- Репликация. Эукариоты. Часть 2

Содержание
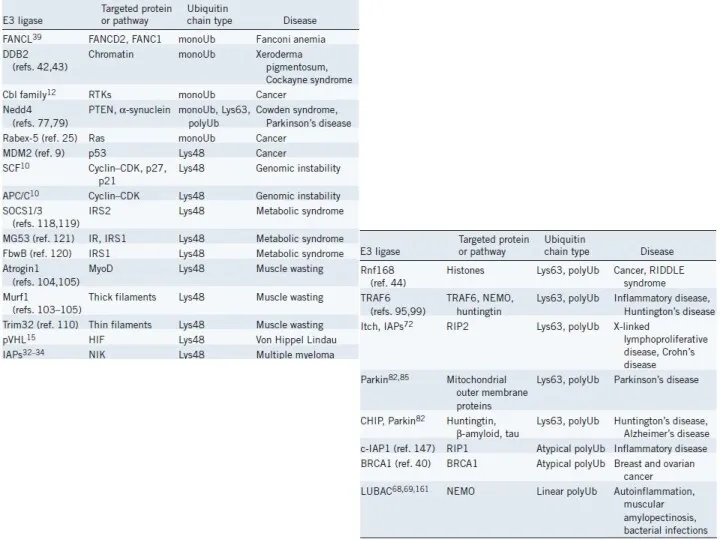
- 3. cdc-гены, сеll division control Реакции обратимого фосфорования: киназы, активируемые циклинами: 2 субъединицы – CDK – каталитическая
- 4. Концентрация циклинов по стадиям клеточного цикла
- 5. КОНТРОЛЬ
- 6. КОНТРОЛЬ
- 7. Cdk1, как и другие киназы, регулирующие клеточный цикл, экспрессируется постоянно, тогда как уровень экспрессии циклинов изменяется
- 8. Фосфорилирование целевых белков “включает или выключает” процесс G1a - R (restriction point) - G1b – CycD/Cdc4,
- 9. Cdk1 регулирует переход M/G1 Активность самой Cdk1 регулируется на двух уровнях: присоединение циклина B, что приводит
- 10. транскрипционный фактор, активирующий работу генов, участвующих в репликации ДНК стадия R, restriction point CKI – Cdk
- 11. Cdk 2 - циклин E Сdk 4 Cdk 6 циклин D Cdk 2,4 и 6 являются
- 12. циклины + циклин-зависимые киназы, Cdks G1/S ATR, ataxia telangiectasia and Rad3-related protein, кратковременная остановка ATM, ataxia
- 13. Cdc2 = Cdk1 (cell division cycle protein 2 homolog or Cyclin-dependent kinase 1) – серин/треониновая портеинкиназа,
- 14. Перенос цитоплазмы из митотической клетки в интерфазную стимулирует вхождение интерфазной клетки в митоз MPF - Mitotic
- 15. В поздней М и ранней G1 фазах Cdc6 отвечает за загрузку неактивной геликазы MCM(2-7) в pre-RC.
- 16. Привлечение Mcm зависит от киназ Cdc6 и Cdt1. Фосфорилирование Mcm приводит к изменению конформации и привлечению
