- Роль ретикулярной формации в интегративной деятельности мозга

Содержание
- 2. РОЛЬ РЕТИКУЛЯРНОЙ ФОРМАЦИИ (РФ) В ИНТЕГРАТИВНОЙ ДЕЯТЕЛЬНОСТИ МОЗГА Мэгун и Моруцци (1949) - РФ действительно оказывает
- 3. Неспецифическая проекционная таламо-кортикальная система составляющая неспецифическую проекционную таламо-кортикальную систему (НПТКС). Мэгун и Моруцци предположили, что одним
- 4. СОН Благослови, господь, изобретшего сон. М. Сервантес де Сааведра. "Дон Кихот" Сон — это чудо матери
- 5. Сон - активное физиологическое состояние, которое характеризуется потерей активных психических связей субъекта с окружающим его миром.
- 6. Выделяют несколько видов сна: физиологический периодический ежесуточный сон нефизиологический сон: - наркотический сон, вызываемый различными химическими
- 7. СТРУКТУРА СНА "Медленный" сон: 1. Стадия тета-волн. 2. Стадия «сонных веретен» и «К-комплексов». 3. Стадия дельта-волн
- 8. "Медленный" сон: Очень поверхностный сон. Высокоамплитудные медленные тета-волны (3-7 Гц). Поверхностный сон. Веретенообразные всплески бета-ритма (сонные
- 9. СТАДИИ СНА 1 стадия 2 стадия БДГ сон 4 стадия Стадии медленного сна
- 10. "Быстрый" сон (БДГ- сон) Основной отличительной особенностью "быстрого" сна является наличие быстрых движений глазных яблок (фаза
- 11. ТЕОРИИ СНА Переход от бодрствования ко сну предполагает два возможных пути: Пассивный, в результате утомления ЦНС
- 12. 1. Гипотеза гипнотоксинов или гуморальная теория Лежандр и Пьерон (1910) Сон обусловлен накоплением в крови особых
- 13. Традиционные обсуждаемые нейрохимические агенты, имеющие значение в организации цикла "сон- бодрствование": Фаза медленного сна: ГАМК, серотонин,
- 14. Однако многие факты и, в частности, то, что можно преодолеть сон интересным занятием или то, что
- 15. 2. Теория центров сна Фон Экономо (1930); Гесс (1933) передний гипоталамус
- 16. Однако в дальнейшем было показано, что состояние сна можно было получить при электростимуляции в определенном ритме
- 17. 3. Теория условного торможения И.П. Павлов даже сделал вывод о том, что условное торможение и сон
- 18. 4. Теории деафферентации сенсорных систем Бремер, конец 1930-х годов Для бодрствования необходим хотя бы минимальный уровень
- 19. Однако даже в изолированном переднем мозге появляются ритмичные колебания, характерные для цикла сна/бодрствования. Кроме того, лишение
