- Регуляция экспрессии генов. (Лекция 5)

Содержание
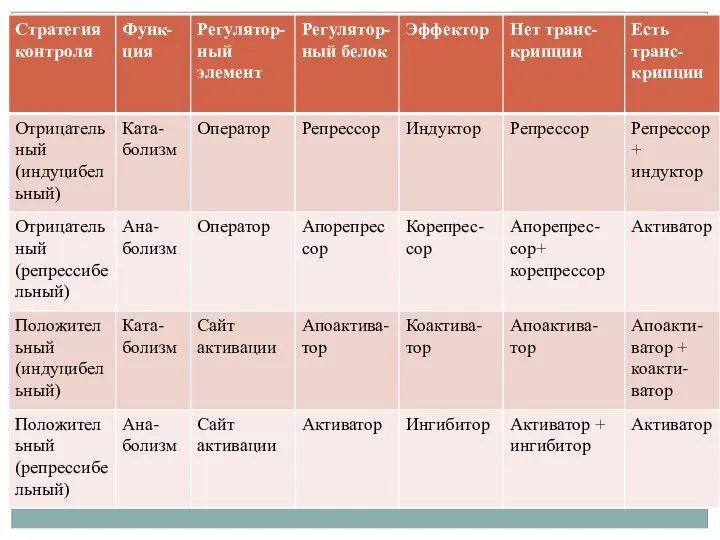
- 2. Регуляция прокариот Регуляция метаболизма, поведения, морфологии бактерий осуществляется с помощью контроля экспрессии генов. Прокариоты способны быстро
- 3. Регуляция прокариот Контроль осуществляется на уровне инициации и элонгации транскрипции, трансляции и посттрансляции. Археи схожи с
- 4. Регуляция прокариот Примеры: Почвенный микроорганизм Bacillus subtilis при понижении концентрации питательных веществ запускает процессы споруляции. Патогенные
- 5. Уровни регуляции в трех доменах жизни Механизмы транскрипции и трансляции схожи у всех живых организмов. Бактерии
- 6. Регуляция у Бактерий
- 7. Регуляция у Архей
- 8. Регуляция у Эукариот
- 9. История открытия регуляции 1900г. Эмиль Дюкло обнаружил спобность Aspergillus niger продуцировать фермент гидролизующий сукрозу только в
- 10. История открытия регуляции Позже Ледерберг, Моно, Жакоб и Парди обнаружили наличие гена, синтезирующего продукт ингибирующий запуск
- 11. Регуляция прокариот Исследования Е. coli показали, что у бактерий существуют ферменты 3 типов: конститутивные, присутствующие в
- 12. Лактозный оперон При выращивании E.coli на среде с лактозой, содержание лактозы достигает 3000 молекул на клетку,
- 13. Лактозный оперон
- 14. Лактозный оперон
- 17. Регуляция лактозного оперона
- 19. Инициация транскрипции
- 20. Отрицательный контроль индуцибельных генов Пример: лактозный оперон. В отсутствии индуктора, репрессорный белок блокирует транскрипцию. При появлении
- 21. Отрицательный контроль репрессибельных генов Пример: оперон синтеза триптофана. В отсутствии корепрессора белок-репрессор не способен связываться с
- 22. Положительный контроль индуцибельных генов Пример: оперон деградации арабинозы. Активаторный белок (апоактиватор) способен связываться с сайтом активации
- 23. Положительный контроль репрессибельных генов Пример: оперон синтеза лейцина. Активаторный белок связывается с ДНК и запускает транскрипцию.
