- Передвижение веществ по растению

Содержание
- 2. I. Роль изучения транспорта веществ: теоретическое значение как одна из проблем физиологии практическое значение взаимосвязь отдельных
- 3. Большую роль в выяснении путей передвижения отдельных питательных веществ сыграл прием кольцевания растений. Этот прием был
- 4. Организация системы транспорта Внутриклеточный Ближний: в пределах одного органа, по неспецифическим тканям, на короткие расстояния. Дальний:
- 5. II. Передвижение элементов минерального питания по растению Назовите акцепторы минеральных веществ Как осуществляется внутриклеточный транспорт Назовите
- 7. Круговорот минеральных веществ в растении. Реутилизация Для растительного организма характерна экономность в использовании питательных веществ, что
- 8. В растении существуют два градиента распределения минеральных веществ: для элементов, подвергающихся повторному использованию, характерен базипетальный градиент
- 9. III. ТРАНСПОРТ ОРГАНИЧЕСКИХ ВЕЩЕСТВ Распределение ассимилятов в растении. Пути передвижения ассимилятов. 3. Механизм транспорта. 4. Регуляция
- 10. 1.Распределение ассимилятов в растении Доноры (источники) ассимилятов - фотосинтезирующие ткани, запасающие ткани (органы). Акцепторы (потребители) –
- 11. Движение по флоэме не имеет определенного направления в отличие от ксилемы, зависит от расположения донора и
- 12. 2. Пути передвижения ассимилятов
- 13. 2.1. Внутриклеточный транспорт Это транспорт ассимилятов из хлоропластов в цитоплазму Крахмал → глюкоза → фруктозодифосфат →
- 14. 2.2. Межклеточный паренхимный транспорт Ближний транспорт может осуществляться двумя путями — по плазмодесмам (симпласту) или по
- 15. Из апопласта и симпласта ассимиляты поступают в сопровождающие (передаточные) клетки (посредники между клетками листовой паренхимы и
- 17. Доказательства флоэмного транспорта 1) Кольцевание, 1679 г. итал. Марчелло Мальпиги. 2) Использование радиоактивных меток 14СО2. 3)
- 18. Структура флоэмы В отличие от ксилемы флоэма представляет собой совокупность живых клеток. Флоэма состоит из нескольких
- 20. Особенности ситовидных трубок протопласты с ограниченной метаболической активностью; система межклеточных контактов посредством ситовидных полей СП; вертикальные
- 21. По мере развития структура СТ претерпевает изменения: распадается ядро; уменьшаются размеры и количество пластид и митохондрий;
- 22. Клетки-спутницы Примыкают к каждой клетке ситовидной трубки. Богаты цитоплазмой Крупное ядро и ядрышко, Многочисленные митохондрии и