- 20. 5. Теория неспецифических регуляторов сна Активирующие структуры: ВАРС Неспецифическая и специфическая таламическая система Ядра заднего гипоталамуса
- 21. Однако в ретикулярной формации существуют не только центры бодрствования, но и сна. Каудальные отделы оказывают тормозное
- 22. 6.Серотонинергическая теория М. Жуве - конец 60-х годов В верхних отделах ствола мозга есть две области
- 23. Однако сейчас доказано, что нейроны ядер шва наиболее активны при бодрствовании и серотонин большую роль не
- 24. ПАМЯТЬ
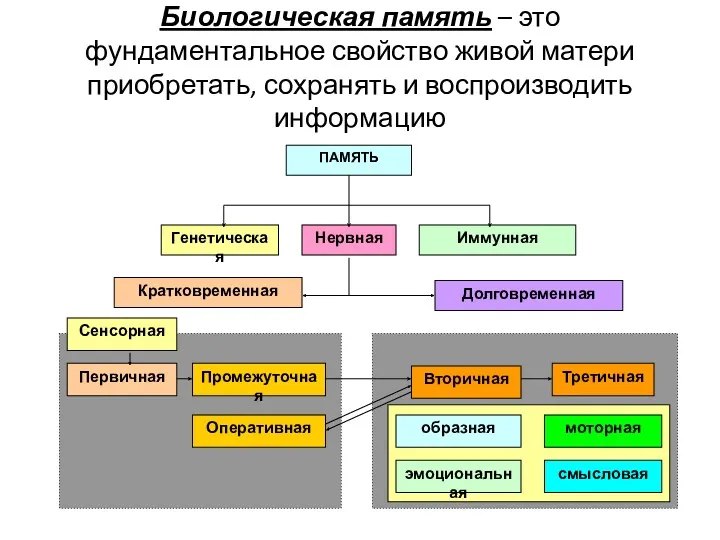
- 25. Биологическая память – это фундаментальное свойство живой матери приобретать, сохранять и воспроизводить информацию
- 26. Генетическая - эволюционно наиболее ранняя форма памяти. Материальным носителем является геном - генетический аппарат клетки, ДНК
- 27. НЕРВНАЯ ПАМЯТЬ Нервная память - совокупность процессов фиксации, хранения и извлечения информации, получаемой организмом в течение
- 28. ВРЕМЕННАЯ ОРГАНИЗАЦИЯ ПАМЯТИ Концепция временной организации памяти принадлежит канадскому психологу Д. Хеббу (D.Hebb). Память принято делить
- 29. Классификация видов долговременной памяти, предложенная И.С.Бериташвили (1968) образная (декларативная) - сохранение и репродукция однажды воспринятого важного
- 30. Фиксация образов, явлений происходит в три этапа: формирование энграммы, сортировка и выделение новой информации, долговременное хранение
- 31. МЕХАНИЗМЫ ПАМЯТИ Кратковременная, и долговременная памяти имеют разную природу и регулируются разными механизмами, Механизмы кратковременной, и
- 32. МЕХАНИЗМЫ КРАТКОВРЕМЕННОЙ ПАМЯТИ По современным представлениям в основе кратковременной памяти лежат два взаимно дополняющих друг друга
- 33. Механизм кратковременной синаптической пластичности - изменение амплитуды постсинаптического потенциала за счет либо увеличения либо уменьшения величины
- 34. МЕХАНИЗМЫ ДОЛГОВРЕМЕННОЙ ПАМЯТИ Механизм долговременной синаптической пластичности – изменение амплитуды постсинаптического потенциала за счет постсинаптических механизмов:
- 35. Основной причиной является изменение концентрации внутриклеточного кальция в постсинаптической клетке: Через NMDAзависимые каналы. Вход Са2+ зависит
- 36. Модель формирования долговременной памяти (Анохин К.В., 1997): Активация кальцием внутриклеточных протеинкиназ (кальций-кальмодулинзависимые киназы – СаМКII, цАМФ-зависимая