- 17. Mcm2-P и Cdc6-P способны связать cdc45 и RPA уже в поздней G1 или на границе G1/S-фаз.
- 18. Pol alfa/prim загружается также посредством cdc45 и фофорилирования двух больших субъединиц, опосредованное CycE/Cdk2. После загрузки Pol
- 19. После синтеза РНК-ДНК-праймеров с праймированной структурой связывается RFC, который в свою очередь в АТФ-зависимой манере загружает
- 20. После того, как синтез ДНК закончен, ori вновьАТФ-зависимо заполняются комплексом ORC на протяжении частично G2 и
- 21. ORC цикл, исключающий возможность повторной инициации до прохождения митоза
- 22. Фосфорилирование компонентов ORC : препятствует формированию новых pre-RC; активирует-запускает ориджин. 1 2 (до G1/S-checkpoint) (G1/S-checkpoint и
- 24. Начало клеточного цикла – restriction point (R) – - решается вопрос о дальнейшем продвижении по G1-фазе
- 25. ОСНОВНЫЕ ПОСТТРАНСЛЯЦИОННЫЕ МОДИФИКАЦИИ БЕЛКОВ Martina Audagnotto and Matteo Dal Peraro, Comput Struct Biotechnol J. 2017; 15:
- 26. ser thr tyr lys arg his OH NH2 cys SH asp COOH glu ОСНОВНЫЕ ПОСТТРАНСЛЯЦИОННЫЕ МОДИФИКАЦИИ
- 27. ОСНОВНЫЕ ПОСТТРАНСЛЯЦИОННЫЕ МОДИФИКАЦИИ БЕЛКОВ 108222 Phosphorylation 104966 N-linked glycosylation 33291 Acetylation 10295 Methylation 6069 Palmitoylation 5548
- 28. ОСНОВНЫЕ ПОСТТРАНСЛЯЦИОННЫЕ МОДИФИКАЦИИ БЕЛКОВ небольшие химические группы: фосфорилирование, гликозилирование, S-нитрозилирование, метилирование, N-ацетилирование; липиды: пальмитилирование - присоединение
