- Сенсорные системы. Общая характеристика сенсорных рецепторов. Зрительная система

Содержание
- 2. Общая характеристика сенсорных рецепторов
- 3. Стимул Рецепторный потенциал Сенсорный сигнал Трансдукция Первичночувствующие рецепторы Вторичночувствующие рецепторы Механизмы преобразования сенсорного стимула
- 4. Разнообразие рецепторных клеток сенсорных органов Красным обведены первичночувствующие клетки Синим обведены вторичночувствующие клетки
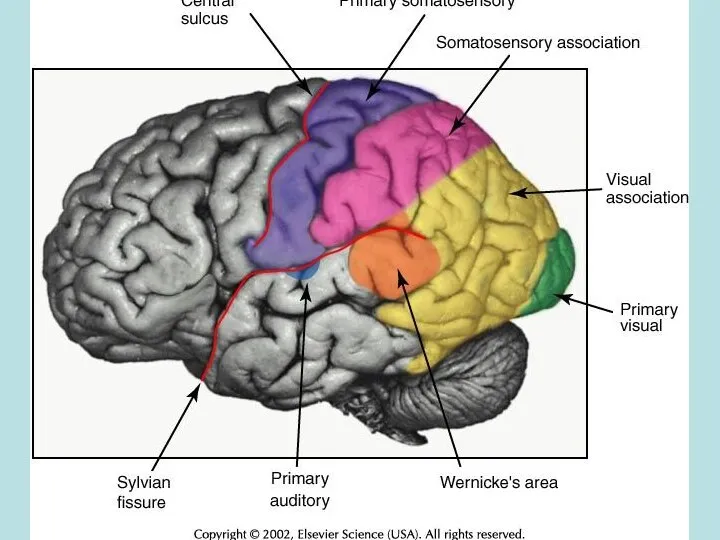
- 5. Receptor morphology and relationship to ganglion cells in the somatosensory, auditory, and visual systems. Receptors are
- 6. Классификация рецепторов,основанная на их модальности
- 7. Последовательность восприятия сенсорного стимула
- 8. Этапы генерации сенсорного сигнала
- 9. Модальность Локализация Интенсивность рецепторный потенциал градуален, однако интенсивность не прямо пропорциональна амплитуде Закон Вебера-Фехнера, «основной психофизический
- 10. Зрение
- 11. Более половины всех сенсорных клеток человека реализуют зрение Значительная часть коры больших полушарий заняты обработкой зрительных
- 12. Глаз и его компоненты Рефракция + 48 D Аккомодация 14 D
- 13. Figure 16.8 Структуры глаза, ответственные за адаптацию и аккомодацию
- 14. Эволюция зрительного аппарата
- 15. Фасеточный глаз насекомого
- 16. Глаз и сетчатка «Сетчатка – это часть мозга, помещенная в глаз» / Рамон-a-Кахал, 1901 г/ Fovea
- 17. Строение сетчатки и фоторецепторов Сетчатка осуществляет фототрансдукцию – процесс, посредством которого поглощение кванта света фоторецепторными клетками
- 18. Поверхность сетчатки (микрофотография) Светлые клетки - палочки Темные клетки - колбочки В периферических отделах сетчатки преобладают
- 19. Клеточная организация сетчатки позвоночных Ядра палочек и колбочек Ядра биполярных, горизонтальных и амакриновых клеток Ядра ганглиозных
- 20. Summary diagram of the cell types and connections in the primate retina. R, rod; C, cone;
- 21. Палочки и колбочки: световая микроскопия, флуоресцентная краска Модель палочки и колбочки
- 22. Наружный сегмент палочки Одиночная колбочка Двойная колбочка Колбочка Фоторецепторы сетчатки: электронная микроскопия
- 23. Палочки и колбочки У человека в сетчатке имеется примерно 120 млн палочек и от 4 до
- 24. Фоторецепторные мембранные диски в наружных сегментах палочек и колбочек Родопсин
- 25. 11-цис ретиналь – хромофорная группа родопсина (в темноте) Белковая часть (опсин) 11-цис ретиналь Родопсин От палочки
- 26. Структура ретинали, ее цис- и транс- формы Достаточен 1 фотон
- 27. Превращения родопсина при воздействии фотона света: первый этап восприятия света
- 28. 11-цис ретиналь показан фиолетовым цветом Хромофорный центр родопсина Функции ретиналя как хромофорной группы: Спектральная настройка зрительных
- 29. Второй этап: взаимодействие метародопсина II с ГТФ-связывющим белком (трансдуцином), что вызывает его диссоциацию на Gα и
- 30. Три ключевых стадии : фотоизомеризация 11-цис ретиналя взаимодействие метародопсина II с G-белком разрыв связи полностью-транс ретиналя
- 31. Механизмы трансдукции (состояние темноты) Фотородопсин и трансдуцин разобщены Трансдуцин не диссоциирован ФДЭ ингибирована Концентрация цГМФ высока
- 32. Темнота Свет - 30 мВ - 70 мВ Деполяризована Гиперполяризована Ионный механизм генерации сигнала в фоторецепторах
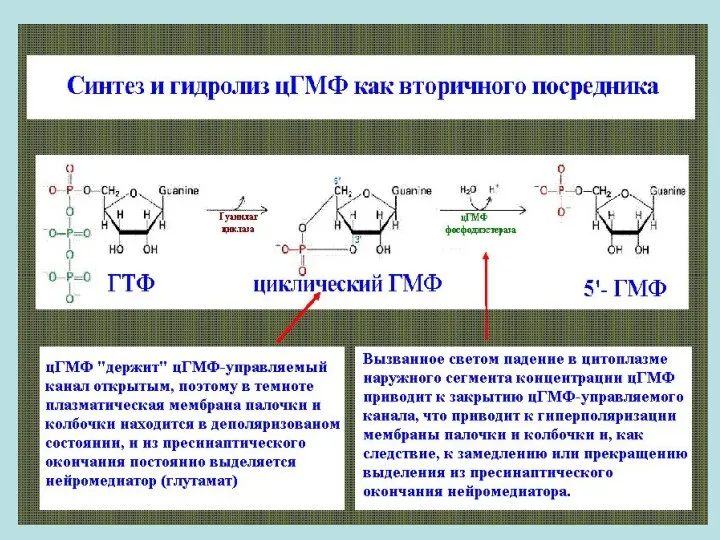
- 33. В темноте Внутриклеточным посредником возбуждения в палочках и колбочках служит циклический гуанозинмонофосфат (3’, 5’-цГМФ). Концентрация цГМФ
- 34. Механизмы трансдукции На свету: Трансдуцин диссоциирован (2-й этап) альфа-субед. активирует ФДЭ (3-й этап) Уровень цГМФ снижается
- 35. John Hopp родопсин трансдуцин фосфодиэстераза
- 36. На свету Перевод молекулы родопсина квантом света в активное состояние (метародопсин II, обозначенный на рис. как
- 37. Возврат фоторецептора в темновое состояние (выделено синим) Через открытые каналы входит Са Активируется белок – рековерин,
- 38. Выключение каскада и возврат его в темновое состояние происходят на всех стадиях. (1) Метародопсин II теряет
- 40. П е р е р ы в
- 41. Схема клеточной организации сетчатки Пространственная конвергенция сигналов от нескольких рецепторов на одном биполяре. Хотя общее число
- 42. Строение сетчатки Путь зрительного сигнала от фоторецепторов на биполяры, затем на ганглиозные клетки Их аксоны в
- 43. Палочки не связаны с ганглиозными клетками непосредственно, существуют два пути передачи сигнала: (i) синапс биполяра палочки
- 44. Существует относительная корреляция между субъективной светлотой и частотой разряда on-ганглиозных клеток и соответственно темнотой и частотой
- 45. Нейрональная часть сетчатки (без фоторецепторов) наружный синаптический слой горизонтальные, биполярные и амакриновые клетки внутренний синаптический слой
- 49. Рецептивное поле – зона сетчатки, в которой изменяется активность нейрона при падении на нее пятна света.
- 50. Сохраняющие (Н) и инвертирующие (D) знак сигнала синапсы колбочек с биполярными клетками. Горизонтальная клетка (H) (D)
- 51. WHAT THE FROG'S EYE TELLS THE FROG'S BRAIN * t Y. LETIVIN, H. R. MATURANA,§ W.
