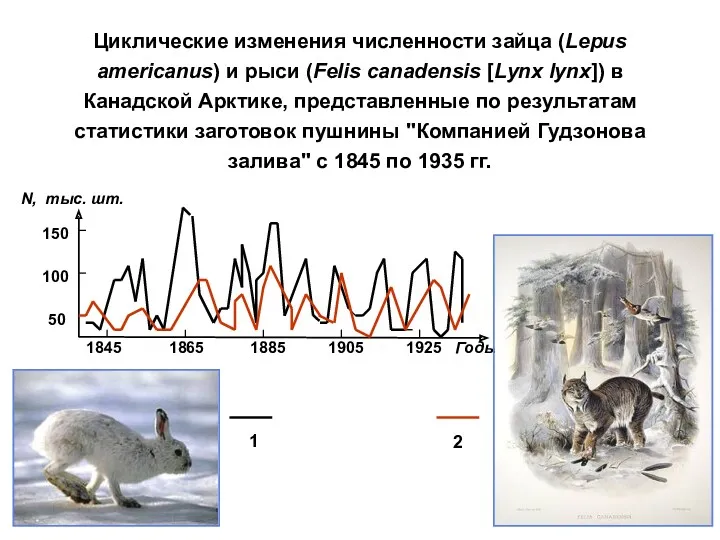
Циклические изменения численности зайца (Lepus americanus) и рыси (Felis canadensis [Lynx
Циклические изменения численности зайца (Lepus americanus) и рыси (Felis canadensis [Lynx

dN1 / dt = N1 • (r1 – b • N2)
dN2
dN1 / dt = N1 • (r1 – b • N2)
dN2

ОБОБЩЕННАЯ МОДЕЛЬ Колмогорова
где a(N1) – функция естественного прироста жертвы; V(N1)
ОБОБЩЕННАЯ МОДЕЛЬ Колмогорова
где a(N1) – функция естественного прироста жертвы; V(N1)

УРАВНЕНИЕ (ФОРМУЛА) Ивлева
Эта формула задает следующий вид трофической функции хищника от
УРАВНЕНИЕ (ФОРМУЛА) Ивлева
Эта формула задает следующий вид трофической функции хищника от
 Эволюция растений
Эволюция растений презентация Самые высокие горы
презентация Самые высокие горы Оценка качества почвы с помощью растения – биоиндикатора
Оценка качества почвы с помощью растения – биоиндикатора Мінеральне та повітряне живлення рослин
Мінеральне та повітряне живлення рослин Биология оқыту үрдісінде тәрбиелеу Ептіліктердің және білімдердің қалыптастыру
Биология оқыту үрдісінде тәрбиелеу Ептіліктердің және білімдердің қалыптастыру Генетические основы селекции.
Генетические основы селекции. Поджелудочная железа
Поджелудочная железа Интерактивная игра Кто? Как? Почему? Играя, вспоминаем для 8 класса (по материалу I полугодия)
Интерактивная игра Кто? Как? Почему? Играя, вспоминаем для 8 класса (по материалу I полугодия) Лекарственные растения
Лекарственные растения Презентация к уроку биологии на тему Витамины.
Презентация к уроку биологии на тему Витамины. Gardis. Хвойные декоративные растения (каталог)
Gardis. Хвойные декоративные растения (каталог) Семейство Rosacea. Розоцветные
Семейство Rosacea. Розоцветные Эволюция нервной, кровеносной и выделительной систем органов
Эволюция нервной, кровеносной и выделительной систем органов Общая характеристика типа Моллюски
Общая характеристика типа Моллюски Биопестициды
Биопестициды В гостях у природы
В гостях у природы Тканевый обмен веществ
Тканевый обмен веществ Морфологія, будова та хімічний склад бактеріальної клітини
Морфологія, будова та хімічний склад бактеріальної клітини Функциональное состояние в структуре поведения
Функциональное состояние в структуре поведения Болезни эмбрионов с/х птицы
Болезни эмбрионов с/х птицы Иван Владимирович Мичурин (1855-1935)
Иван Владимирович Мичурин (1855-1935) Основні закономірності незалежного та зчепленого успадкування
Основні закономірності незалежного та зчепленого успадкування Дигибридное скрещивание. Закон независимого наследования признаков
Дигибридное скрещивание. Закон независимого наследования признаков Презентация по биологии Класс Птицы для 7 класса.
Презентация по биологии Класс Птицы для 7 класса. Свободные жирные кислоты и кальциевый гомеостаз скелетно-мышечных клеток
Свободные жирные кислоты и кальциевый гомеостаз скелетно-мышечных клеток Основные понятия о системе движений. Задача биомеханики двигательных действий. (Лекция 2)
Основные понятия о системе движений. Задача биомеханики двигательных действий. (Лекция 2) Бесполое размножение клетки
Бесполое размножение клетки Биотехнологии. Генная инженерия
Биотехнологии. Генная инженерия