- 29. Убиквитин 76 ао, 8.5 кДа Функции: связывание протеасомой и деградация белков; изменение клеточной локализации; контроль функций
- 30. ОСНОВНЫЕ ПОСТТРАНСЛЯЦИОННЫЕ МОДИФИКАЦИИ БЕЛКОВ
- 32. SUMO Small Ubiquitin-like Modifier ~100 ао, 12 кДа Заболевания: наследственные кардиомиопатии; болезнь Альцгемера; болезнь Паркинсона; болезнь
- 34. Скачать презентацию
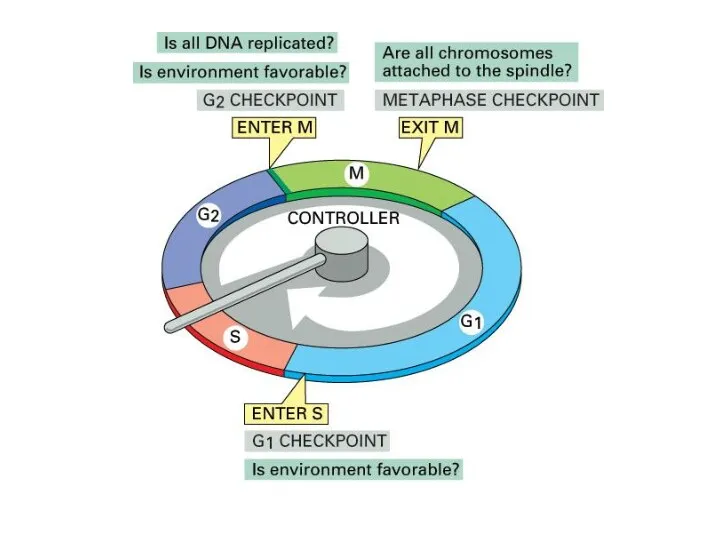

cdc-гены, сеll division control
Реакции обратимого фосфорования:
киназы, активируемые циклинами:
2 субъединицы –
cdc-гены, сеll division control
Реакции обратимого фосфорования:
киназы, активируемые циклинами:
2 субъединицы –
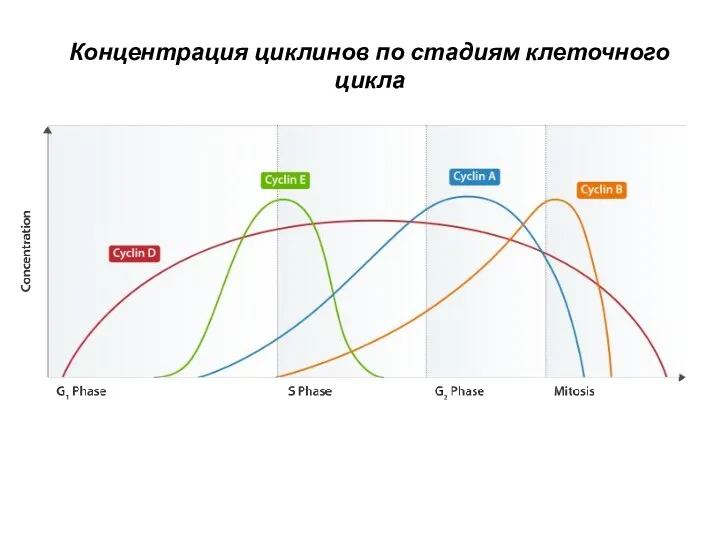
Концентрация циклинов по стадиям клеточного цикла
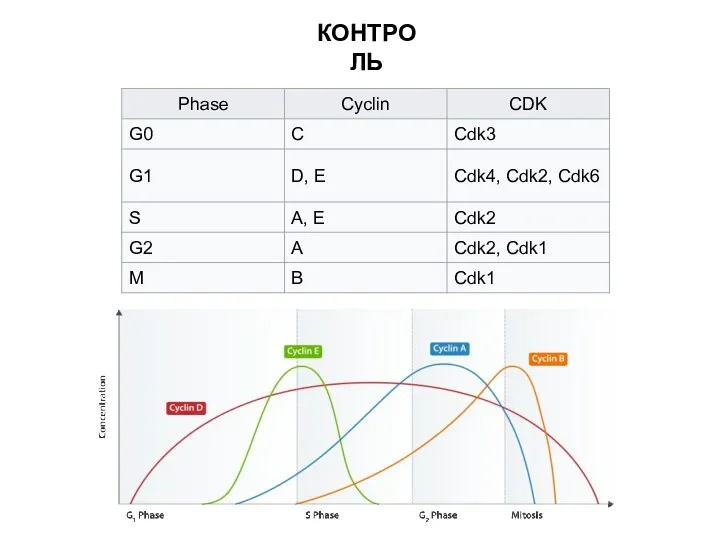
КОНТРОЛЬ
КОНТРОЛЬ
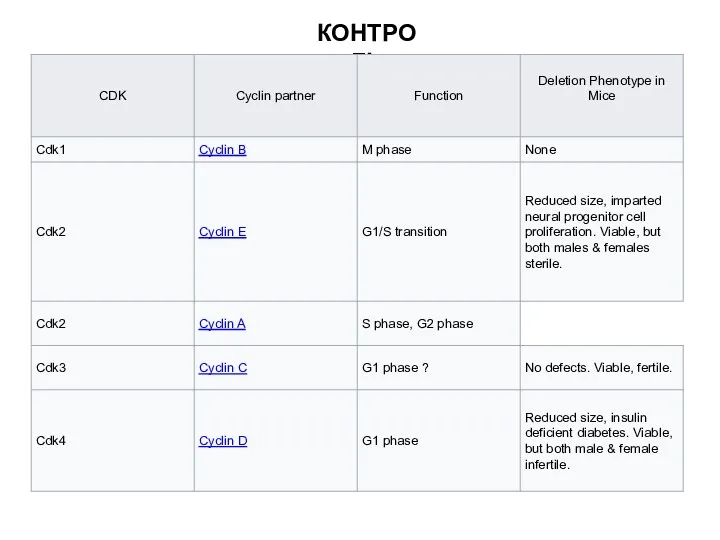
КОНТРОЛЬ
КОНТРОЛЬ
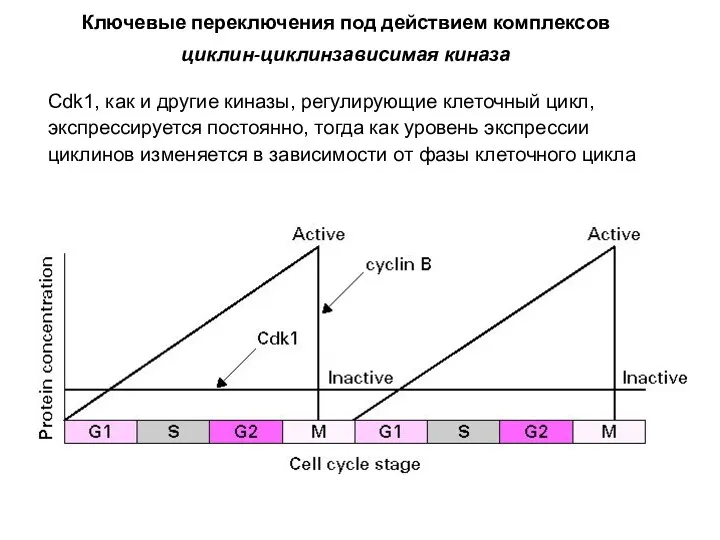
Cdk1, как и другие киназы, регулирующие клеточный цикл,
экспрессируется постоянно, тогда
Cdk1, как и другие киназы, регулирующие клеточный цикл,
экспрессируется постоянно, тогда

Фосфорилирование целевых белков “включает или выключает” процесс
G1a - R (restriction point)
Фосфорилирование целевых белков “включает или выключает” процесс
G1a - R (restriction point)
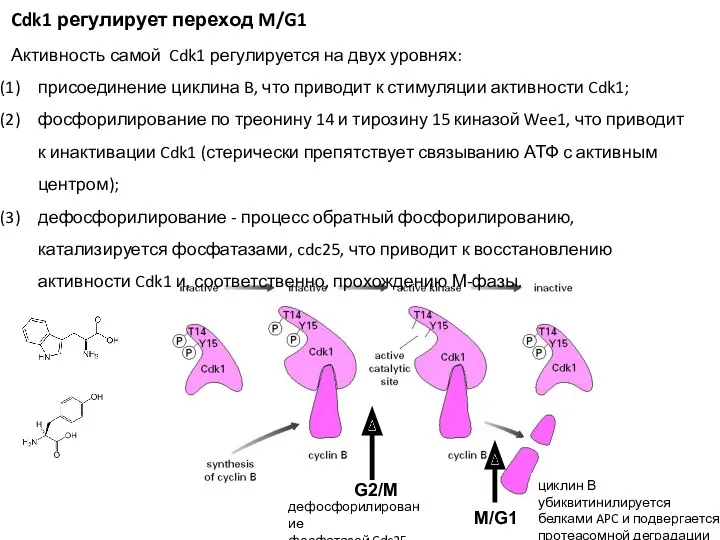
Cdk1 регулирует переход M/G1
Активность самой Cdk1 регулируется на двух уровнях:
присоединение циклина