- 52. Н- биполяр отвечает гиперполяризацией на освещение центра рецептивного поля Н- биполяр отвечает деполяризацией на освещение вокруг
- 54. Реакция сетчатки на пятна и кольца света
- 55. Электрические ответы клеток сетчатки на световые пятна и кольца
- 56. Ганглиозные клетки не передают информацию об абсолютном уровне освещенности. Их задача измерять различия света в пределах
- 57. Электрические ответы клеток сетчатки на световые пятна и кольца
- 58. The retino-geniculo-cortical pathway in the human. Optic nerve axons from the nasal retina cross at the
- 59. Трейсерный метод выявления нервных связей. Пероксидаза, введенная в сетчатку, транспортируется по аксонам и выявляется в нейронах
- 60. The six-layered LGN of the macaque monkey. The arrow from layer 6 to 1 indicates organization
- 62. Examples of sensory magnification in the visual and somatosensory systems. (A) Determination of a visual field
- 64. Глаз и его компоненты
- 66. Скачать презентацию

Общая характеристика сенсорных рецепторов
Общая характеристика сенсорных рецепторов
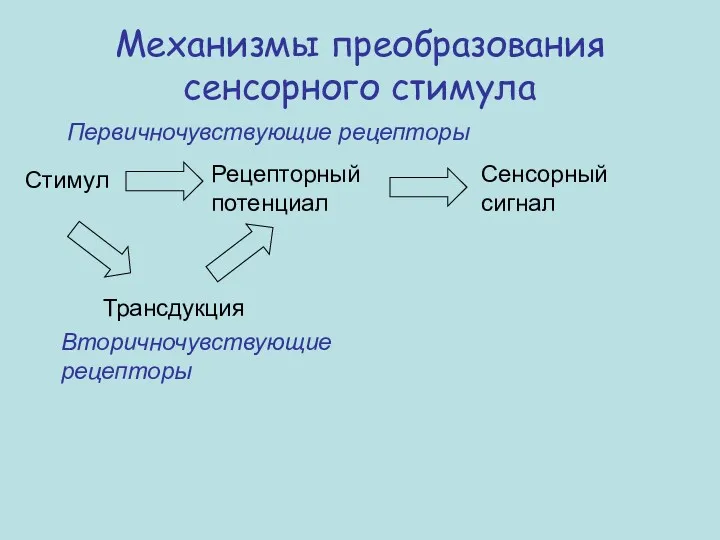
Стимул
Рецепторный потенциал
Сенсорный сигнал
Трансдукция
Первичночувствующие рецепторы
Вторичночувствующие рецепторы
Механизмы преобразования сенсорного стимула
Стимул
Рецепторный потенциал
Сенсорный сигнал
Трансдукция
Первичночувствующие рецепторы
Вторичночувствующие рецепторы
Механизмы преобразования сенсорного стимула
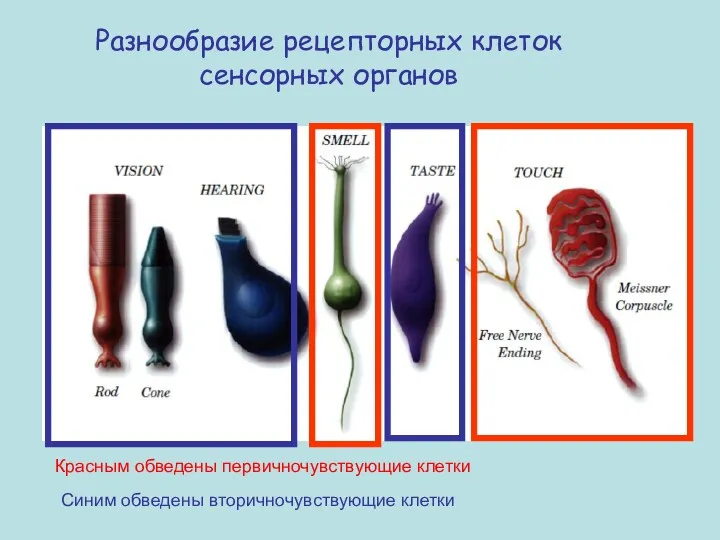
Разнообразие рецепторных клеток сенсорных органов
Красным обведены первичночувствующие клетки
Синим обведены вторичночувствующие клетки
Разнообразие рецепторных клеток сенсорных органов
Красным обведены первичночувствующие клетки
Синим обведены вторичночувствующие клетки
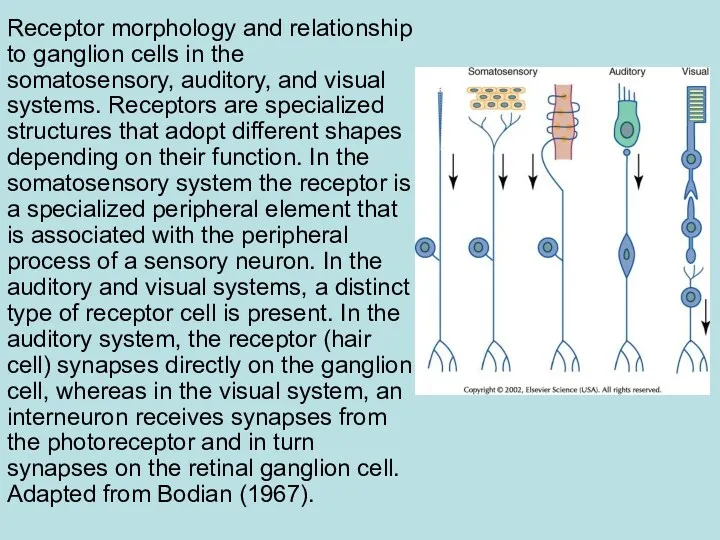
Receptor morphology and relationship to ganglion cells in the somatosensory, auditory,