- 23. Состав флоэмного экссудата Концентрация флоэмного сока колеблется в пределах от 8 до 20%. На 90% или
- 24. Особенности передвижения по флоэме Высокая скорость - 50—100 см/ч (по симпласту 6 см/час). Большое количество переносимого
- 25. Влияние условий внешней среды Транспорт веществ по флоэме зависит: от температуры. Оптимальная температура 20 и 30
- 26. Механизм флоэмного транспорта Гипотеза «массового тока» Выдвинута в 1930 г. Э. Мюнхом. Ассимиляты транспортируются от источника
- 27. Гипотеза электроосмотического потока Выдвинута в 1979 году Д. Спаннером На каждой ситовидной пластинке возникает электрический потенциал,
- 28. Разгрузка флоэмы В плазмалемме акцепторов работает Н+-помпа. Н+ выкачиваются (апопласт закисляется), что способствует отдаче К+ и
- 29. Непрерывная циркуляция внутренней водной среды – неотъемлемый атрибут жизни Структурные и функциональные взаимосвязи между восходящим и
- 31. Скачать презентацию
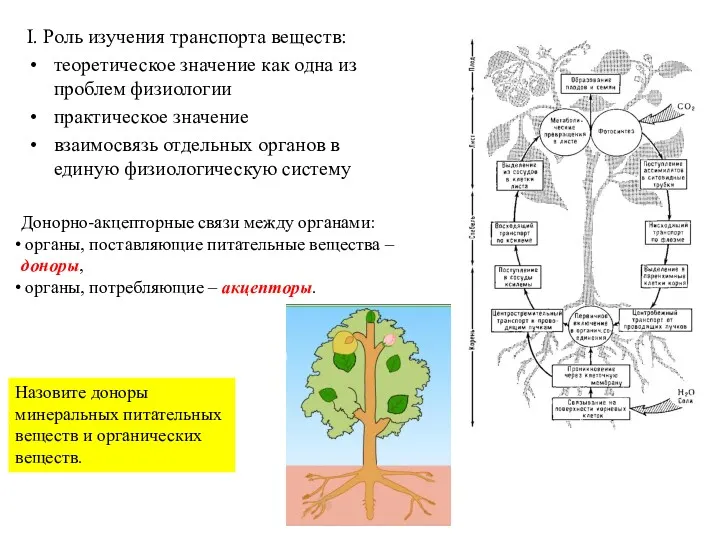
I. Роль изучения транспорта веществ:
теоретическое значение как одна из проблем физиологии
I. Роль изучения транспорта веществ:
теоретическое значение как одна из проблем физиологии

Большую роль в выяснении путей передвижения отдельных питательных веществ сыграл прием
Большую роль в выяснении путей передвижения отдельных питательных веществ сыграл прием

Организация системы транспорта
Внутриклеточный
Ближний: в пределах одного органа, по неспецифическим тканям, на
Организация системы транспорта
Внутриклеточный
Ближний: в пределах одного органа, по неспецифическим тканям, на

II. Передвижение элементов минерального питания по растению
Назовите акцепторы минеральных веществ
Как осуществляется
II. Передвижение элементов минерального питания по растению
Назовите акцепторы минеральных веществ
Как осуществляется

Круговорот минеральных веществ в растении. Реутилизация
Для растительного организма характерна экономность в
Круговорот минеральных веществ в растении. Реутилизация
Для растительного организма характерна экономность в

В растении существуют два градиента распределения минеральных веществ:
для элементов, подвергающихся повторному
В растении существуют два градиента распределения минеральных веществ:
для элементов, подвергающихся повторному

III. ТРАНСПОРТ ОРГАНИЧЕСКИХ ВЕЩЕСТВ
Распределение ассимилятов в растении.
Пути передвижения ассимилятов.
3. Механизм транспорта.
III. ТРАНСПОРТ ОРГАНИЧЕСКИХ ВЕЩЕСТВ
Распределение ассимилятов в растении.
Пути передвижения ассимилятов.
3. Механизм транспорта.