- 25. Скачать презентацию

Регуляция прокариот
Регуляция метаболизма, поведения, морфологии бактерий осуществляется с помощью контроля экспрессии
Регуляция прокариот
Регуляция метаболизма, поведения, морфологии бактерий осуществляется с помощью контроля экспрессии

Регуляция прокариот
Контроль осуществляется на уровне инициации и элонгации транскрипции, трансляции и
Регуляция прокариот
Контроль осуществляется на уровне инициации и элонгации транскрипции, трансляции и

Регуляция прокариот
Примеры: Почвенный микроорганизм Bacillus subtilis при понижении концентрации питательных веществ
Регуляция прокариот
Примеры: Почвенный микроорганизм Bacillus subtilis при понижении концентрации питательных веществ

Уровни регуляции в трех доменах жизни
Механизмы транскрипции и трансляции схожи
Уровни регуляции в трех доменах жизни
Механизмы транскрипции и трансляции схожи
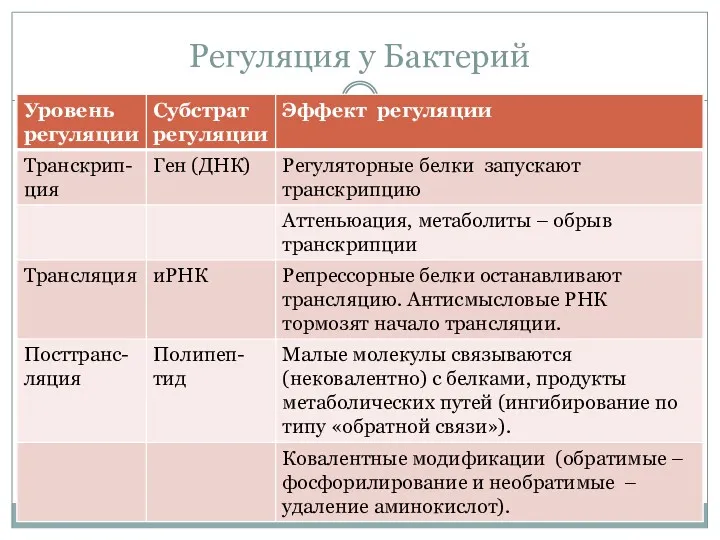
Регуляция у Бактерий
Регуляция у Бактерий
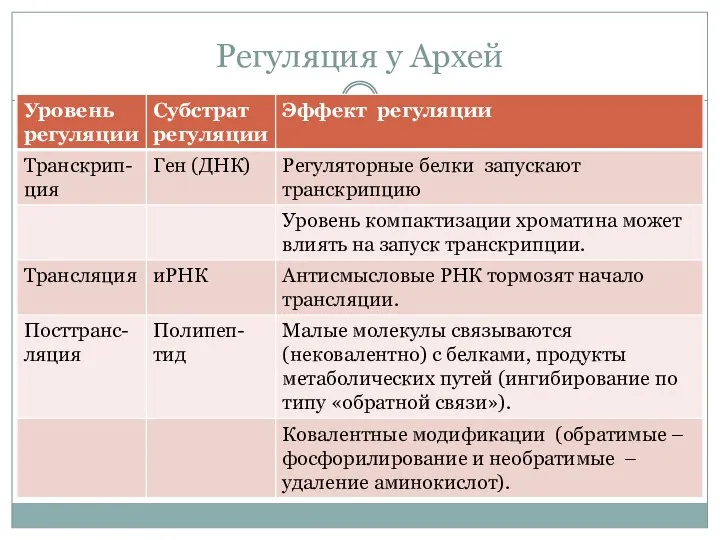
Регуляция у Архей
Регуляция у Архей
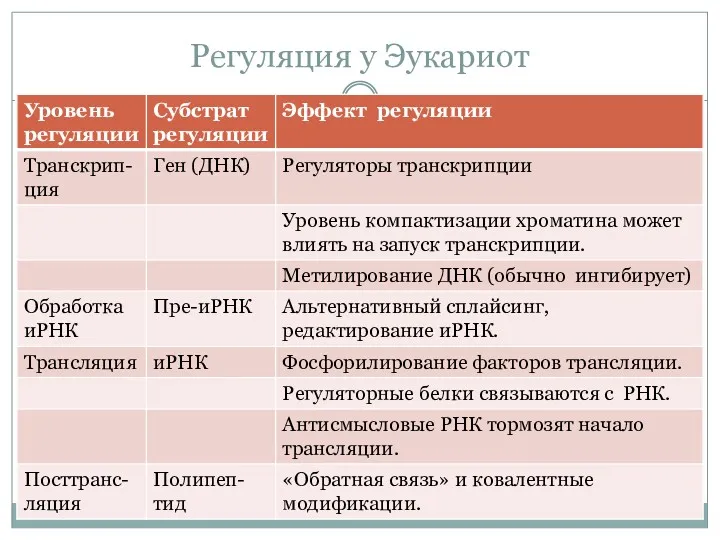
Регуляция у Эукариот
Регуляция у Эукариот

История открытия регуляции
1900г. Эмиль Дюкло обнаружил спобность Aspergillus niger продуцировать фермент
История открытия регуляции
1900г. Эмиль Дюкло обнаружил спобность Aspergillus niger продуцировать фермент

История открытия регуляции
Позже Ледерберг, Моно, Жакоб и Парди обнаружили наличие гена,
История открытия регуляции
Позже Ледерберг, Моно, Жакоб и Парди обнаружили наличие гена,