Receptor morphology and relationship to ganglion cells in the somatosensory, auditory,
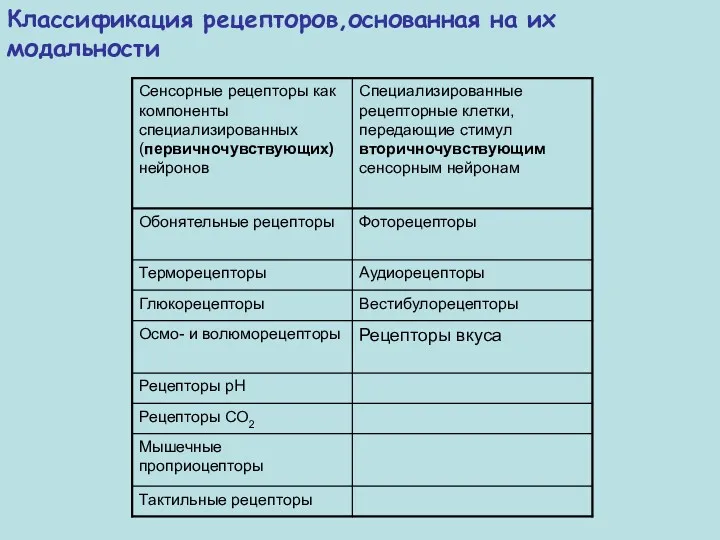
Классификация рецепторов,основанная на их модальности
Классификация рецепторов,основанная на их модальности
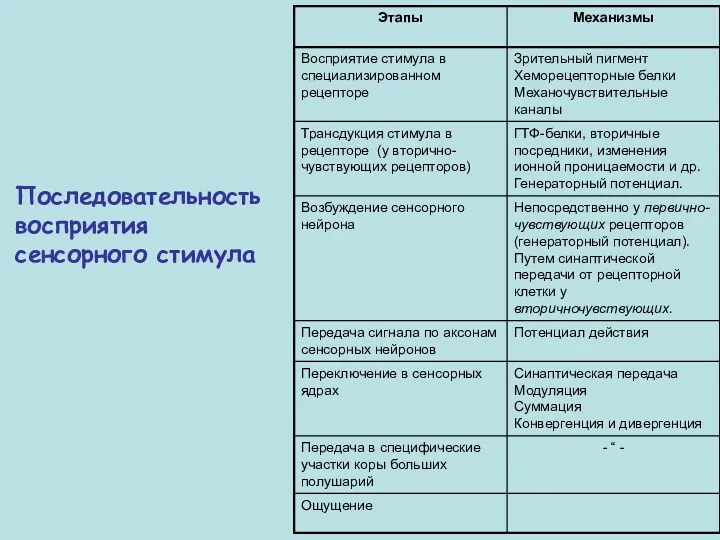
Последовательность восприятия сенсорного стимула
Последовательность восприятия сенсорного стимула
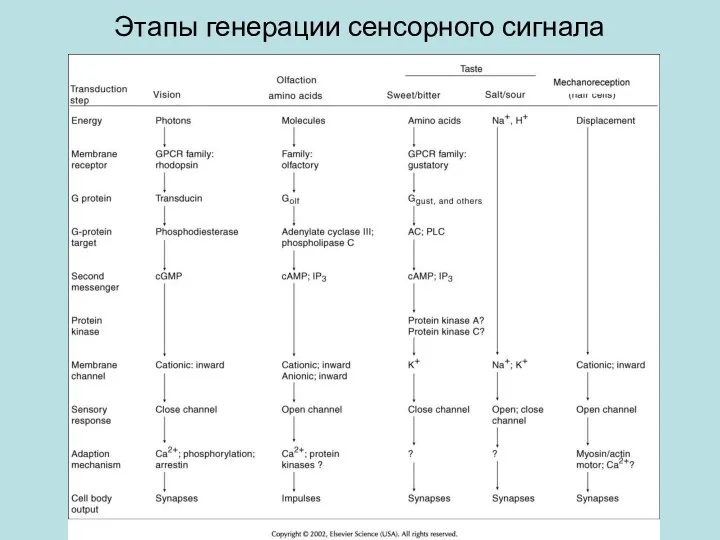
Этапы генерации сенсорного сигнала
Этапы генерации сенсорного сигнала

Модальность
Локализация
Интенсивность
рецепторный потенциал градуален, однако интенсивность не прямо
Модальность
Локализация
Интенсивность
рецепторный потенциал градуален, однако интенсивность не прямо

Зрение
Зрение
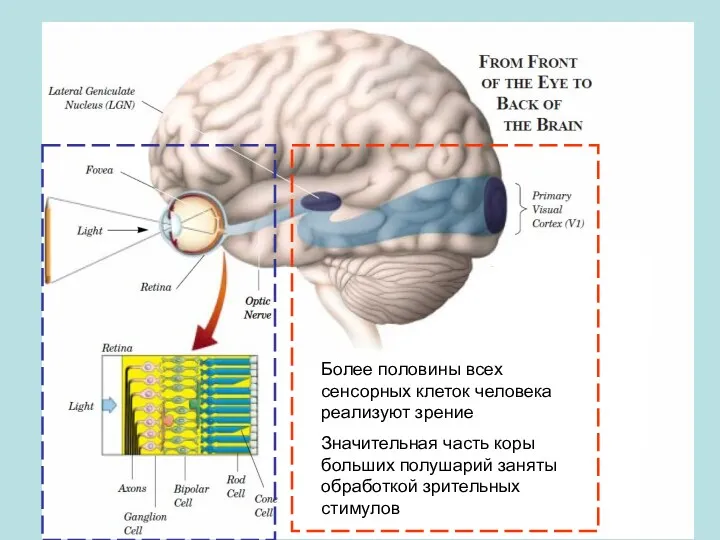
Более половины всех сенсорных клеток человека реализуют зрение
Значительная часть коры больших
Более половины всех сенсорных клеток человека реализуют зрение
Значительная часть коры больших
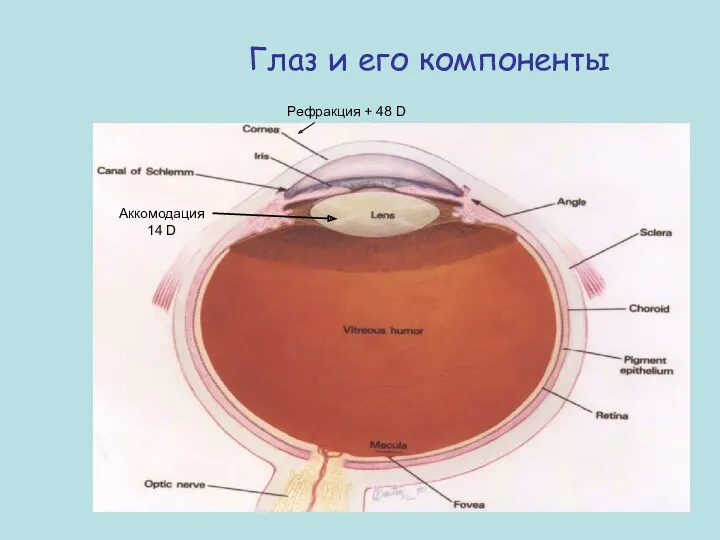
Глаз и его компоненты
Рефракция + 48 D
Аккомодация 14 D
Глаз и его компоненты
Рефракция + 48 D
Аккомодация 14 D
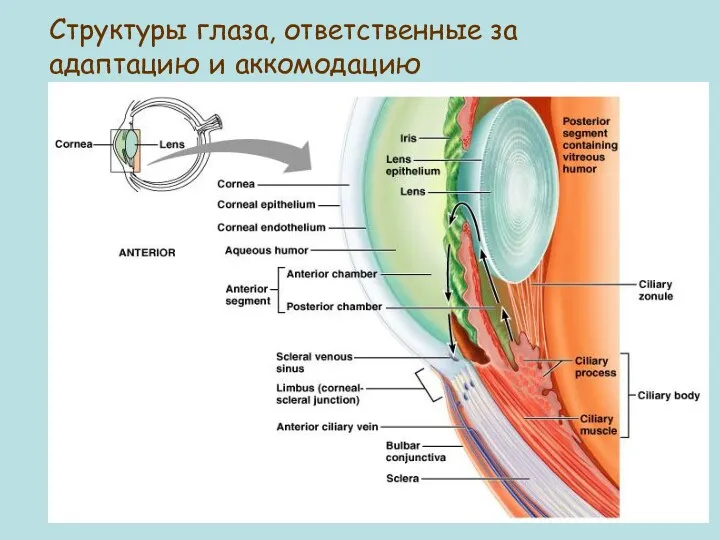
Figure 16.8
Структуры глаза, ответственные за адаптацию и аккомодацию
Figure 16.8
Структуры глаза, ответственные за адаптацию и аккомодацию
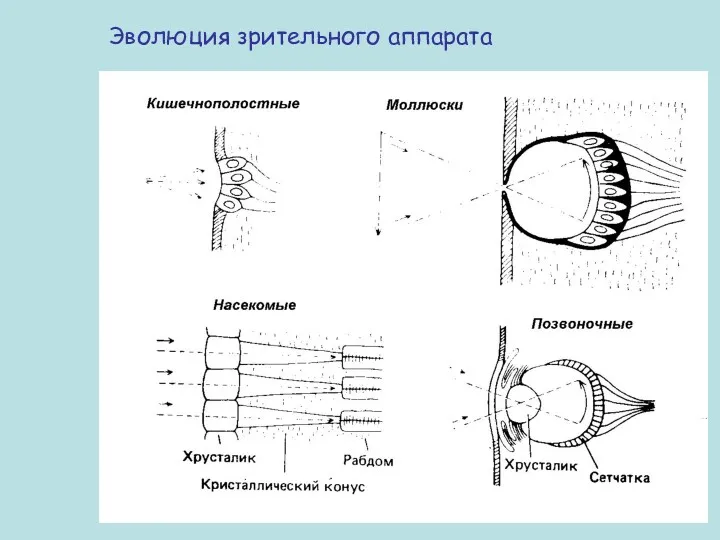
Эволюция зрительного аппарата
Эволюция зрительного аппарата
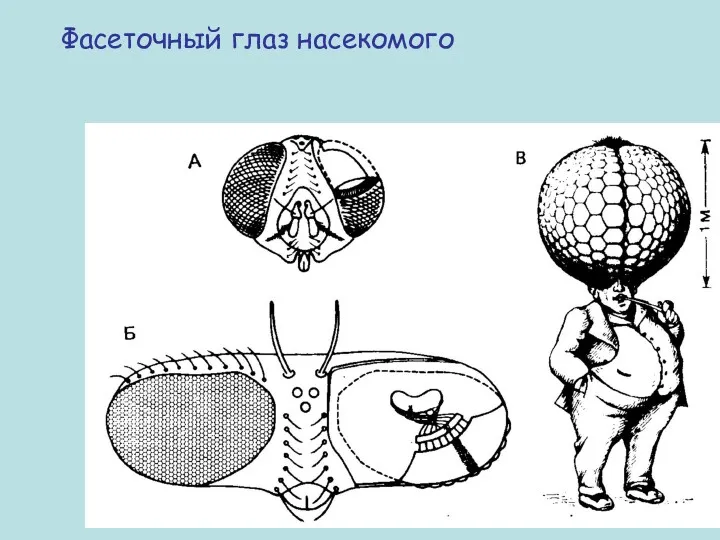
Фасеточный глаз насекомого
Фасеточный глаз насекомого
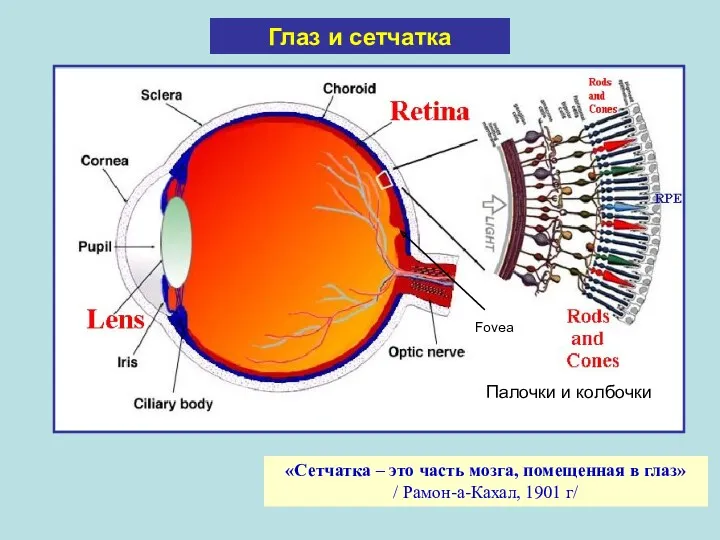
Глаз и сетчатка
«Сетчатка – это часть мозга, помещенная в глаз» /
Глаз и сетчатка
«Сетчатка – это часть мозга, помещенная в глаз» /

Строение сетчатки и фоторецепторов
Сетчатка осуществляет фототрансдукцию – процесс, посредством которого поглощение
Строение сетчатки и фоторецепторов