Cdk1 регулирует переход M/G1
Активность самой Cdk1 регулируется на двух уровнях:
присоединение циклина
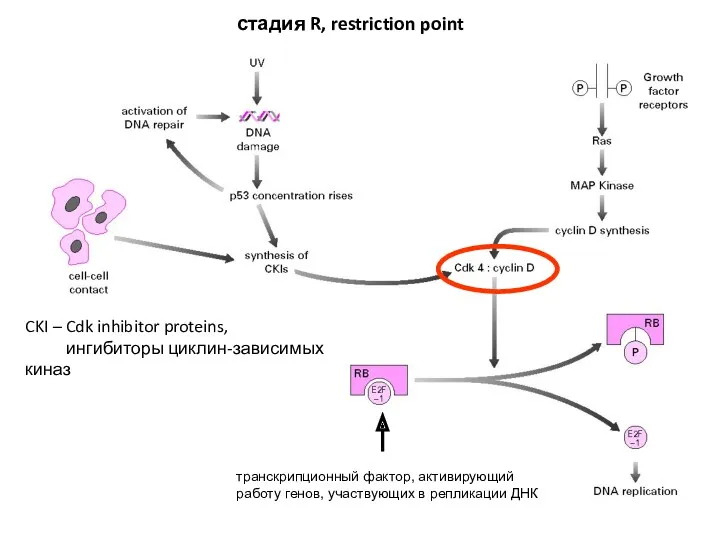
транскрипционный фактор, активирующий
работу генов, участвующих в репликации ДНК
стадия R, restriction point
CKI
транскрипционный фактор, активирующий
работу генов, участвующих в репликации ДНК
стадия R, restriction point
CKI
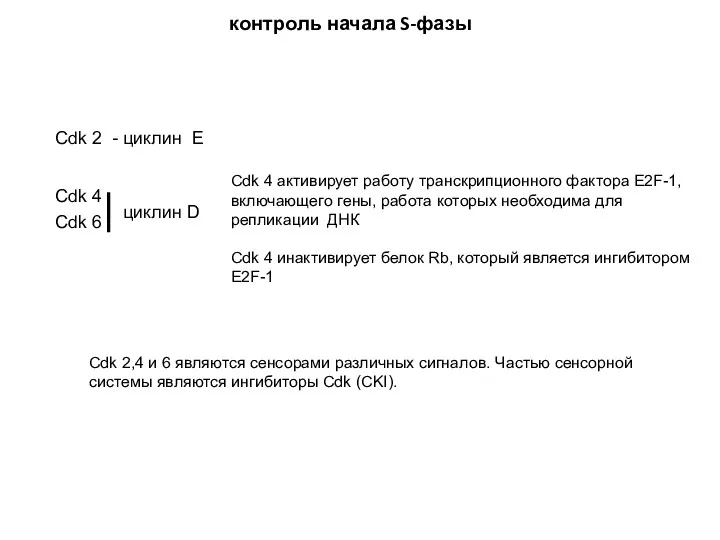
Cdk 2 - циклин E
Сdk 4
Cdk 6
циклин D
Cdk 2,4 и 6
Cdk 2 - циклин E
Сdk 4
Cdk 6
циклин D
Cdk 2,4 и 6
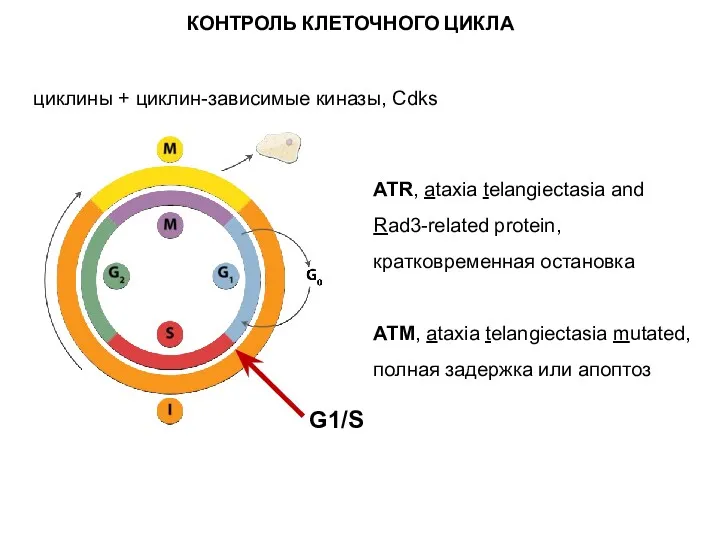
циклины + циклин-зависимые киназы, Cdks
G1/S
ATR, ataxia telangiectasia and Rad3-related protein, кратковременная
циклины + циклин-зависимые киназы, Cdks
G1/S
ATR, ataxia telangiectasia and Rad3-related protein, кратковременная
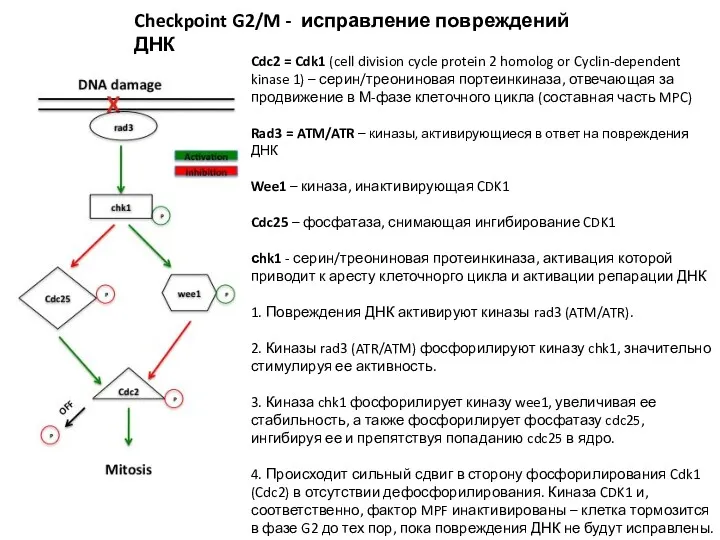
Cdc2 = Cdk1 (cell division cycle protein 2 homolog or Cyclin-dependent
Cdc2 = Cdk1 (cell division cycle protein 2 homolog or Cyclin-dependent
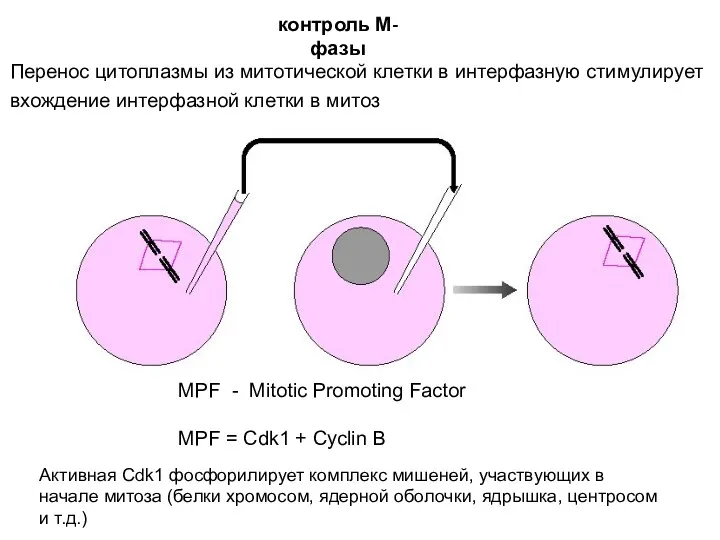
Перенос цитоплазмы из митотической клетки в интерфазную стимулирует
вхождение интерфазной клетки в
Перенос цитоплазмы из митотической клетки в интерфазную стимулирует
вхождение интерфазной клетки в
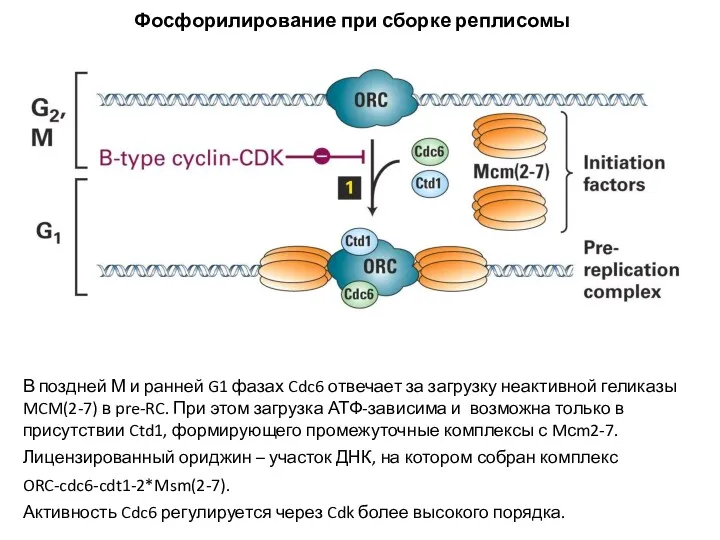
В поздней М и ранней G1 фазах Cdc6 отвечает за загрузку
В поздней М и ранней G1 фазах Cdc6 отвечает за загрузку
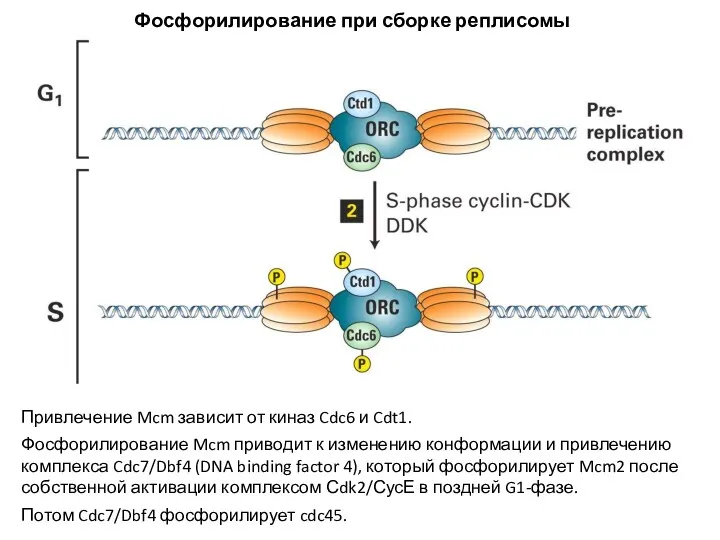
Привлечение Mcm зависит от киназ Cdc6 и Cdt1.