Регуляция прокариот
Исследования Е. coli показали, что у бактерий существуют ферменты 3
Регуляция прокариот
Исследования Е. coli показали, что у бактерий существуют ферменты 3

Лактозный оперон
При выращивании E.coli на среде с лактозой, содержание лактозы достигает
Лактозный оперон
При выращивании E.coli на среде с лактозой, содержание лактозы достигает
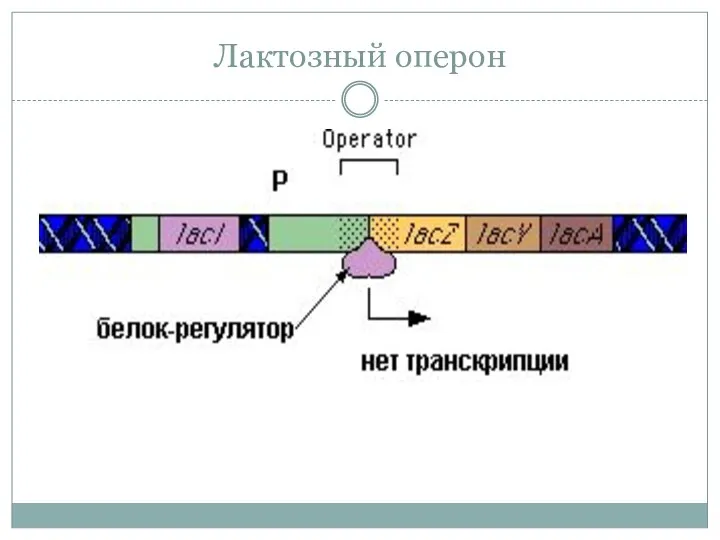
Лактозный оперон
Лактозный оперон
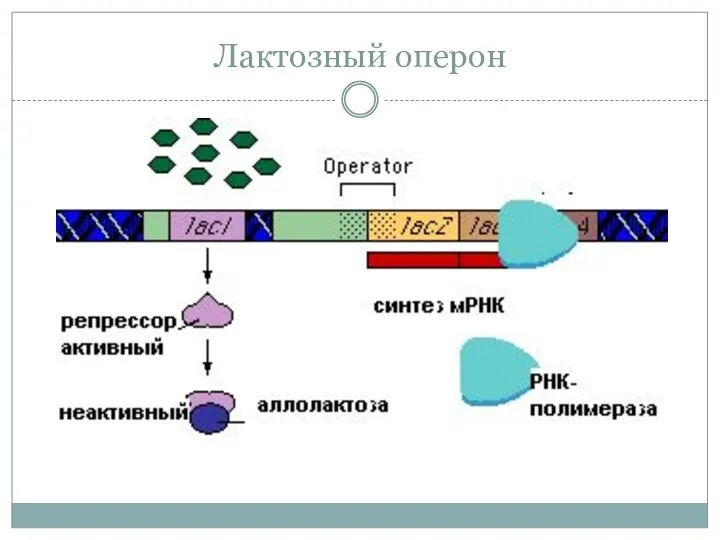
Лактозный оперон
Лактозный оперон
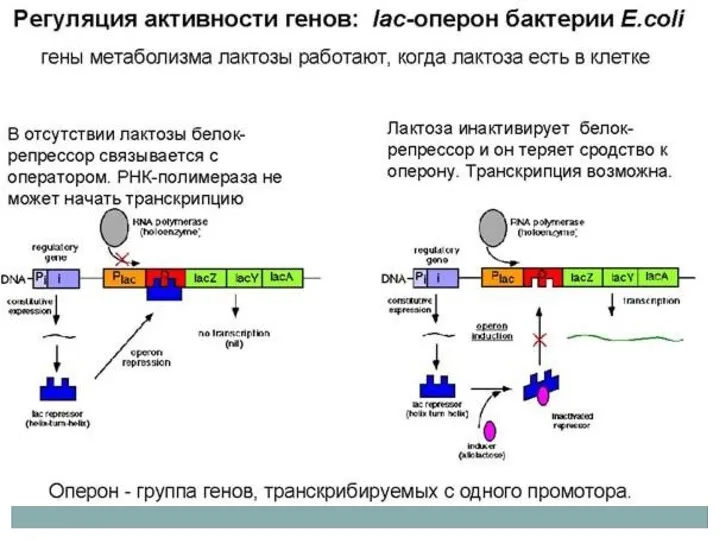
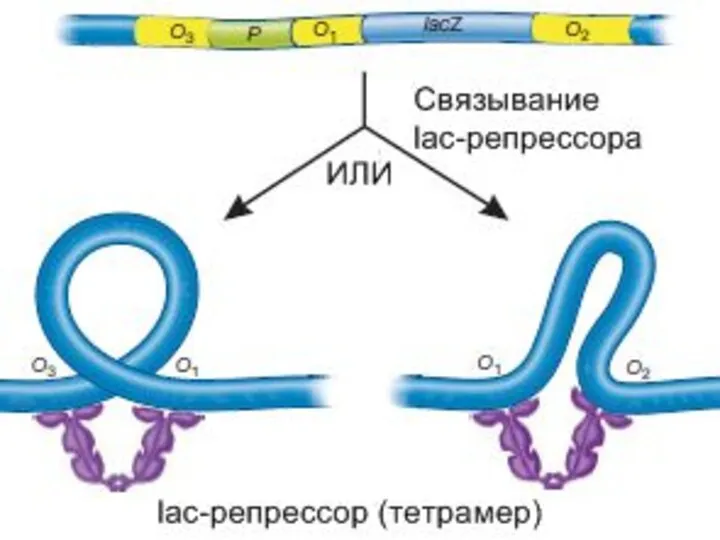
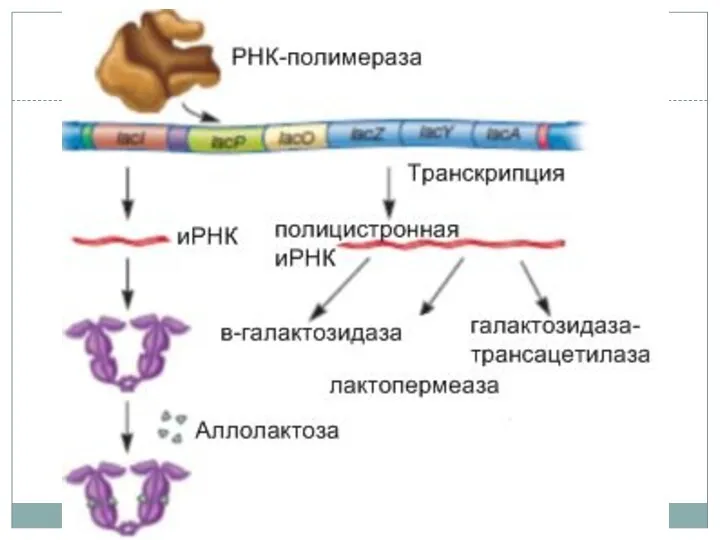
Регуляция лактозного оперона
Регуляция лактозного оперона