Сетчатка осуществляет фототрансдукцию – процесс, посредством которого поглощение
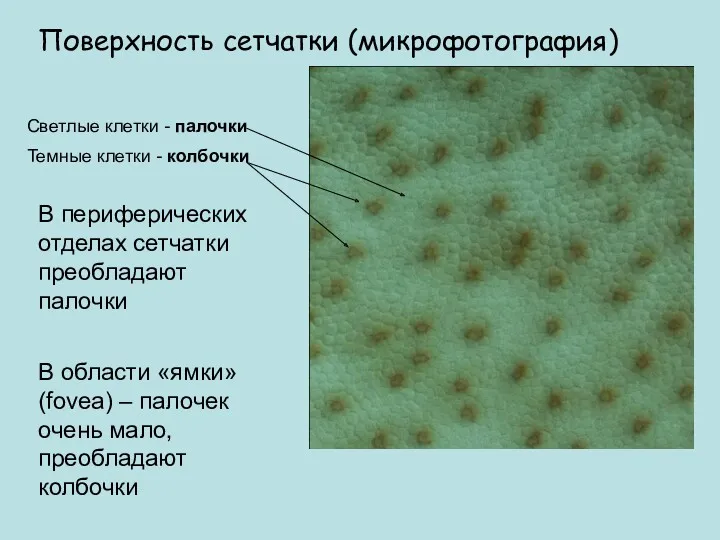
Поверхность сетчатки (микрофотография)
Светлые клетки - палочки
Темные клетки - колбочки
В периферических отделах
Поверхность сетчатки (микрофотография)
Светлые клетки - палочки
Темные клетки - колбочки
В периферических отделах
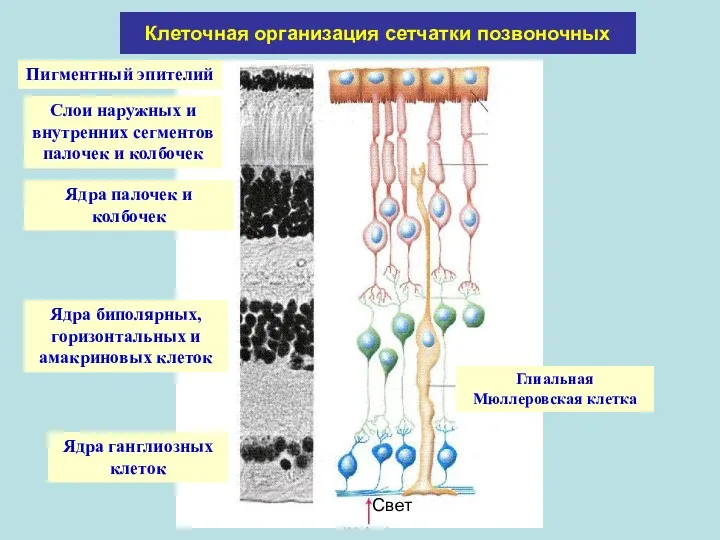
Клеточная организация сетчатки позвоночных
Ядра палочек и колбочек
Ядра биполярных, горизонтальных и амакриновых
Клеточная организация сетчатки позвоночных
Ядра палочек и колбочек
Ядра биполярных, горизонтальных и амакриновых
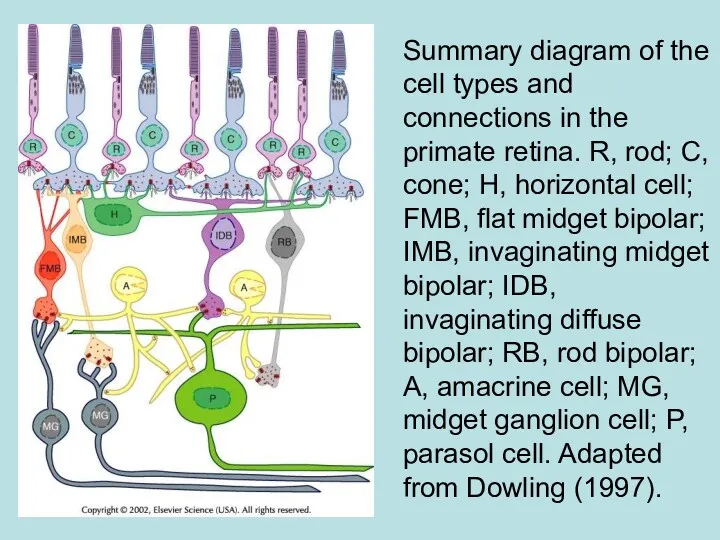
Summary diagram of the cell types and connections in the primate
Summary diagram of the cell types and connections in the primate
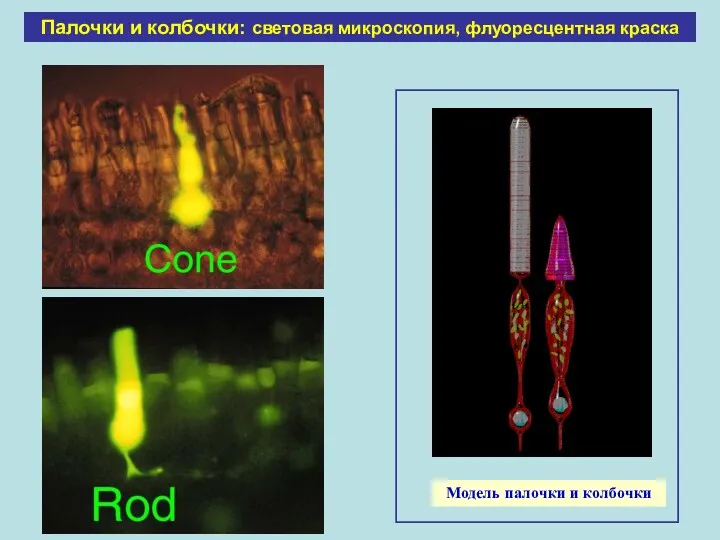
Палочки и колбочки: световая микроскопия, флуоресцентная краска
Модель палочки и колбочки
Палочки и колбочки: световая микроскопия, флуоресцентная краска
Модель палочки и колбочки
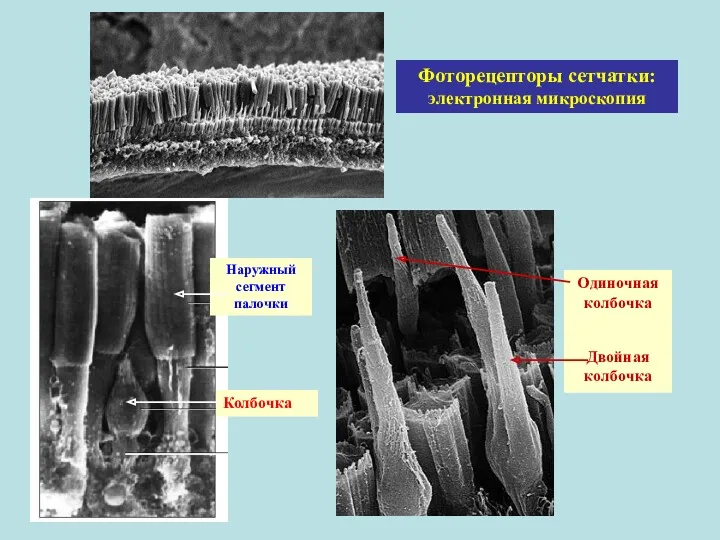
Наружный сегмент палочки
Одиночная колбочка
Двойная колбочка
Колбочка
Фоторецепторы сетчатки: электронная микроскопия
Наружный сегмент палочки
Одиночная колбочка
Двойная колбочка
Колбочка
Фоторецепторы сетчатки: электронная микроскопия
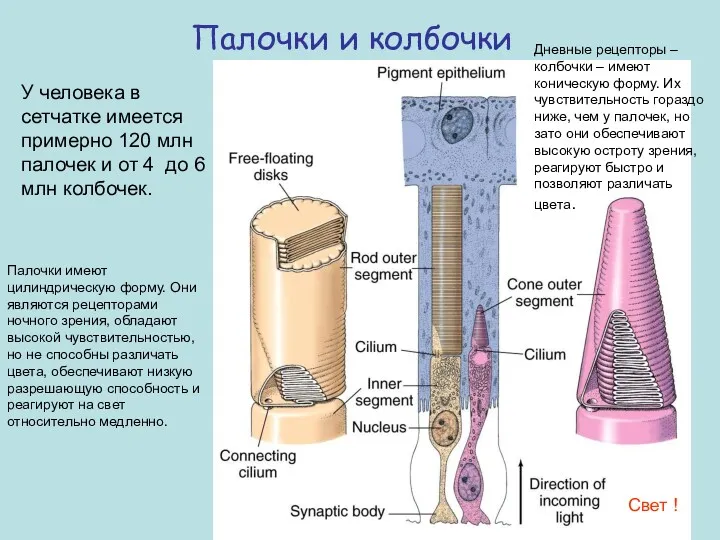
Палочки и колбочки
У человека в сетчатке имеется примерно 120 млн палочек
Палочки и колбочки
У человека в сетчатке имеется примерно 120 млн палочек
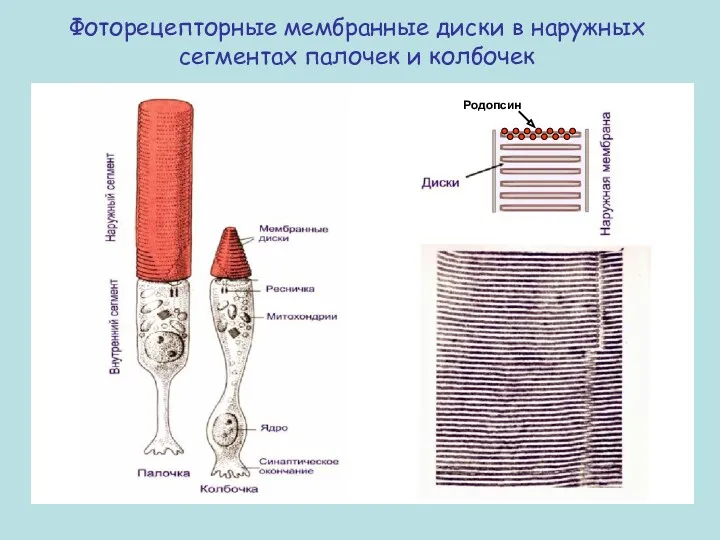
Фоторецепторные мембранные диски в наружных сегментах палочек и колбочек
Родопсин
Фоторецепторные мембранные диски в наружных сегментах палочек и колбочек
Родопсин
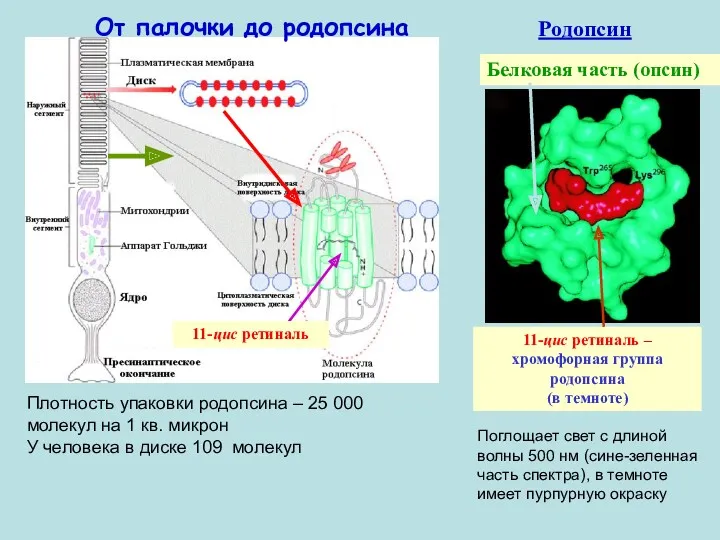
11-цис ретиналь – хромофорная группа родопсина
(в темноте)
Белковая часть (опсин)
11-цис ретиналь
Родопсин
От палочки