Фосфорилирование Mcm приводит к
Привлечение Mcm зависит от киназ Cdc6 и Cdt1.
Фосфорилирование Mcm приводит к
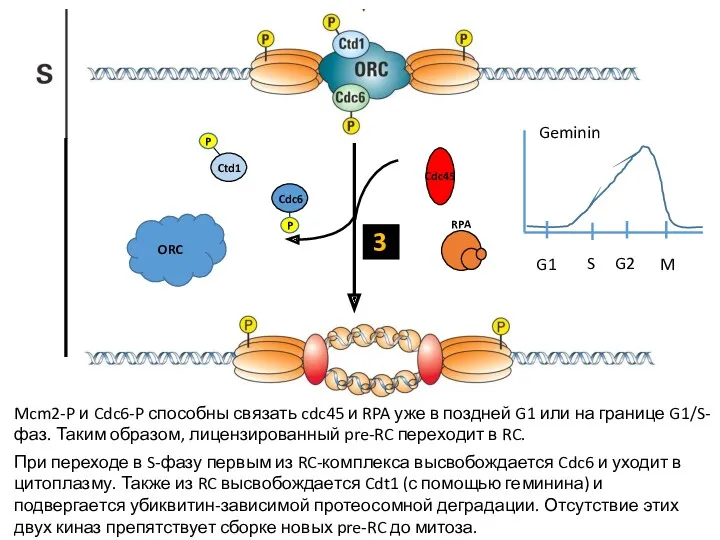
Mcm2-P и Cdc6-P способны связать cdc45 и RPA уже в поздней
Mcm2-P и Cdc6-P способны связать cdc45 и RPA уже в поздней
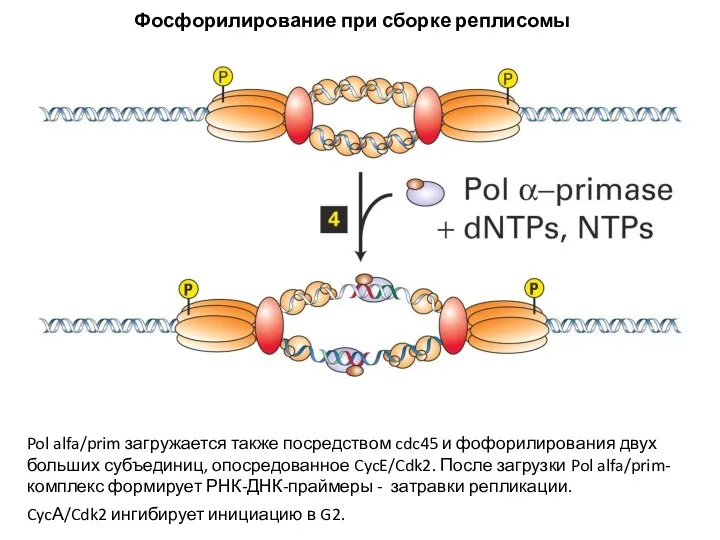
Pol alfa/prim загружается также посредством cdc45 и фофорилирования двух больших субъединиц,
Pol alfa/prim загружается также посредством cdc45 и фофорилирования двух больших субъединиц,
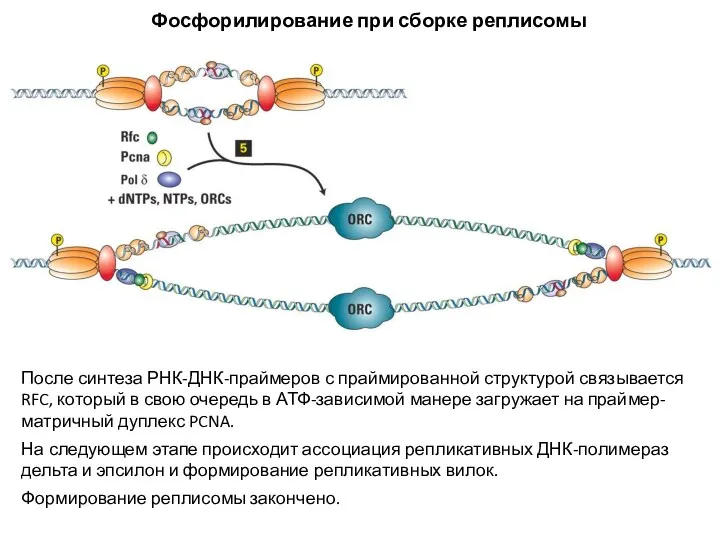
После синтеза РНК-ДНК-праймеров с праймированной структурой связывается RFC, который в свою
После синтеза РНК-ДНК-праймеров с праймированной структурой связывается RFC, который в свою

После того, как синтез ДНК закончен, ori вновьАТФ-зависимо заполняются комплексом ORC
После того, как синтез ДНК закончен, ori вновьАТФ-зависимо заполняются комплексом ORC
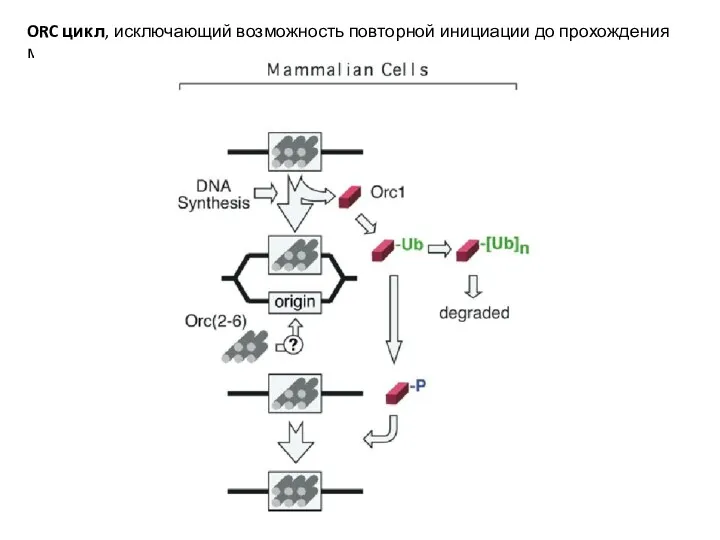
ORC цикл, исключающий возможность повторной инициации до прохождения митоза
ORC цикл, исключающий возможность повторной инициации до прохождения митоза
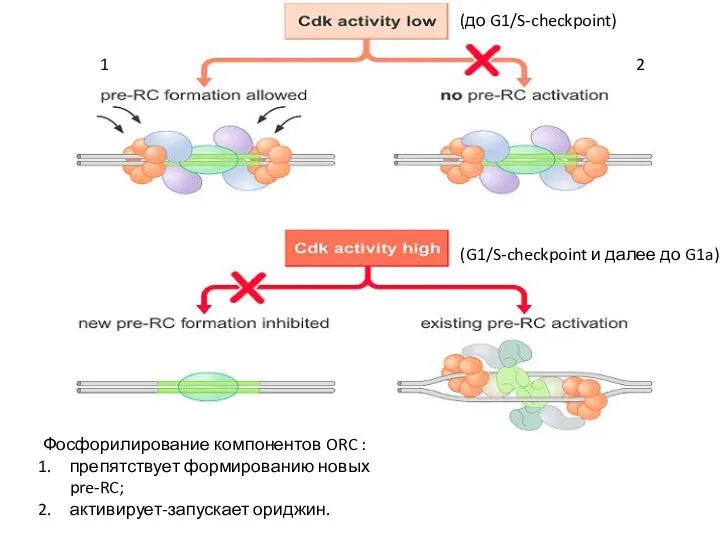
Фосфорилирование компонентов ORC :
препятствует формированию новых pre-RC;
активирует-запускает ориджин.
1
2
(до G1/S-checkpoint)
(G1/S-checkpoint и далее
Фосфорилирование компонентов ORC :
препятствует формированию новых pre-RC;
активирует-запускает ориджин.
1
2
(до G1/S-checkpoint)
(G1/S-checkpoint и далее
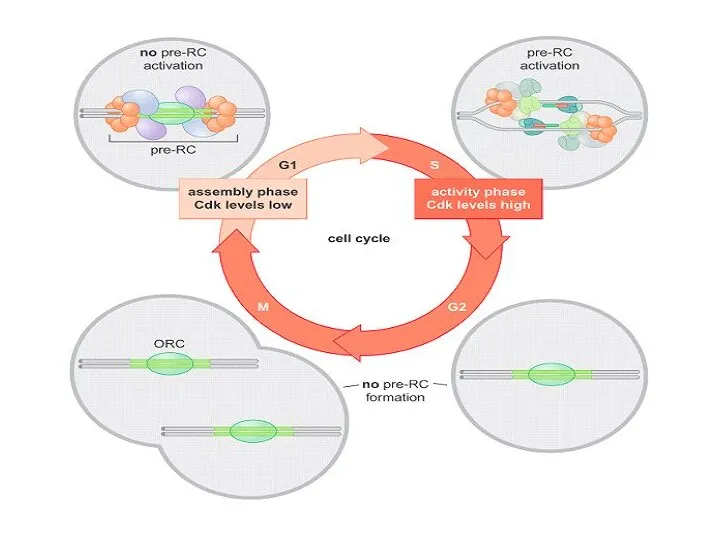

Начало клеточного цикла – restriction point (R) –
- решается вопрос
Начало клеточного цикла – restriction point (R) –
- решается вопрос
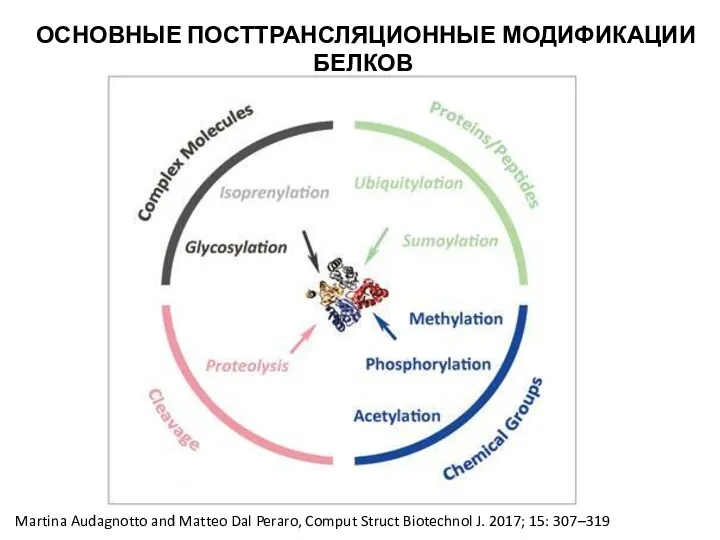
ОСНОВНЫЕ ПОСТТРАНСЛЯЦИОННЫЕ МОДИФИКАЦИИ БЕЛКОВ
Martina Audagnotto and Matteo Dal Peraro, Comput Struct