- 37. ЛОКАЛИЗАЦИЯ СЛЕДОВ ПАМЯТИ Большинство нейрофизиологов полагает, что запечатление и извлечение энграмм идет при участии тысяч нейронов
- 38. МОТИВАЦИИ
- 39. Мотивация - стремление к удовлетворению потребностей (влечения, побуждения, драйвы), имеющая эмоциональную окраску. Различают: низшие (биологические) высшие
- 40. ПЕРИФЕРИЧЕСКИЕ И ЦЕНТРАЛЬНЫЕ МЕХАНИЗМЫ МОТИВАЦИЙ Причиной мотиваций являются преимущественно раздражители внутренней среды – это изменения тех
- 41. Мотивационные центры: 1. Центр голода - вентромедиальное ядро и латеральный ГПТ – нейроны которых избирательно чувствительны
- 42. Мотивации – это особое интегрированное состояние головного мозга, в которое каждая его структура вносит свой специфический
- 43. ЭМОЦИИ
- 44. Эмоция (волнение, возбуждение) – субъективное состояние человека и животных, которое возникает в ответ на внешние и
- 45. Функции эмоций: Эмоции являются одним из главных механизмов регуляции психической деятельности, направленной на удовлетворение потребностей. Эмоции
- 46. ТЕОРИИ ВОЗНИКНОВЕНИЯ ЭМОЦИЙ 1. Периферическая теория (Джеймс – Ланге, 1885). Эмоции рассматриваются как следствие изменений в
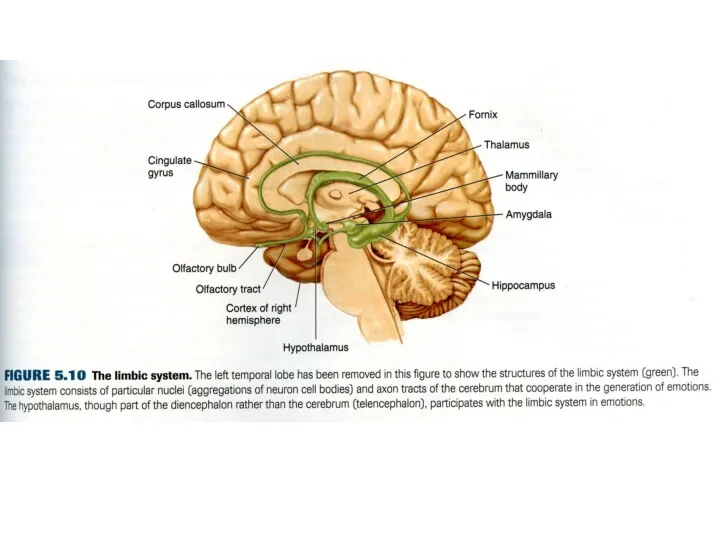
- 47. 3. Центральная лимбическая теория Пейпс (Papez), 1937. Ведущая роль в формировании эмоций отдается лимбическим структурам. Гиппокамп
- 48. 4. Активационная теория (Линдсли, 1958). Подчеркивает роль РФ в формировании эмоций разного знака. Вызывают эмоции нейроны
- 50. Симонов различает четыре состояния эмоционального напряжения (СН):
- 51. Периферические компоненты эмоций представлены: произвольно управляемыми проявлениями, такими, как движения, мимика, речь, дыхание, и неуправляемыми (вегетативными):
- 53. Скачать презентацию
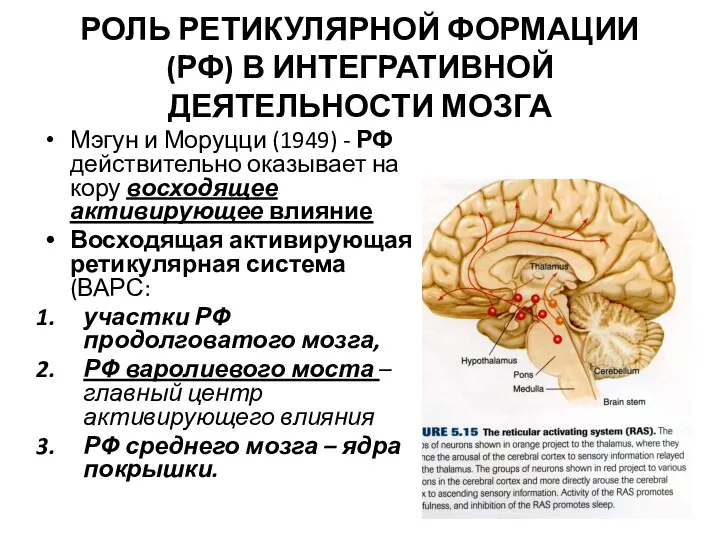
РОЛЬ РЕТИКУЛЯРНОЙ ФОРМАЦИИ (РФ) В ИНТЕГРАТИВНОЙ ДЕЯТЕЛЬНОСТИ МОЗГА
Мэгун и Моруцци (1949)
РОЛЬ РЕТИКУЛЯРНОЙ ФОРМАЦИИ (РФ) В ИНТЕГРАТИВНОЙ ДЕЯТЕЛЬНОСТИ МОЗГА
Мэгун и Моруцци (1949)
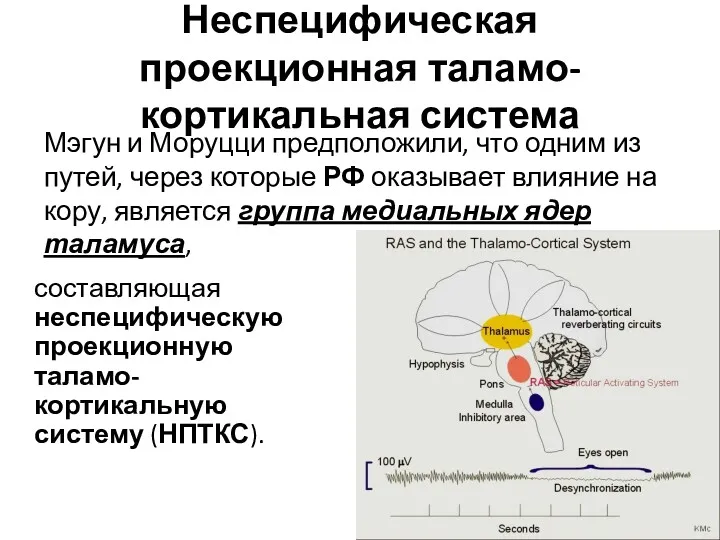
Неспецифическая проекционная таламо-кортикальная система
составляющая неспецифическую проекционную таламо-кортикальную систему (НПТКС).
Мэгун и
Неспецифическая проекционная таламо-кортикальная система
составляющая неспецифическую проекционную таламо-кортикальную систему (НПТКС).
Мэгун и