11-цис ретиналь – хромофорная группа родопсина
(в темноте)
Белковая часть (опсин)
11-цис ретиналь
Родопсин
От палочки
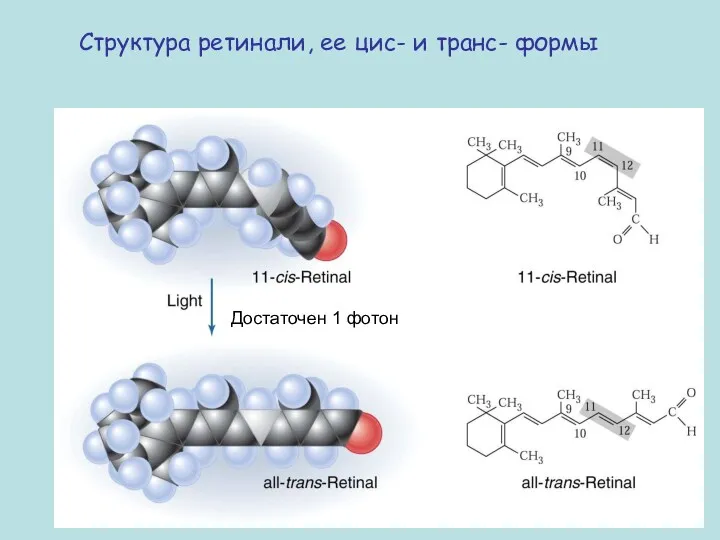
Структура ретинали, ее цис- и транс- формы
Достаточен 1 фотон
Структура ретинали, ее цис- и транс- формы
Достаточен 1 фотон
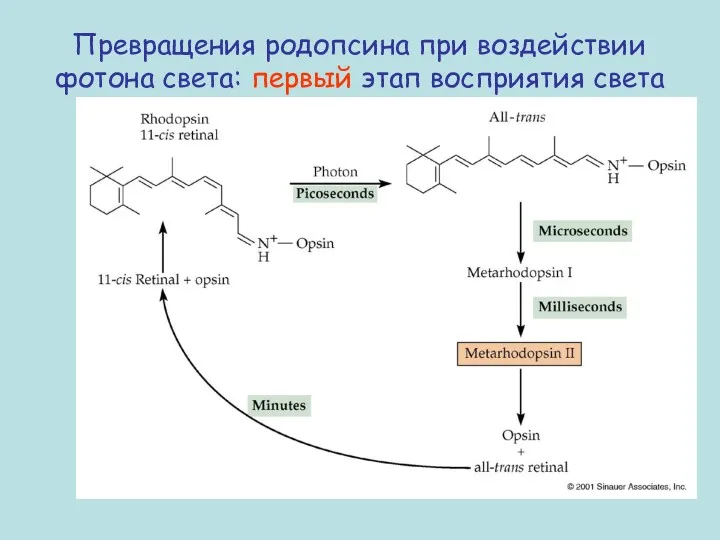
Превращения родопсина при воздействии фотона света: первый этап восприятия света
Превращения родопсина при воздействии фотона света: первый этап восприятия света
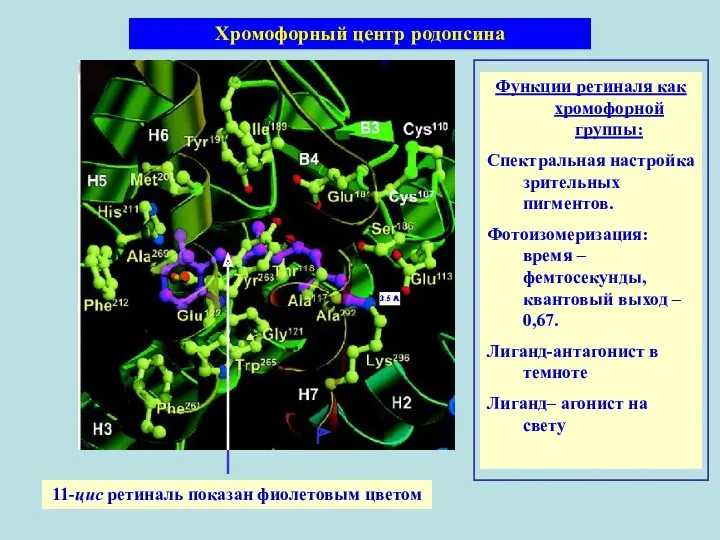
11-цис ретиналь показан фиолетовым цветом
Хромофорный центр родопсина
Функции ретиналя как хромофорной группы:
Спектральная
11-цис ретиналь показан фиолетовым цветом
Хромофорный центр родопсина
Функции ретиналя как хромофорной группы:
Спектральная
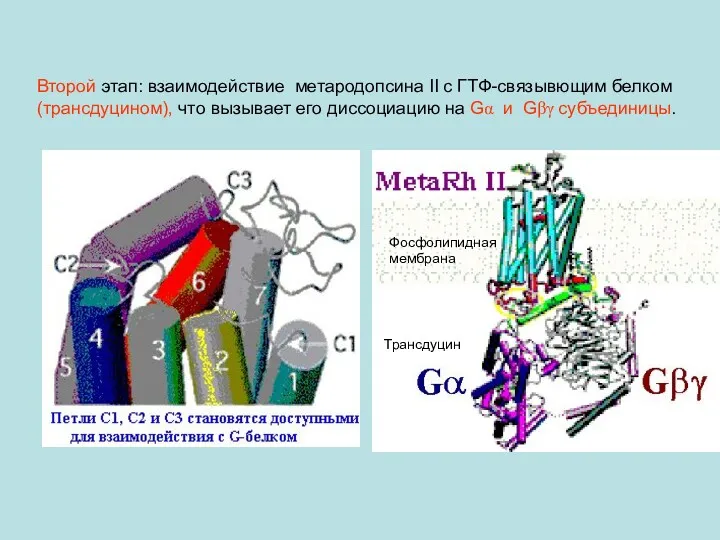
Второй этап: взаимодействие метародопсина II с ГТФ-связывющим белком (трансдуцином), что вызывает
Второй этап: взаимодействие метародопсина II с ГТФ-связывющим белком (трансдуцином), что вызывает
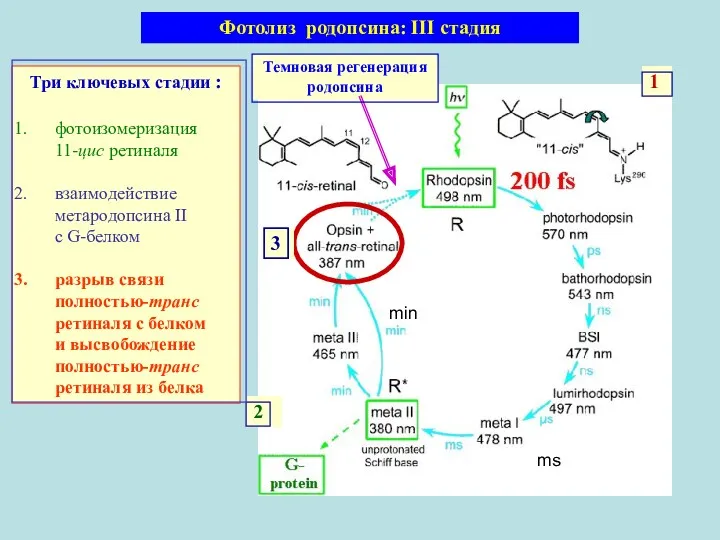
Три ключевых стадии :
фотоизомеризация 11-цис ретиналя
взаимодействие метародопсина II с G-белком
разрыв
Три ключевых стадии :
фотоизомеризация 11-цис ретиналя
взаимодействие метародопсина II с G-белком
разрыв
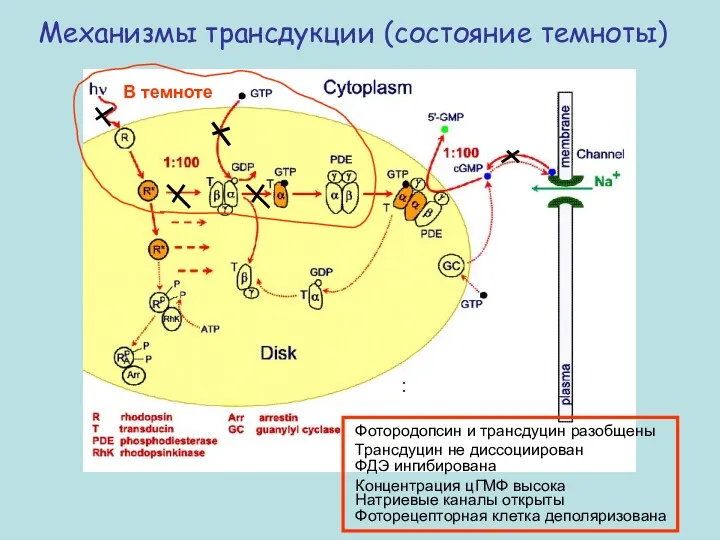
Механизмы трансдукции (состояние темноты)
Фотородопсин и трансдуцин разобщены Трансдуцин не диссоциирован
ФДЭ ингибирована
Концентрация
Механизмы трансдукции (состояние темноты)
Фотородопсин и трансдуцин разобщены Трансдуцин не диссоциирован
ФДЭ ингибирована
Концентрация
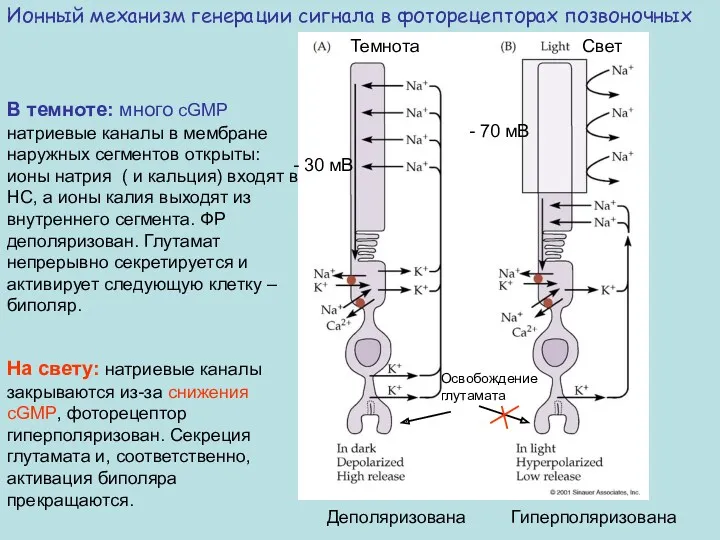
Темнота
Свет
- 30 мВ
- 70 мВ
Деполяризована Гиперполяризована
Ионный механизм генерации сигнала в
Темнота
Свет
- 30 мВ
- 70 мВ
Деполяризована Гиперполяризована
Ионный механизм генерации сигнала в

В темноте