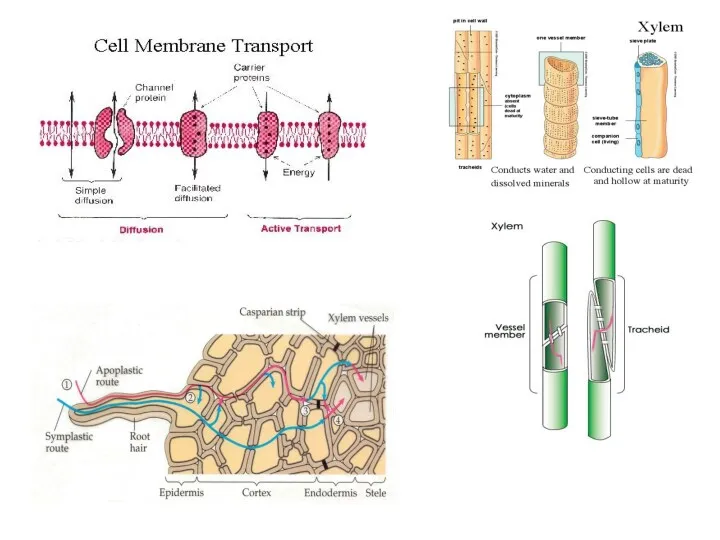
1.Распределение ассимилятов в растении
Доноры (источники) ассимилятов - фотосинтезирующие ткани, запасающие ткани
1.Распределение ассимилятов в растении
Доноры (источники) ассимилятов - фотосинтезирующие ткани, запасающие ткани
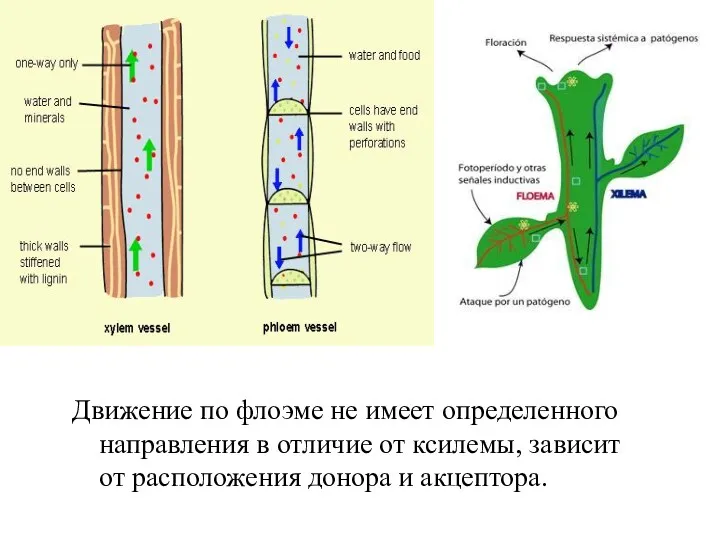
Движение по флоэме не имеет определенного направления в отличие от ксилемы,
Движение по флоэме не имеет определенного направления в отличие от ксилемы,
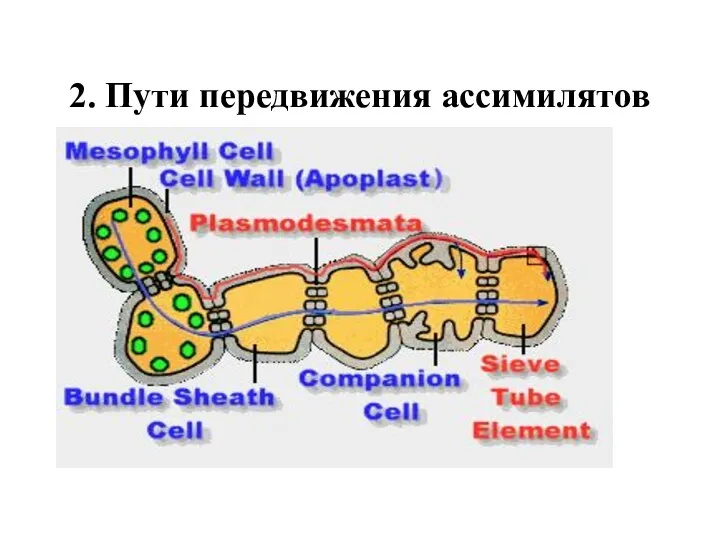
2. Пути передвижения ассимилятов
2. Пути передвижения ассимилятов

2.1. Внутриклеточный транспорт
Это транспорт ассимилятов из хлоропластов в цитоплазму
Крахмал →
2.1. Внутриклеточный транспорт
Это транспорт ассимилятов из хлоропластов в цитоплазму
Крахмал →
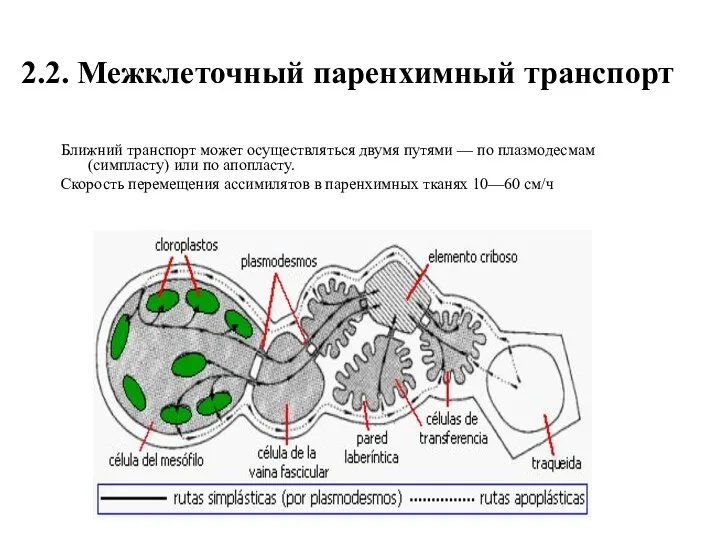