ОСНОВНЫЕ ПОСТТРАНСЛЯЦИОННЫЕ МОДИФИКАЦИИ БЕЛКОВ
Martina Audagnotto and Matteo Dal Peraro, Comput Struct
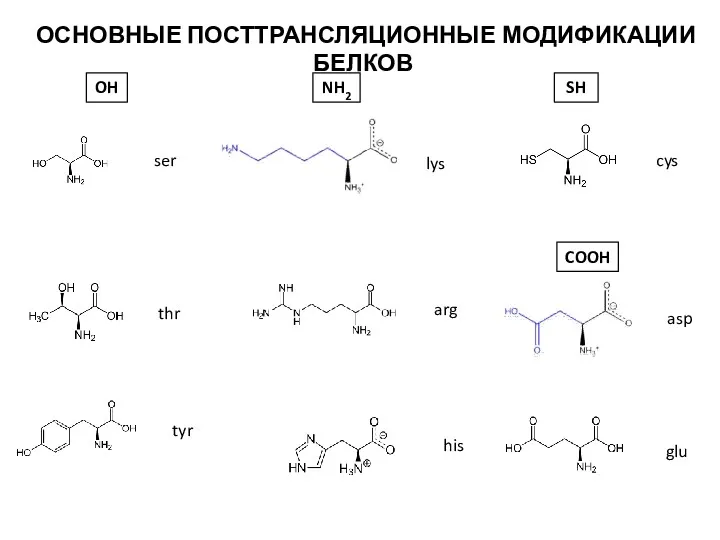
ser
thr
tyr
lys
arg
his
OH
NH2
cys
SH
asp
COOH
glu
ОСНОВНЫЕ ПОСТТРАНСЛЯЦИОННЫЕ МОДИФИКАЦИИ БЕЛКОВ
ser
thr
tyr
lys
arg
his
OH
NH2
cys
SH
asp
COOH
glu
ОСНОВНЫЕ ПОСТТРАНСЛЯЦИОННЫЕ МОДИФИКАЦИИ БЕЛКОВ

ОСНОВНЫЕ ПОСТТРАНСЛЯЦИОННЫЕ МОДИФИКАЦИИ БЕЛКОВ
108222 Phosphorylation
104966 N-linked glycosylation
33291 Acetylation
10295 Methylation
6069 Palmitoylation
5548 Amidation
4808 Citrullination
4104 O-linked glycosylation
3842 Sulfation
3259 Hydroxylation
2983 Ubiquitylation
2062 S-diacylglycerol cysteine
1616 Pyrrolidone Carboxylic Acid
1508 Myristoylation
1344 Sumoylation
1257 Gamma-Carboxyglutamic Acid
1098 Geranyl-geranylation
1012 GPI anchoring
477 S-nitrosylation
440 Deamidation
384 Farnesylation
325 ADP-ribosylation
305 Nitration
259 C-linked
ОСНОВНЫЕ ПОСТТРАНСЛЯЦИОННЫЕ МОДИФИКАЦИИ БЕЛКОВ
108222 Phosphorylation
104966 N-linked glycosylation
33291 Acetylation
10295 Methylation
6069 Palmitoylation
5548 Amidation
4808 Citrullination
4104 O-linked glycosylation
3842 Sulfation
3259 Hydroxylation
2983 Ubiquitylation
2062 S-diacylglycerol cysteine
1616 Pyrrolidone Carboxylic Acid
1508 Myristoylation
1344 Sumoylation
1257 Gamma-Carboxyglutamic Acid
1098 Geranyl-geranylation
1012 GPI anchoring
477 S-nitrosylation
440 Deamidation
384 Farnesylation
325 ADP-ribosylation
305 Nitration
259 C-linked

ОСНОВНЫЕ ПОСТТРАНСЛЯЦИОННЫЕ МОДИФИКАЦИИ БЕЛКОВ
небольшие химические группы:
фосфорилирование, гликозилирование, S-нитрозилирование, метилирование, N-ацетилирование;
липиды:
пальмитилирование
ОСНОВНЫЕ ПОСТТРАНСЛЯЦИОННЫЕ МОДИФИКАЦИИ БЕЛКОВ
небольшие химические группы:
фосфорилирование, гликозилирование, S-нитрозилирование, метилирование, N-ацетилирование;