Внутриклеточным посредником возбуждения в палочках и колбочках служит циклический гуанозинмонофосфат
В темноте
Внутриклеточным посредником возбуждения в палочках и колбочках служит циклический гуанозинмонофосфат
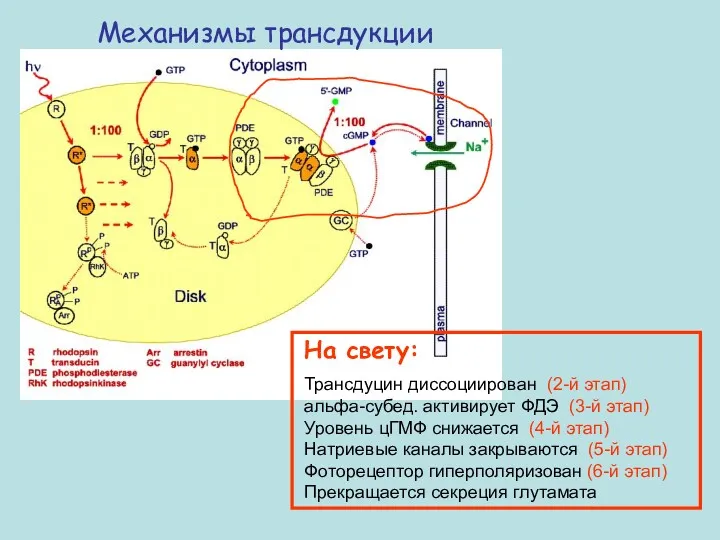
Механизмы трансдукции
На свету:
Трансдуцин диссоциирован (2-й этап) альфа-субед. активирует ФДЭ (3-й этап)
Механизмы трансдукции
На свету:
Трансдуцин диссоциирован (2-й этап) альфа-субед. активирует ФДЭ (3-й этап)
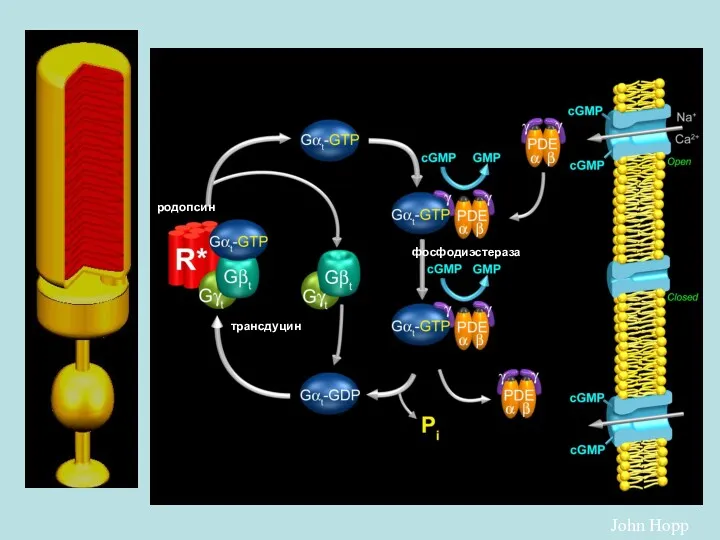
John Hopp
родопсин
трансдуцин
фосфодиэстераза
John Hopp
родопсин
трансдуцин
фосфодиэстераза

На свету
Перевод молекулы родопсина квантом света в активное состояние (метародопсин II,
На свету
Перевод молекулы родопсина квантом света в активное состояние (метародопсин II,
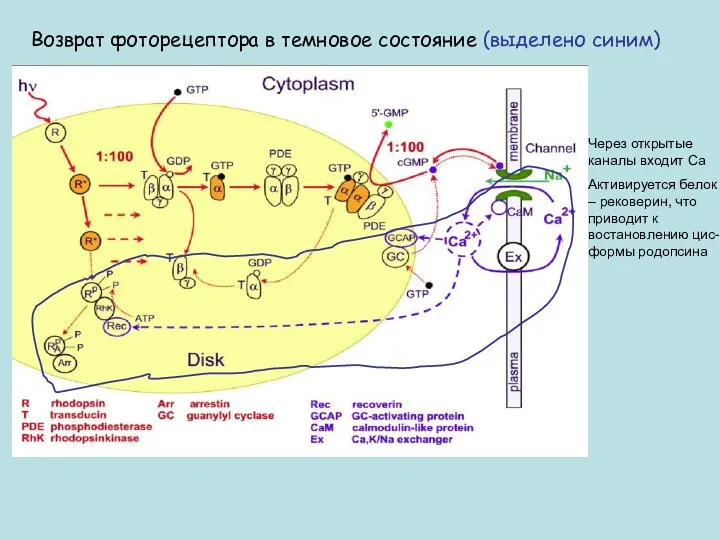
Возврат фоторецептора в темновое состояние (выделено синим)
Через открытые каналы входит Са
Активируется
Возврат фоторецептора в темновое состояние (выделено синим)
Через открытые каналы входит Са
Активируется

Выключение каскада и возврат его в темновое состояние происходят на всех
Выключение каскада и возврат его в темновое состояние происходят на всех

П е р е р ы в
П е р е р ы в
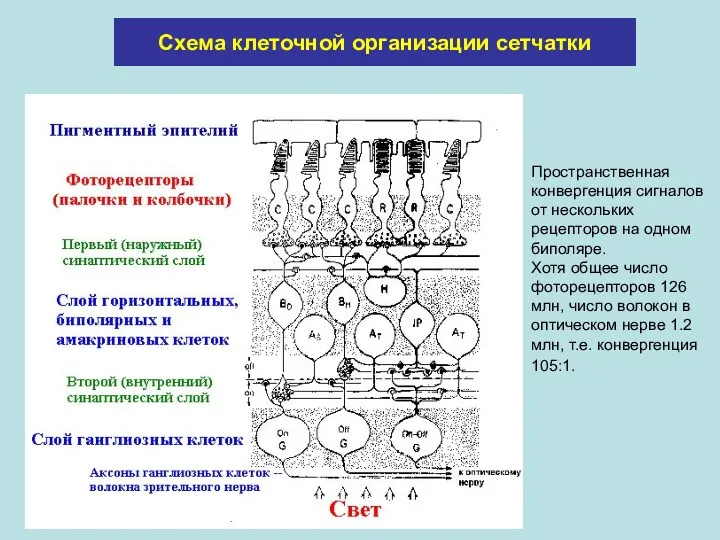
Схема клеточной организации сетчатки
Пространственная конвергенция сигналов от нескольких рецепторов на
Схема клеточной организации сетчатки
Пространственная конвергенция сигналов от нескольких рецепторов на
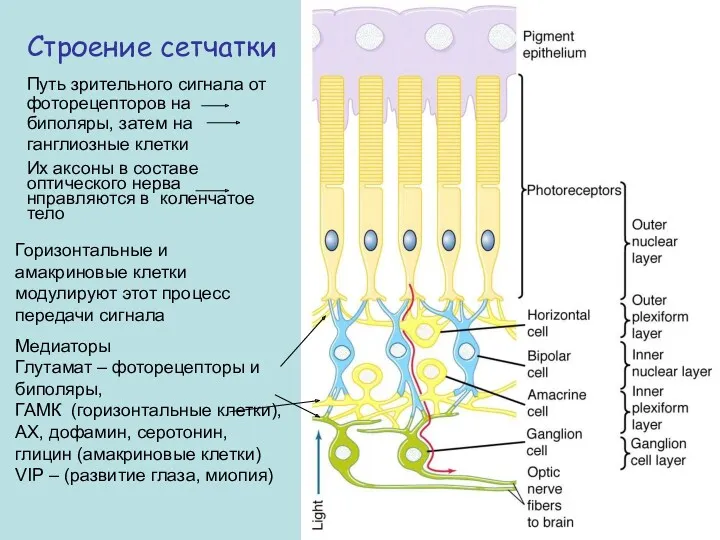
Строение сетчатки
Путь зрительного сигнала от фоторецепторов на биполяры, затем на
ганглиозные клетки
Их
Строение сетчатки
Путь зрительного сигнала от фоторецепторов на биполяры, затем на
ганглиозные клетки
Их

Палочки не связаны с ганглиозными клетками непосредственно, существуют два пути передачи