СОН
Благослови, господь, изобретшего сон.
М. Сервантес де Сааведра. "Дон Кихот"
Сон — это
СОН
Благослови, господь, изобретшего сон.
М. Сервантес де Сааведра. "Дон Кихот"
Сон — это

Сон - активное физиологическое состояние, которое характеризуется потерей активных психических связей
Сон - активное физиологическое состояние, которое характеризуется потерей активных психических связей

Выделяют несколько видов сна:
физиологический периодический ежесуточный сон
нефизиологический сон:
- наркотический сон, вызываемый
Выделяют несколько видов сна:
физиологический периодический ежесуточный сон
нефизиологический сон:
- наркотический сон, вызываемый

СТРУКТУРА СНА
"Медленный" сон:
1. Стадия тета-волн.
2. Стадия «сонных веретен» и «К-комплексов».
3. Стадия
СТРУКТУРА СНА
"Медленный" сон:
1. Стадия тета-волн.
2. Стадия «сонных веретен» и «К-комплексов».
3. Стадия

"Медленный" сон:
Очень поверхностный сон. Высокоамплитудные медленные тета-волны (3-7 Гц).
Поверхностный сон.
"Медленный" сон:
Очень поверхностный сон. Высокоамплитудные медленные тета-волны (3-7 Гц).
Поверхностный сон.
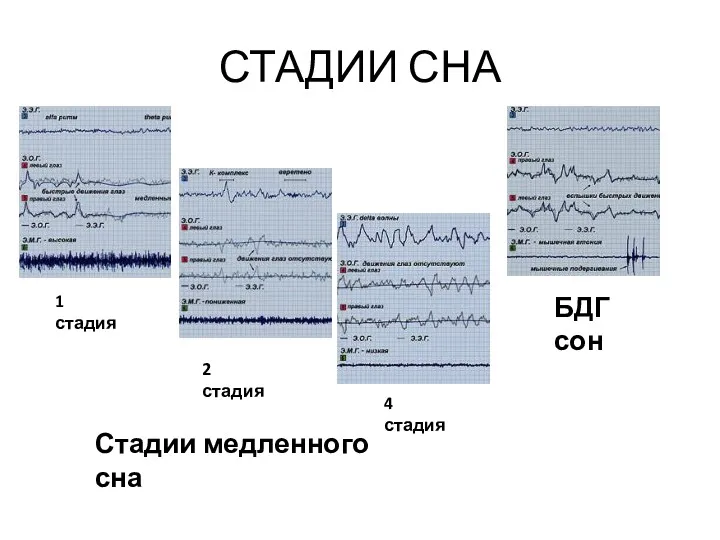
СТАДИИ СНА
1 стадия
2 стадия
БДГ сон
4 стадия
Стадии медленного сна
СТАДИИ СНА
1 стадия
2 стадия
БДГ сон
4 стадия
Стадии медленного сна

"Быстрый" сон (БДГ- сон)
Основной отличительной особенностью "быстрого" сна является наличие быстрых
"Быстрый" сон (БДГ- сон)
Основной отличительной особенностью "быстрого" сна является наличие быстрых