липиды:
пальмитилирование
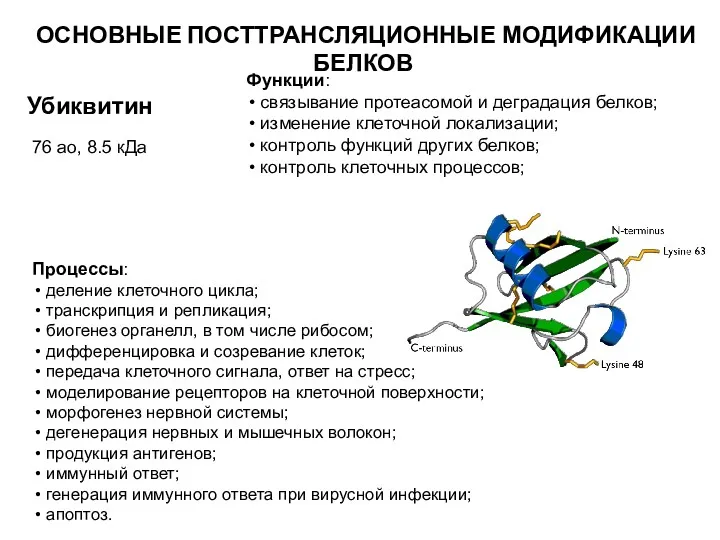
Убиквитин
76 ао, 8.5 кДа
Функции:
связывание протеасомой и деградация белков;
изменение клеточной локализации;
контроль функций
Убиквитин
76 ао, 8.5 кДа
Функции:
связывание протеасомой и деградация белков;
изменение клеточной локализации;
контроль функций
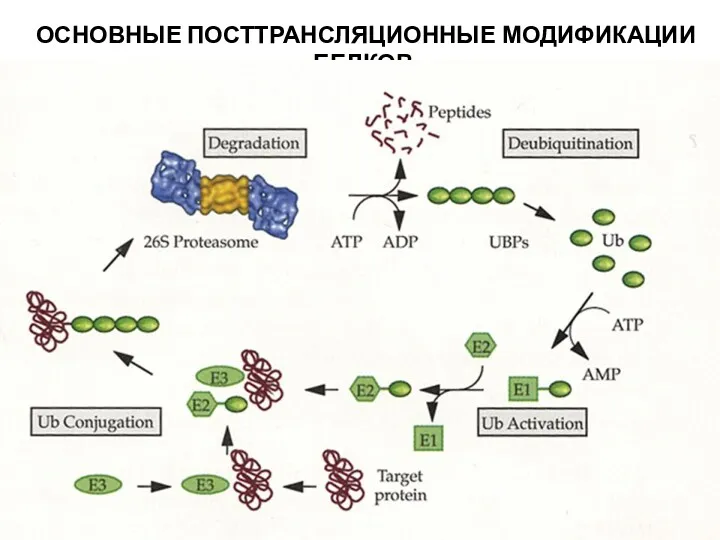
ОСНОВНЫЕ ПОСТТРАНСЛЯЦИОННЫЕ МОДИФИКАЦИИ БЕЛКОВ
ОСНОВНЫЕ ПОСТТРАНСЛЯЦИОННЫЕ МОДИФИКАЦИИ БЕЛКОВ
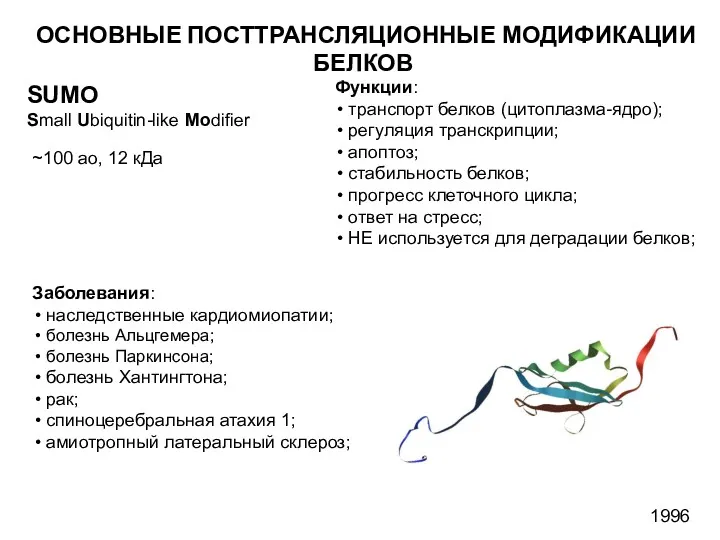
SUMO
Small Ubiquitin-like Modifier
~100 ао, 12 кДа
Заболевания:
наследственные кардиомиопатии;
болезнь Альцгемера;
болезнь Паркинсона;
болезнь Хантингтона;
рак;
спиноцеребральная атахия
SUMO
Small Ubiquitin-like Modifier
~100 ао, 12 кДа
Заболевания:
наследственные кардиомиопатии;
болезнь Альцгемера;
болезнь Паркинсона;
болезнь Хантингтона;
рак;
спиноцеребральная атахия
 Методы исследования биологии
Методы исследования биологии Ферменты в генетической инженерии. Тема 2
Ферменты в генетической инженерии. Тема 2 Внешнее строение листа
Внешнее строение листа Клиническая физиология КЩС
Клиническая физиология КЩС Происхождение человека
Происхождение человека Цветок – орган генеративного размножения
Цветок – орган генеративного размножения General Musculoskeletal Screening: Upper Extremities
General Musculoskeletal Screening: Upper Extremities Эволюционные теории и взгляды
Эволюционные теории и взгляды Систематика растений
Систематика растений Дуб - царь деревьев
Дуб - царь деревьев Эпизоотологическое и эпидемиологическое значение грызунов. Дератизация
Эпизоотологическое и эпидемиологическое значение грызунов. Дератизация Значение водорослей в природе и жизни человека
Значение водорослей в природе и жизни человека презентация к уроку по теме: Цветок. Строение и значение цветка