Палочки не связаны с ганглиозными клетками непосредственно, существуют два пути передачи

Существует относительная корреляция между субъективной светлотой и частотой разряда on-ганглиозных клеток
Существует относительная корреляция между субъективной светлотой и частотой разряда on-ганглиозных клеток
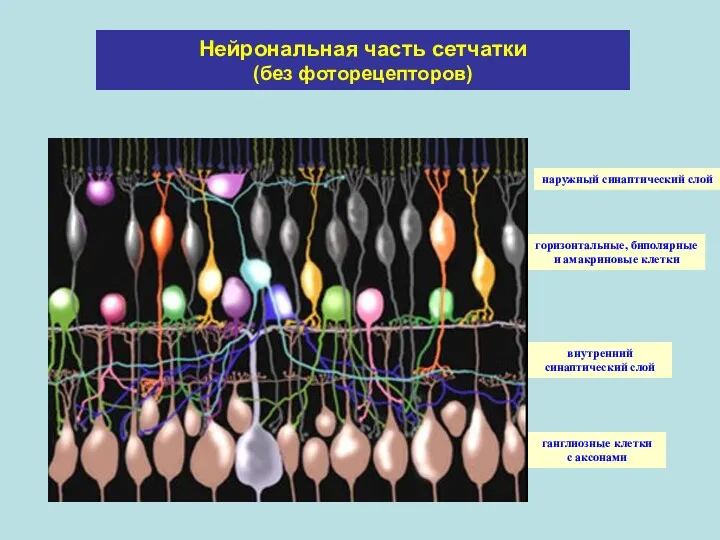
Нейрональная часть сетчатки (без фоторецепторов)
наружный синаптический слой
горизонтальные, биполярные
и амакриновые клетки
внутренний синаптический
Нейрональная часть сетчатки (без фоторецепторов)
наружный синаптический слой
горизонтальные, биполярные
и амакриновые клетки
внутренний синаптический
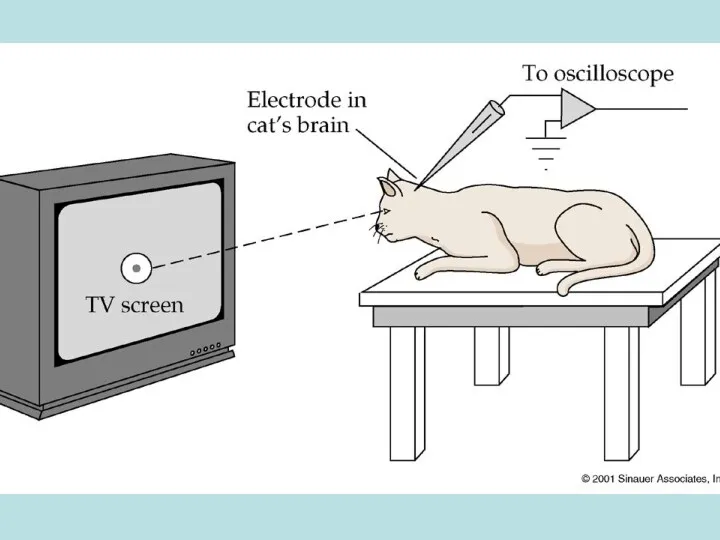
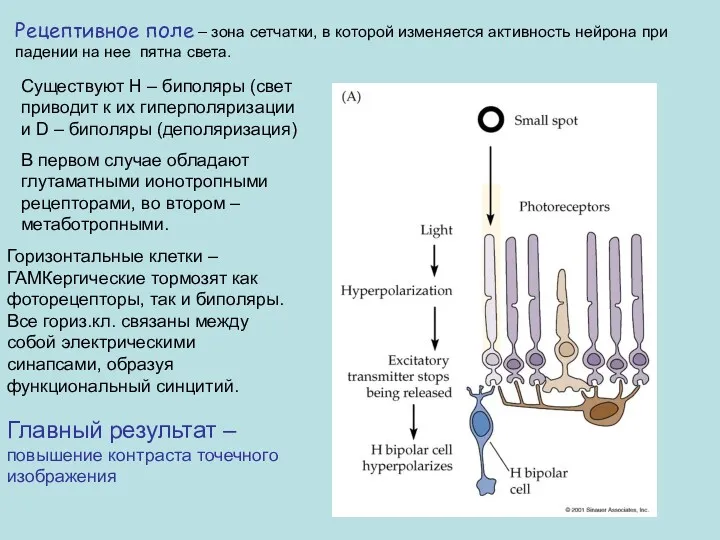
Рецептивное поле – зона сетчатки, в которой изменяется активность нейрона при
Рецептивное поле – зона сетчатки, в которой изменяется активность нейрона при
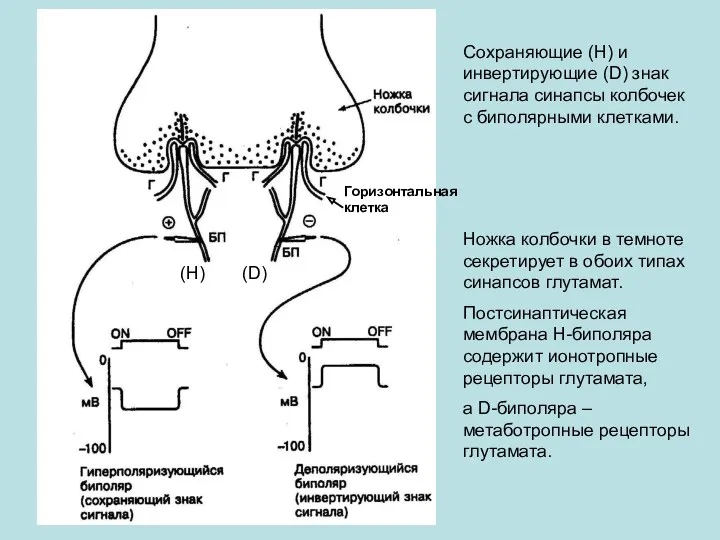
Сохраняющие (Н) и инвертирующие (D) знак сигнала синапсы колбочек с биполярными
Сохраняющие (Н) и инвертирующие (D) знак сигнала синапсы колбочек с биполярными

WHAT THE FROG'S EYE TELLS THE FROG'S BRAIN * t
Y.
WHAT THE FROG'S EYE TELLS THE FROG'S BRAIN * t
Y.
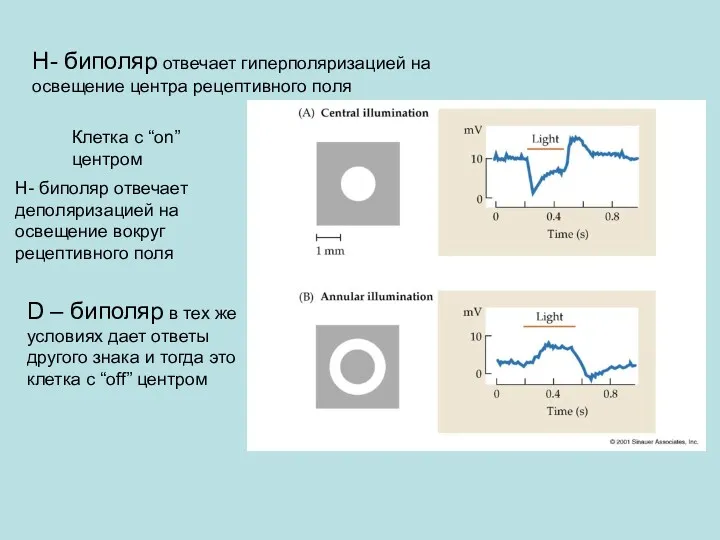
Н- биполяр отвечает гиперполяризацией на освещение центра рецептивного поля
Н- биполяр отвечает
Н- биполяр отвечает гиперполяризацией на освещение центра рецептивного поля
Н- биполяр отвечает
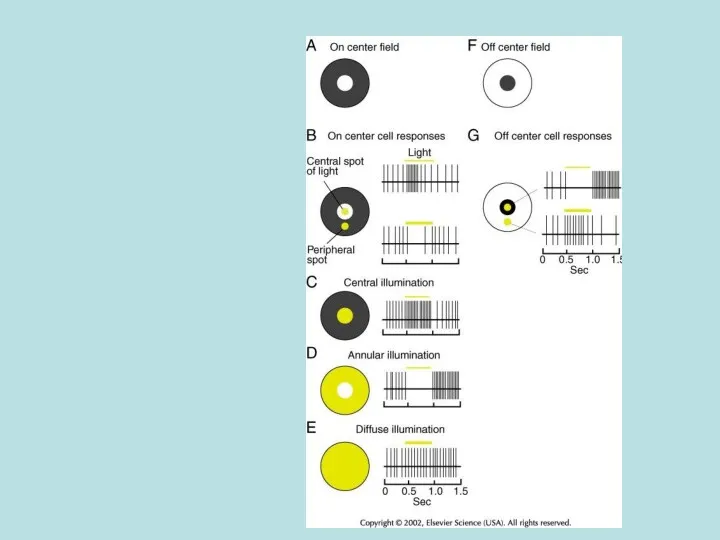
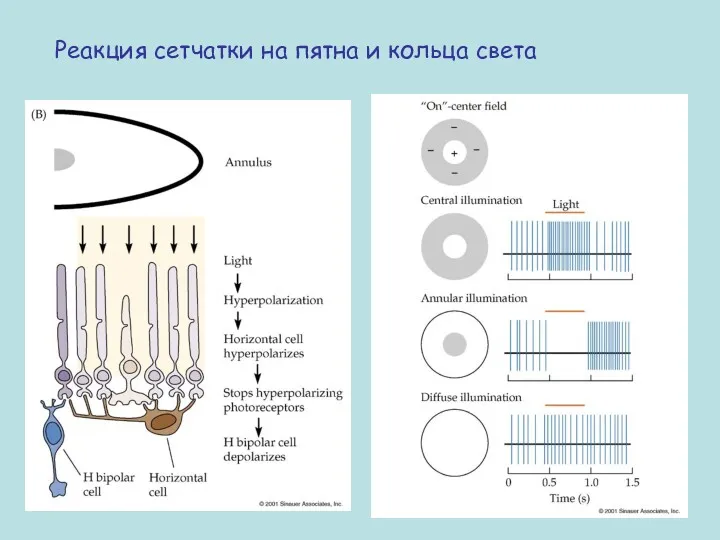
Реакция сетчатки на пятна и кольца света
Реакция сетчатки на пятна и кольца света
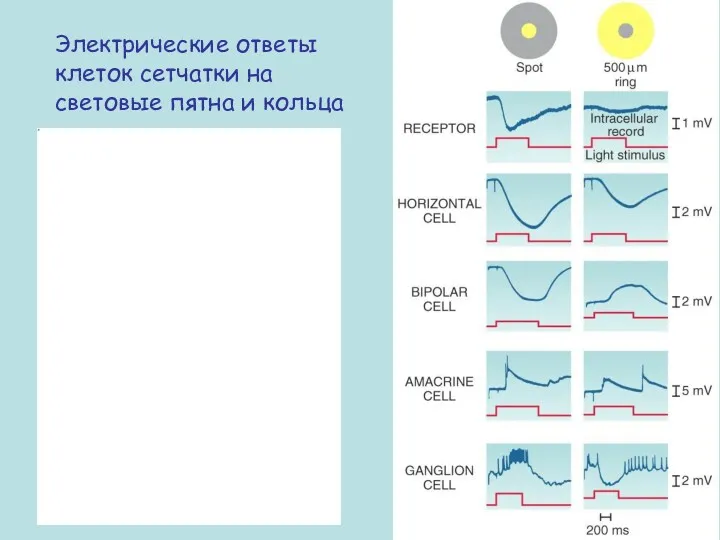
Электрические ответы клеток сетчатки на световые пятна и кольца
Электрические ответы клеток сетчатки на световые пятна и кольца
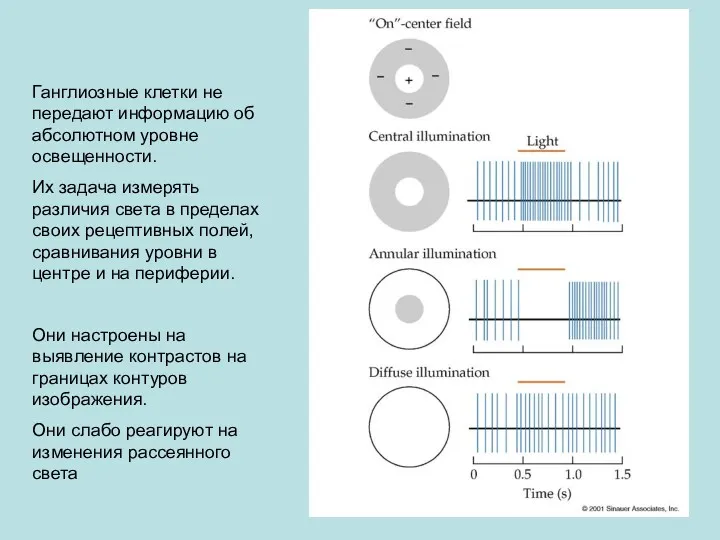
Ганглиозные клетки не передают информацию об абсолютном уровне освещенности.