2.2. Межклеточный паренхимный транспорт
Ближний транспорт может осуществляться двумя путями — по
2.2. Межклеточный паренхимный транспорт
Ближний транспорт может осуществляться двумя путями — по
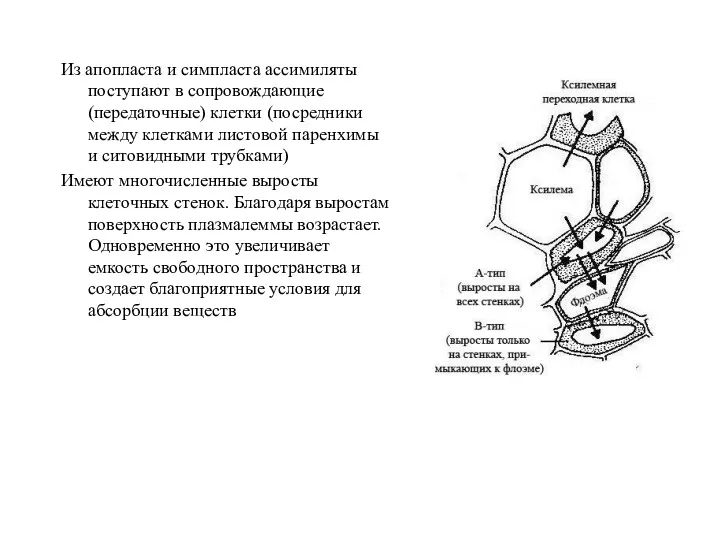
Из апопласта и симпласта ассимиляты поступают в сопровождающие (передаточные) клетки (посредники
Из апопласта и симпласта ассимиляты поступают в сопровождающие (передаточные) клетки (посредники


Доказательства флоэмного транспорта
1) Кольцевание, 1679 г. итал. Марчелло Мальпиги.
2) Использование радиоактивных
Доказательства флоэмного транспорта
1) Кольцевание, 1679 г. итал. Марчелло Мальпиги.
2) Использование радиоактивных
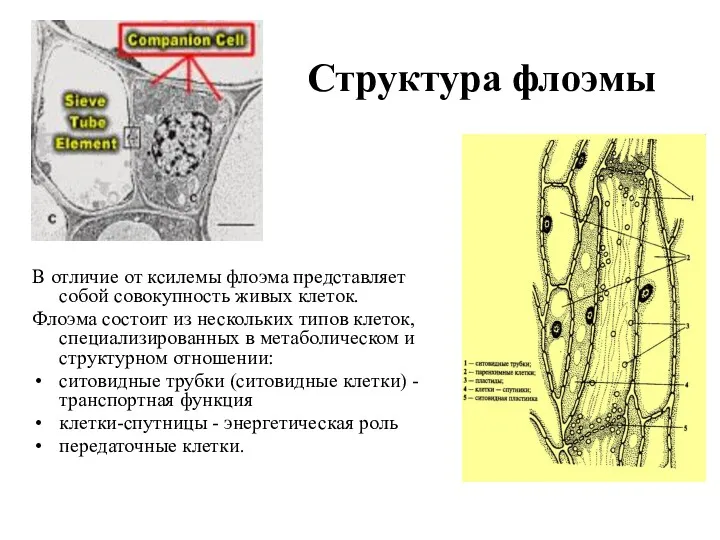
Структура флоэмы
В отличие от ксилемы флоэма представляет собой совокупность живых клеток.