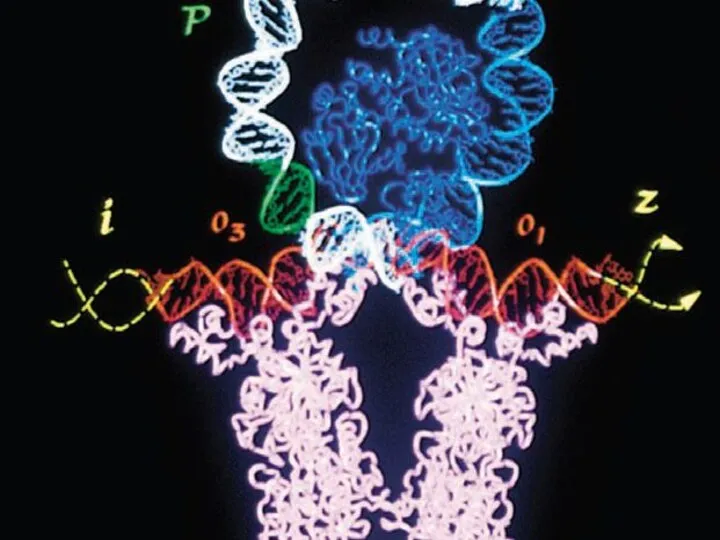
Инициация транскрипции
Инициация транскрипции
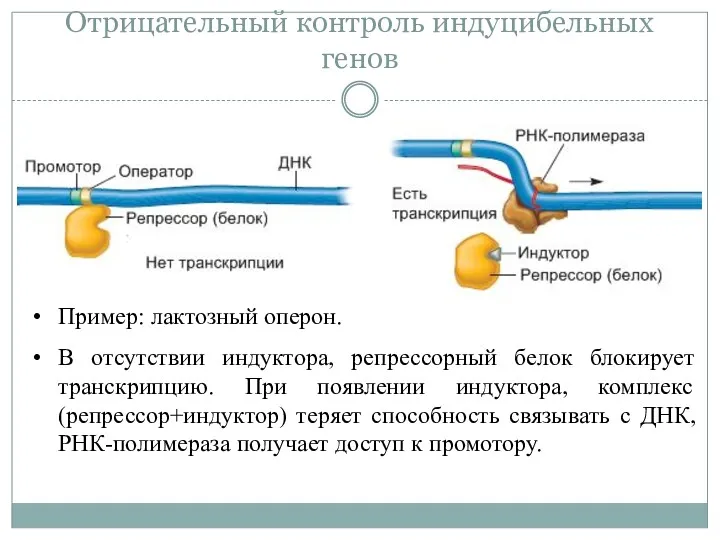
Отрицательный контроль индуцибельных генов
Пример: лактозный оперон.
В отсутствии индуктора, репрессорный белок
Отрицательный контроль индуцибельных генов
Пример: лактозный оперон.
В отсутствии индуктора, репрессорный белок
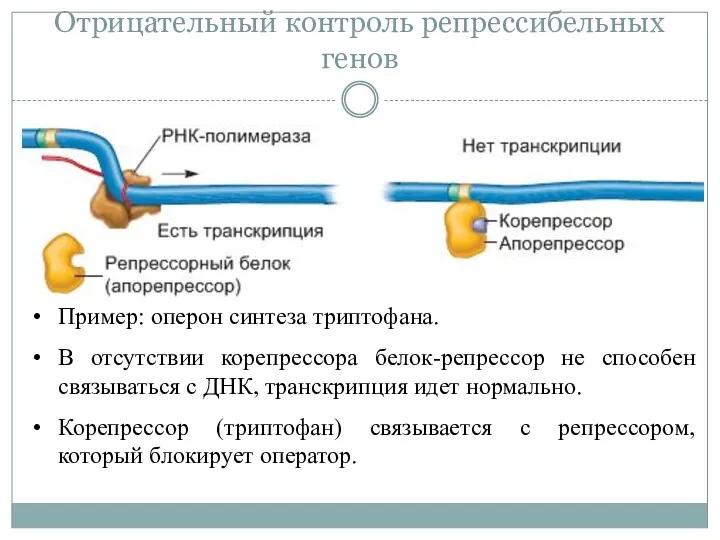
Отрицательный контроль репрессибельных генов
Пример: оперон синтеза триптофана.
В отсутствии корепрессора белок-репрессор
Отрицательный контроль репрессибельных генов
Пример: оперон синтеза триптофана.
В отсутствии корепрессора белок-репрессор
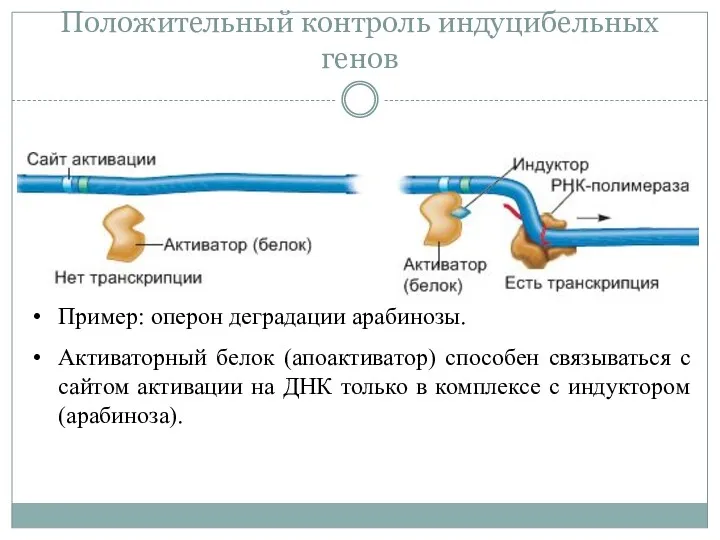
Положительный контроль индуцибельных генов
Пример: оперон деградации арабинозы.