презентация к уроку по теме: Цветок. Строение и значение цветка Антропогенез. Гипотезы возникновения человека. Сходство и различия человека и животных. Часть 4
Антропогенез. Гипотезы возникновения человека. Сходство и различия человека и животных. Часть 4 Живі фільтри
Живі фільтри Слух. Среднее и внутреннее ухо. Функции среднего уха. Строение улитки
Слух. Среднее и внутреннее ухо. Функции среднего уха. Строение улитки Анатомия и физиология ЦНС. Свойства и принципы функционирования нервных центров. Строение СМ. Цереброспинальная жидкость
Анатомия и физиология ЦНС. Свойства и принципы функционирования нервных центров. Строение СМ. Цереброспинальная жидкость Ферменты,структура и механизм действия. Классификация и номенклатура. (Лекция 4)
Ферменты,структура и механизм действия. Классификация и номенклатура. (Лекция 4) Практика по общей биологии
Практика по общей биологии Простейшие – эукариотические одноклеточные микроорганизмы
Простейшие – эукариотические одноклеточные микроорганизмы Главные направления эволюции органического мира
Главные направления эволюции органического мира Желудочная секреция
Желудочная секреция Строение, химический состав и жизнедеятельность растительной клетки
Строение, химический состав и жизнедеятельность растительной клетки презентация по биологии Экологическая задача Ч.Дарвина
презентация по биологии Экологическая задача Ч.Дарвина Nervové mechanismy regulace dýchání
Nervové mechanismy regulace dýchání Минеральные вещества
Минеральные вещества Что такое хвоинки
Что такое хвоинки Глюконеогенез. Регуляция гликолиза и глюконеогенеза. Цикл кори. Пентозофосфатный путь превращения глюкозы. Спиртовое брожение
Глюконеогенез. Регуляция гликолиза и глюконеогенеза. Цикл кори. Пентозофосфатный путь превращения глюкозы. Спиртовое брожение