Структура флоэмы
В отличие от ксилемы флоэма представляет собой совокупность живых клеток.
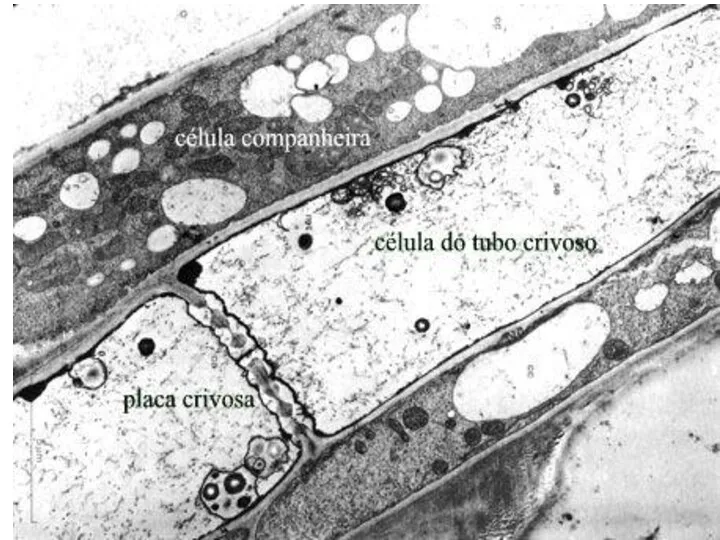
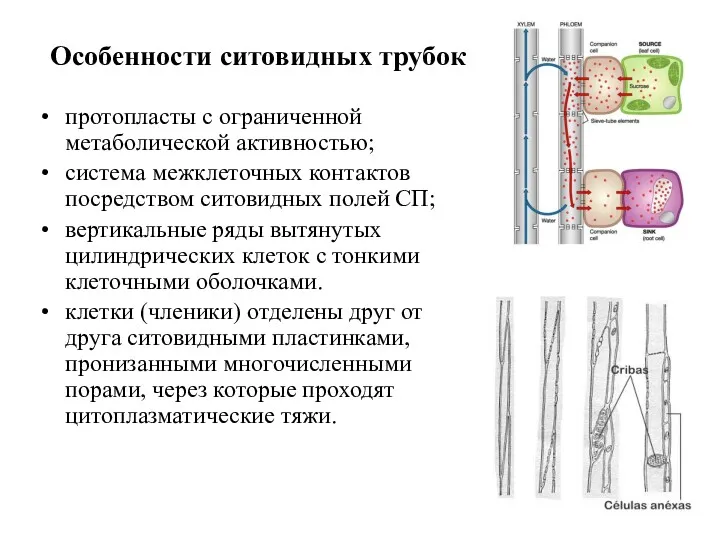
Особенности ситовидных трубок
протопласты с ограниченной метаболической активностью;
система межклеточных контактов посредством ситовидных
Особенности ситовидных трубок
протопласты с ограниченной метаболической активностью;
система межклеточных контактов посредством ситовидных
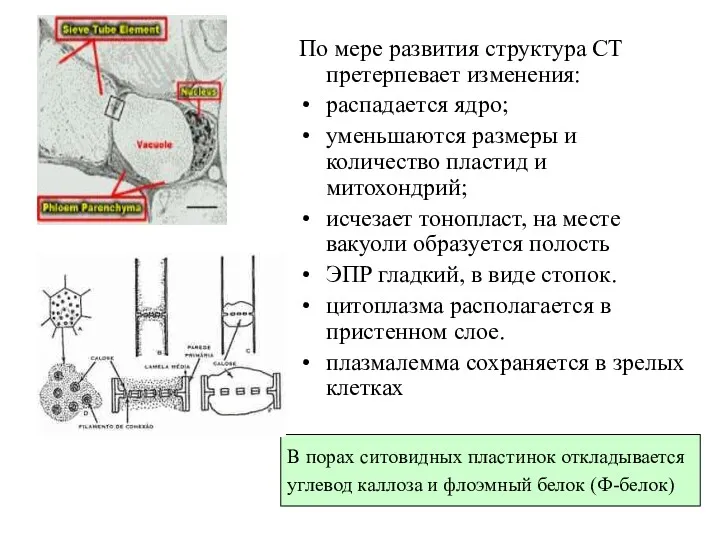
По мере развития структура СТ претерпевает изменения:
распадается ядро;
уменьшаются размеры
По мере развития структура СТ претерпевает изменения:
распадается ядро;
уменьшаются размеры