ТЕОРИИ СНА
Переход от бодрствования ко сну предполагает два возможных пути:
Пассивный, в
ТЕОРИИ СНА
Переход от бодрствования ко сну предполагает два возможных пути:
Пассивный, в

1. Гипотеза гипнотоксинов или гуморальная теория
Лежандр и Пьерон (1910)
Сон обусловлен накоплением
1. Гипотеза гипнотоксинов или гуморальная теория
Лежандр и Пьерон (1910)
Сон обусловлен накоплением

Традиционные обсуждаемые нейрохимические агенты, имеющие значение в организации цикла "сон- бодрствование":
Традиционные обсуждаемые нейрохимические агенты, имеющие значение в организации цикла "сон- бодрствование":

Однако
многие факты и, в частности, то, что можно преодолеть сон интересным
Однако
многие факты и, в частности, то, что можно преодолеть сон интересным

2. Теория центров сна
Фон Экономо (1930); Гесс (1933)
передний гипоталамус
2. Теория центров сна
Фон Экономо (1930); Гесс (1933)
передний гипоталамус

Однако
в дальнейшем было показано, что состояние сна можно было получить при
Однако
в дальнейшем было показано, что состояние сна можно было получить при

3. Теория условного торможения
И.П. Павлов даже сделал вывод о том, что
3. Теория условного торможения
И.П. Павлов даже сделал вывод о том, что

4. Теории деафферентации сенсорных систем
Бремер, конец 1930-х годов
Для бодрствования необходим
4. Теории деафферентации сенсорных систем
Бремер, конец 1930-х годов
Для бодрствования необходим

Однако
даже в изолированном переднем мозге появляются ритмичные колебания, характерные для цикла
Однако
даже в изолированном переднем мозге появляются ритмичные колебания, характерные для цикла

5. Теория неспецифических регуляторов сна
Активирующие структуры:
ВАРС
Неспецифическая и специфическая таламическая система
Ядра заднего
5. Теория неспецифических регуляторов сна
Активирующие структуры:
ВАРС
Неспецифическая и специфическая таламическая система
Ядра заднего

Однако
в ретикулярной формации существуют не только центры бодрствования, но и сна.
Однако
в ретикулярной формации существуют не только центры бодрствования, но и сна.

6.Серотонинергическая теория
М. Жуве - конец 60-х годов
В верхних отделах ствола
6.Серотонинергическая теория
М. Жуве - конец 60-х годов
В верхних отделах ствола
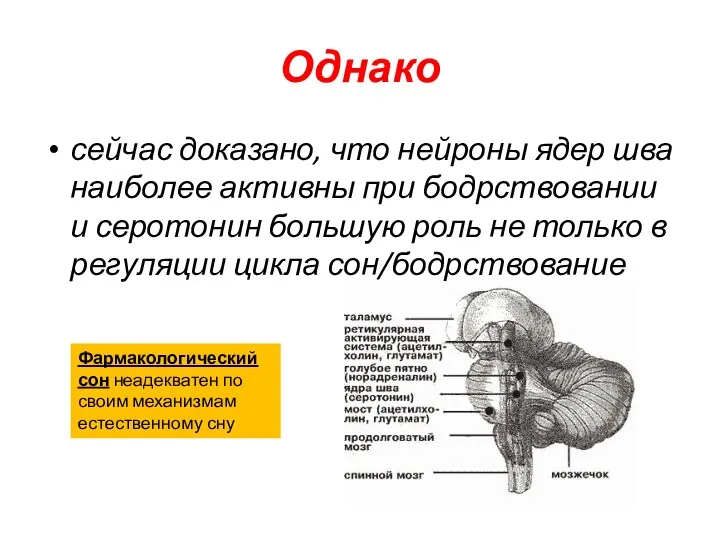
Однако
сейчас доказано, что нейроны ядер шва наиболее активны при бодрствовании и
Однако
сейчас доказано, что нейроны ядер шва наиболее активны при бодрствовании и

ПАМЯТЬ
ПАМЯТЬ
Биологическая память – это фундаментальное свойство живой матери приобретать, сохранять и
Биологическая память – это фундаментальное свойство живой матери приобретать, сохранять и

Генетическая - эволюционно наиболее ранняя форма памяти. Материальным носителем является геном
Генетическая - эволюционно наиболее ранняя форма памяти. Материальным носителем является геном