Активаторный белок (апоактиватор) способен
Положительный контроль индуцибельных генов
Пример: оперон деградации арабинозы.
Активаторный белок (апоактиватор) способен
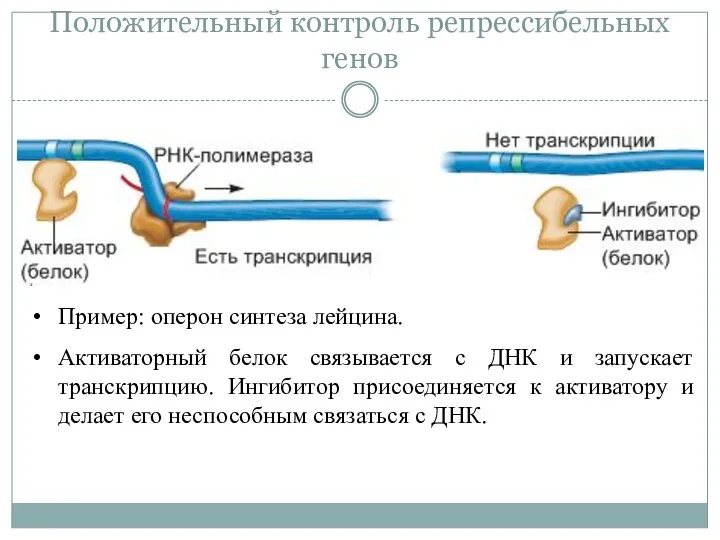
Положительный контроль репрессибельных генов
Пример: оперон синтеза лейцина.
Активаторный белок связывается с
Положительный контроль репрессибельных генов
Пример: оперон синтеза лейцина.
Активаторный белок связывается с
 Анатомия и физиология женских половых органов
Анатомия и физиология женских половых органов Вчення про тканини. Епітелій
Вчення про тканини. Епітелій Антропология, как комплексное учение о человеке. Место человека в животном мире и этапы антропогенеза
Антропология, как комплексное учение о человеке. Место человека в животном мире и этапы антропогенеза Строение стебля
Строение стебля Анатомия внутреннего уха
Анатомия внутреннего уха Физиология центральной нервной системы
Физиология центральной нервной системы Генетически модифицированные организмы (ГМО)
Генетически модифицированные организмы (ГМО) Презентация 1
Презентация 1 Лабораторная работа на тему Приспособленность организмов к их среде обитания
Лабораторная работа на тему Приспособленность организмов к их среде обитания Молодежный инновационный форум Воронежской области
Молодежный инновационный форум Воронежской области Нуклеиновые кислоты и их роль в жизнедеятельности клетки
Нуклеиновые кислоты и их роль в жизнедеятельности клетки презентация к семинару
презентация к семинару Презентация к уроку биологии 6 кл.
Презентация к уроку биологии 6 кл. Вредители и болезни газонных растений и меры борьбы с ними
Вредители и болезни газонных растений и меры борьбы с ними Химерные мыши
Химерные мыши Лишайники. Внешнее строение лишайников
Лишайники. Внешнее строение лишайников Общая характеристика подтипа Позвоночные. Часть1
Общая характеристика подтипа Позвоночные. Часть1 Значення та охорона членистоногих
Значення та охорона членистоногих Животные Омского Прииртышья (насекомые). Занятие № 39
Животные Омского Прииртышья (насекомые). Занятие № 39 Кровеносная система. Строение сердца. Строение сосудов
Кровеносная система. Строение сердца. Строение сосудов Системная биология: предмет, инструменты, экспериментальные методы
Системная биология: предмет, инструменты, экспериментальные методы Моторика (регуляция движений)
Моторика (регуляция движений) История развития генетики. Основные понятия
История развития генетики. Основные понятия Режимы и способы хранения картофеля, плодов, и овощей. Лекция №3-4
Режимы и способы хранения картофеля, плодов, и овощей. Лекция №3-4 Тренировочные задания по биологии в формате ГИА
Тренировочные задания по биологии в формате ГИА Hedera helix
Hedera helix Деление клетки. Митоз. Жизненный цикл клетки
Деление клетки. Митоз. Жизненный цикл клетки Физиология памяти человека
Физиология памяти человека