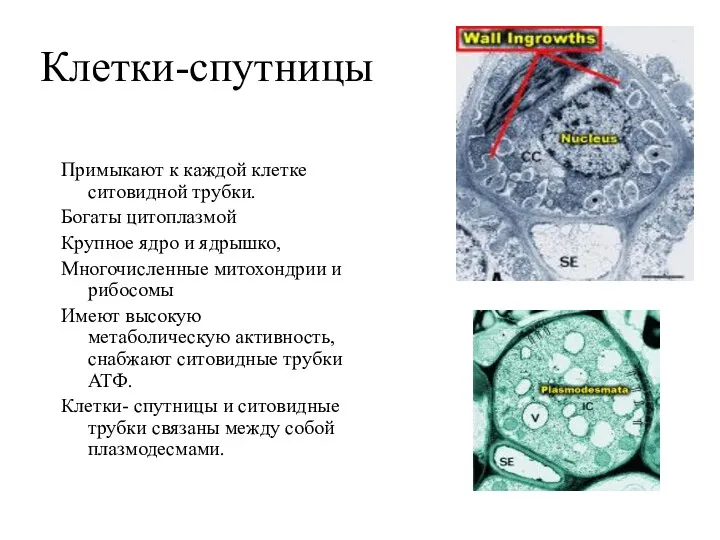
Клетки-спутницы
Примыкают к каждой клетке ситовидной трубки.
Богаты цитоплазмой
Крупное ядро и ядрышко,
Клетки-спутницы
Примыкают к каждой клетке ситовидной трубки.
Богаты цитоплазмой
Крупное ядро и ядрышко,
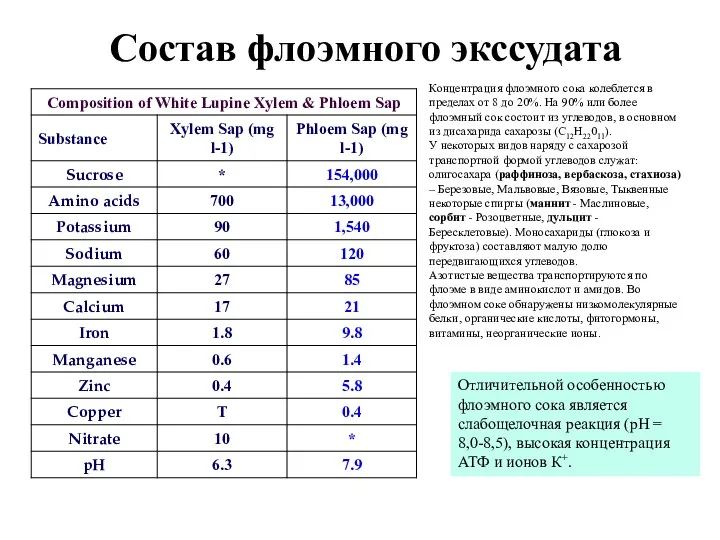
Состав флоэмного экссудата
Концентрация флоэмного сока колеблется в пределах от 8
Состав флоэмного экссудата
Концентрация флоэмного сока колеблется в пределах от 8

Особенности передвижения по флоэме
Высокая скорость - 50—100 см/ч (по симпласту
Особенности передвижения по флоэме
Высокая скорость - 50—100 см/ч (по симпласту

Влияние условий внешней среды
Транспорт веществ по флоэме зависит:
от температуры.