НЕРВНАЯ ПАМЯТЬ
Нервная память - совокупность процессов фиксации, хранения и извлечения информации,
НЕРВНАЯ ПАМЯТЬ
Нервная память - совокупность процессов фиксации, хранения и извлечения информации,

ВРЕМЕННАЯ ОРГАНИЗАЦИЯ ПАМЯТИ
Концепция временной организации памяти принадлежит канадскому психологу Д. Хеббу
ВРЕМЕННАЯ ОРГАНИЗАЦИЯ ПАМЯТИ
Концепция временной организации памяти принадлежит канадскому психологу Д. Хеббу

Классификация видов долговременной памяти, предложенная И.С.Бериташвили (1968)
образная (декларативная) - сохранение и
Классификация видов долговременной памяти, предложенная И.С.Бериташвили (1968)
образная (декларативная) - сохранение и

Фиксация образов, явлений происходит в три этапа:
формирование энграммы,
сортировка и
Фиксация образов, явлений происходит в три этапа:
формирование энграммы,
сортировка и

МЕХАНИЗМЫ ПАМЯТИ
Кратковременная, и долговременная памяти имеют разную природу и регулируются разными
МЕХАНИЗМЫ ПАМЯТИ
Кратковременная, и долговременная памяти имеют разную природу и регулируются разными

МЕХАНИЗМЫ КРАТКОВРЕМЕННОЙ ПАМЯТИ
По современным представлениям в основе кратковременной памяти лежат два
МЕХАНИЗМЫ КРАТКОВРЕМЕННОЙ ПАМЯТИ
По современным представлениям в основе кратковременной памяти лежат два

Механизм кратковременной синаптической пластичности
- изменение амплитуды постсинаптического потенциала за счет
Механизм кратковременной синаптической пластичности
- изменение амплитуды постсинаптического потенциала за счет

МЕХАНИЗМЫ ДОЛГОВРЕМЕННОЙ ПАМЯТИ
Механизм долговременной синаптической пластичности – изменение амплитуды постсинаптического потенциала
МЕХАНИЗМЫ ДОЛГОВРЕМЕННОЙ ПАМЯТИ
Механизм долговременной синаптической пластичности – изменение амплитуды постсинаптического потенциала

Основной причиной является изменение концентрации внутриклеточного кальция в постсинаптической клетке:
Через NMDAзависимые
Основной причиной является изменение концентрации внутриклеточного кальция в постсинаптической клетке:
Через NMDAзависимые

Модель формирования долговременной памяти (Анохин К.В., 1997):
Активация кальцием внутриклеточных протеинкиназ (кальций-кальмодулинзависимые
Модель формирования долговременной памяти (Анохин К.В., 1997):
Активация кальцием внутриклеточных протеинкиназ (кальций-кальмодулинзависимые

ЛОКАЛИЗАЦИЯ СЛЕДОВ ПАМЯТИ
Большинство нейрофизиологов полагает, что запечатление и извлечение энграмм идет
ЛОКАЛИЗАЦИЯ СЛЕДОВ ПАМЯТИ
Большинство нейрофизиологов полагает, что запечатление и извлечение энграмм идет

МОТИВАЦИИ
МОТИВАЦИИ

Мотивация - стремление к удовлетворению потребностей (влечения, побуждения, драйвы), имеющая эмоциональную
Мотивация - стремление к удовлетворению потребностей (влечения, побуждения, драйвы), имеющая эмоциональную

ПЕРИФЕРИЧЕСКИЕ И ЦЕНТРАЛЬНЫЕ МЕХАНИЗМЫ МОТИВАЦИЙ
Причиной мотиваций являются преимущественно раздражители внутренней среды
ПЕРИФЕРИЧЕСКИЕ И ЦЕНТРАЛЬНЫЕ МЕХАНИЗМЫ МОТИВАЦИЙ
Причиной мотиваций являются преимущественно раздражители внутренней среды

Мотивационные центры:
1. Центр голода - вентромедиальное ядро и латеральный ГПТ –
Мотивационные центры:
1. Центр голода - вентромедиальное ядро и латеральный ГПТ –