Влияние условий внешней среды
Транспорт веществ по флоэме зависит:
от температуры.
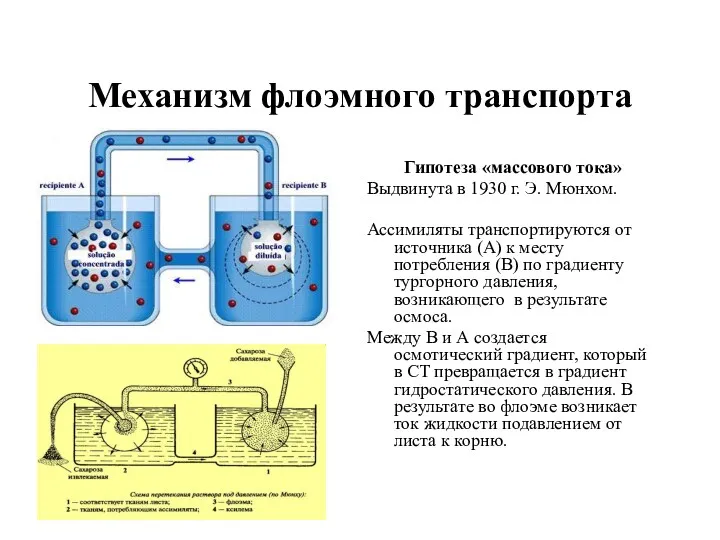
Механизм флоэмного транспорта
Гипотеза «массового тока»
Выдвинута в 1930 г. Э. Мюнхом.
Ассимиляты
Механизм флоэмного транспорта
Гипотеза «массового тока»
Выдвинута в 1930 г. Э. Мюнхом.
Ассимиляты
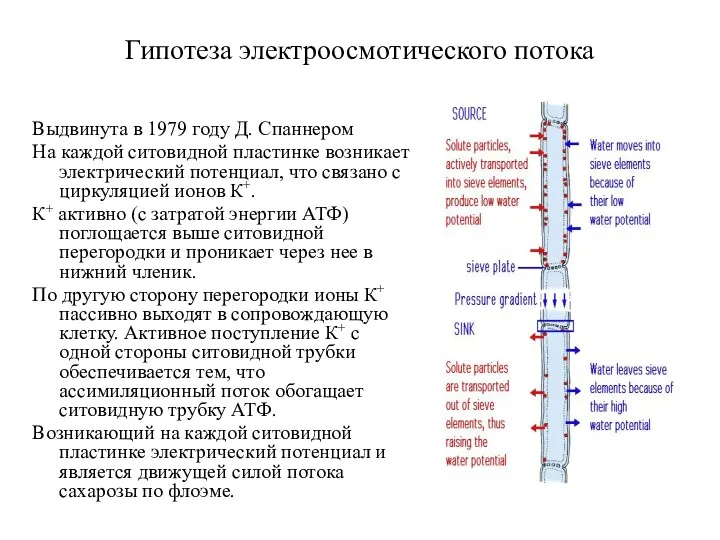
Гипотеза электроосмотического потока
Выдвинута в 1979 году Д. Спаннером
На каждой ситовидной
Гипотеза электроосмотического потока
Выдвинута в 1979 году Д. Спаннером
На каждой ситовидной
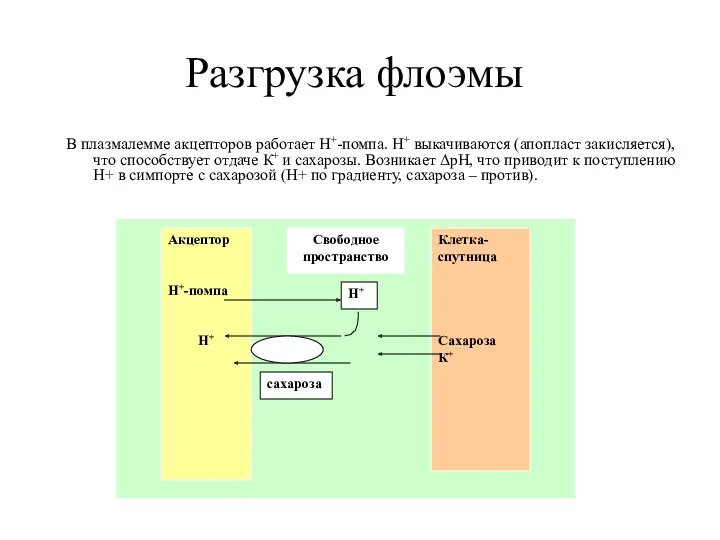
Разгрузка флоэмы
В плазмалемме акцепторов работает Н+-помпа. Н+ выкачиваются (апопласт закисляется), что
Разгрузка флоэмы
В плазмалемме акцепторов работает Н+-помпа. Н+ выкачиваются (апопласт закисляется), что
Непрерывная циркуляция внутренней водной среды – неотъемлемый атрибут жизни
Структурные и функциональные
Непрерывная циркуляция внутренней водной среды – неотъемлемый атрибут жизни
Структурные и функциональные
 Genetically modification organism (GMO)
Genetically modification organism (GMO) Мир рептилий. Первые пресмыкающиеся (рептилии)
Мир рептилий. Первые пресмыкающиеся (рептилии) Тренажёр В мире животных
Тренажёр В мире животных Старение человека и возможность бессмертия
Старение человека и возможность бессмертия Хемосинтез. Хемосинтезирующие организмы
Хемосинтез. Хемосинтезирующие организмы Взаимодействия организмов в экосистемах
Взаимодействия организмов в экосистемах Презентация по теме Антропогенез часть 1
Презентация по теме Антропогенез часть 1 Основные методы селекции и биотехнологии
Основные методы селекции и биотехнологии Органы чувств. Зрение и слух
Органы чувств. Зрение и слух Синапс. Классификация синапсов
Синапс. Классификация синапсов Тип моллюски
Тип моллюски Определение содержания аскорбиновой кислоты (витамина С) в различных сортах яблок
Определение содержания аскорбиновой кислоты (витамина С) в различных сортах яблок Mutation
Mutation Биополимеры. Нуклеиновые кислоты. АТФ
Биополимеры. Нуклеиновые кислоты. АТФ сон и сновидения
сон и сновидения Влияние засоления почвы на процессы роста растений
Влияние засоления почвы на процессы роста растений Отряд хищные
Отряд хищные Соматическая и вегетативная НС
Соматическая и вегетативная НС Биологическая обработка органических отходов , их характеристика и принципы обработки
Биологическая обработка органических отходов , их характеристика и принципы обработки Зимующие птицы
Зимующие птицы Задачи и методы генетики. Подготовка к ЕГЭ
Задачи и методы генетики. Подготовка к ЕГЭ Густота лесных культур
Густота лесных культур Работы Никитина Никиты уч-ся 7 класса
Работы Никитина Никиты уч-ся 7 класса Царство Грибы. Лишайники. Общая характеристика грибов
Царство Грибы. Лишайники. Общая характеристика грибов урок биологии в 8 классе Живая крепость
урок биологии в 8 классе Живая крепость презентация на тему Плоды
презентация на тему Плоды Кругообіг азоту в природі
Кругообіг азоту в природі Конкурс внутреннего и внешнего озеленения образовательных организаций
Конкурс внутреннего и внешнего озеленения образовательных организаций