Мотивации – это особое интегрированное состояние головного мозга, в которое каждая
Мотивации – это особое интегрированное состояние головного мозга, в которое каждая

ЭМОЦИИ
ЭМОЦИИ

Эмоция (волнение, возбуждение) – субъективное состояние человека и животных, которое возникает
Эмоция (волнение, возбуждение) – субъективное состояние человека и животных, которое возникает

Функции эмоций:
Эмоции являются одним из главных механизмов регуляции психической деятельности, направленной
Функции эмоций:
Эмоции являются одним из главных механизмов регуляции психической деятельности, направленной

ТЕОРИИ ВОЗНИКНОВЕНИЯ ЭМОЦИЙ
1. Периферическая теория (Джеймс – Ланге, 1885). Эмоции рассматриваются
ТЕОРИИ ВОЗНИКНОВЕНИЯ ЭМОЦИЙ
1. Периферическая теория (Джеймс – Ланге, 1885). Эмоции рассматриваются

3. Центральная лимбическая теория Пейпс (Papez), 1937. Ведущая роль в формировании
3. Центральная лимбическая теория Пейпс (Papez), 1937. Ведущая роль в формировании

4. Активационная теория (Линдсли, 1958). Подчеркивает роль РФ в формировании эмоций
4. Активационная теория (Линдсли, 1958). Подчеркивает роль РФ в формировании эмоций

Симонов различает четыре состояния эмоционального напряжения (СН):
Симонов различает четыре состояния эмоционального напряжения (СН):

Периферические компоненты эмоций представлены:
произвольно управляемыми проявлениями, такими, как движения, мимика, речь,
Периферические компоненты эмоций представлены:
произвольно управляемыми проявлениями, такими, как движения, мимика, речь,
 Внесение удобрений под овощные растения
Внесение удобрений под овощные растения Конечный мозг
Конечный мозг Дикие и домашние животные
Дикие и домашние животные Электронный задачник. Часть 1. Содержание (Моногибридное скрещивание)
Электронный задачник. Часть 1. Содержание (Моногибридное скрещивание) Гипотезы происхождения жизни на Земле
Гипотезы происхождения жизни на Земле Разнообразие животных. Урок 19
Разнообразие животных. Урок 19 Становление наук о человеке. (8 класс)
Становление наук о человеке. (8 класс) Стати собак
Стати собак Үсемлек белән тукланучы һәм ерткыч хайваннар
Үсемлек белән тукланучы һәм ерткыч хайваннар Обмен углеводов-1
Обмен углеводов-1 Познавательные беседы о птицах
Познавательные беседы о птицах Влияние куриного помёта на урожайность огурцов
Влияние куриного помёта на урожайность огурцов Фізіологія нейрона. Збудження та гальмування в ЦНС. Інтегративна функція нейронних ланцюгів
Фізіологія нейрона. Збудження та гальмування в ЦНС. Інтегративна функція нейронних ланцюгів Белки - строение, функции
Белки - строение, функции Красная книга Алтайского края
Красная книга Алтайского края Томас Морган Хант
Томас Морган Хант Размножение живых организмов. Гаметогенез
Размножение живых организмов. Гаметогенез Моллюски. Класс брюхоногие. Виноградная улитка
Моллюски. Класс брюхоногие. Виноградная улитка Вода - источник жизни комнатных растений
Вода - источник жизни комнатных растений Биопотенциалы покоя. Генерация и распространение потенциала действия. (Лекция 5)
Биопотенциалы покоя. Генерация и распространение потенциала действия. (Лекция 5) Развитие генетики
Развитие генетики Тип Членистоногие
Тип Членистоногие Тварини
Тварини Плечевое сплетение
Плечевое сплетение Мутации. Классификации. Мутации и мобильные элементы. Мобильные элементы генома, классификация и их генетическая роль
Мутации. Классификации. Мутации и мобильные элементы. Мобильные элементы генома, классификация и их генетическая роль Рибонуклеиновые кислоты АТФ. Химический состав клетки. Рибонуклеиновые кислоты
Рибонуклеиновые кислоты АТФ. Химический состав клетки. Рибонуклеиновые кислоты Головний мозок
Головний мозок Цветы и растения, как символы стран мира
Цветы и растения, как символы стран мира