Их задача измерять
Ганглиозные клетки не передают информацию об абсолютном уровне освещенности.
Их задача измерять
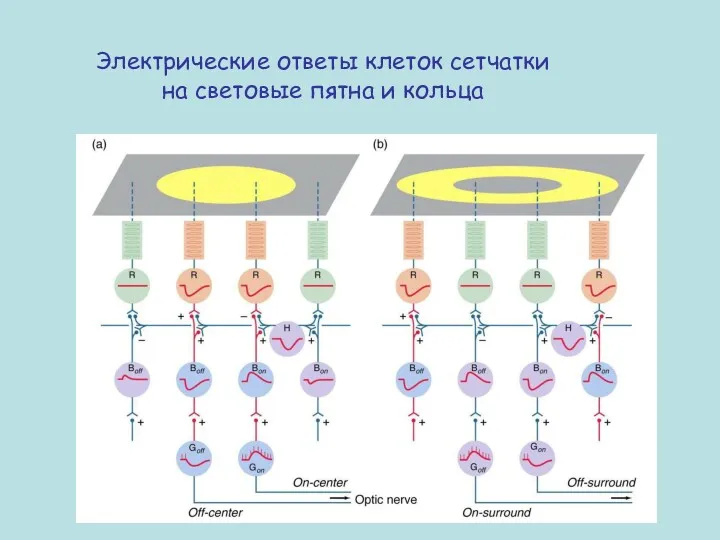
Электрические ответы клеток сетчатки на световые пятна и кольца
Электрические ответы клеток сетчатки на световые пятна и кольца
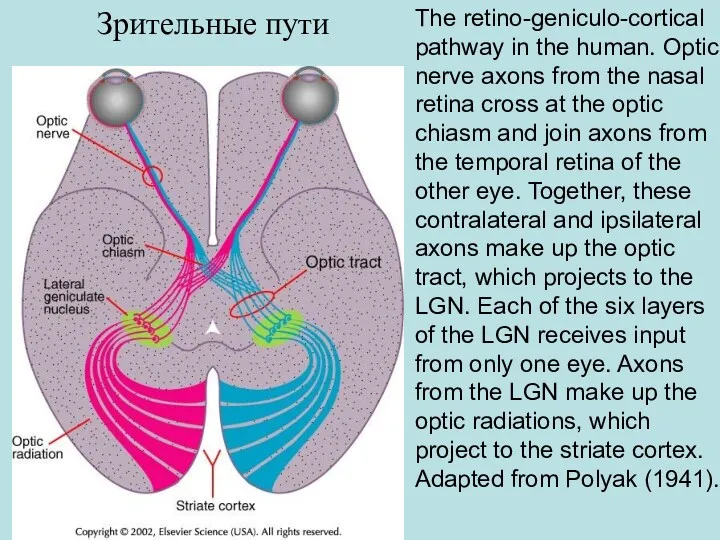
The retino-geniculo-cortical pathway in the human. Optic nerve axons from the
The retino-geniculo-cortical pathway in the human. Optic nerve axons from the
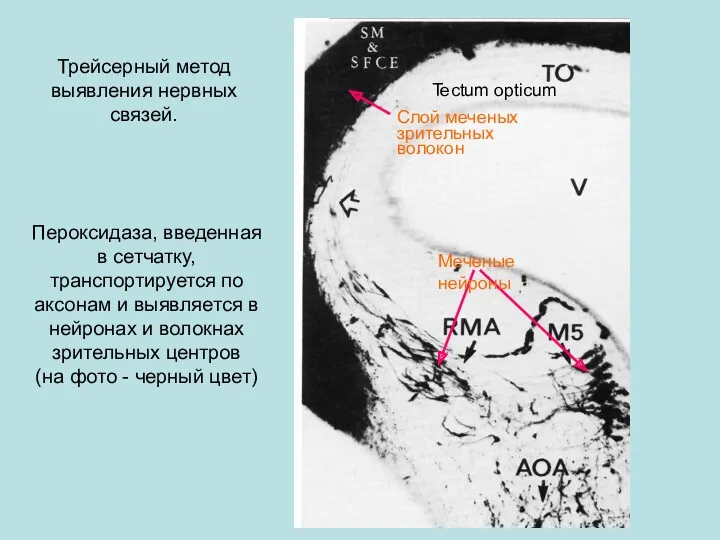
Трейсерный метод выявления нервных связей.
Пероксидаза, введенная в сетчатку, транспортируется по
Трейсерный метод выявления нервных связей.
Пероксидаза, введенная в сетчатку, транспортируется по
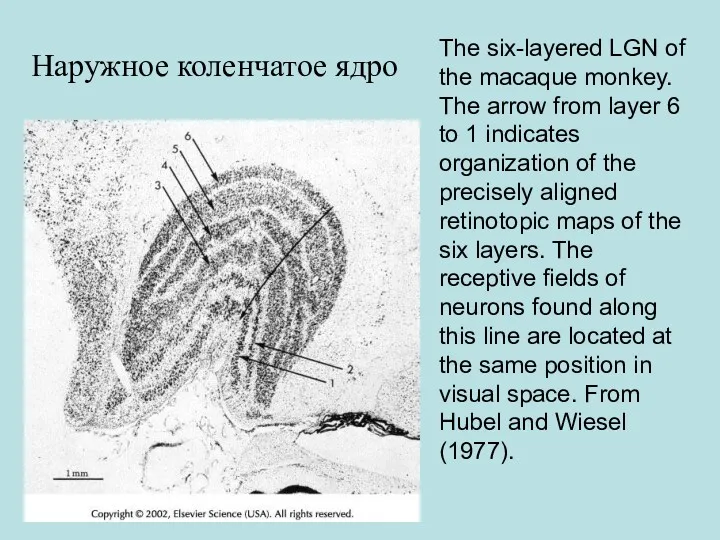
The six-layered LGN of the macaque monkey. The arrow from layer
The six-layered LGN of the macaque monkey. The arrow from layer
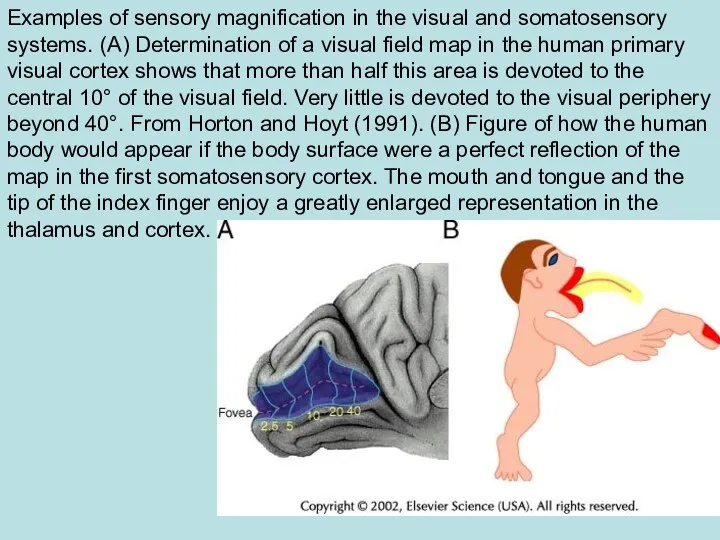
Examples of sensory magnification in the visual and somatosensory systems. (A)
Examples of sensory magnification in the visual and somatosensory systems. (A)

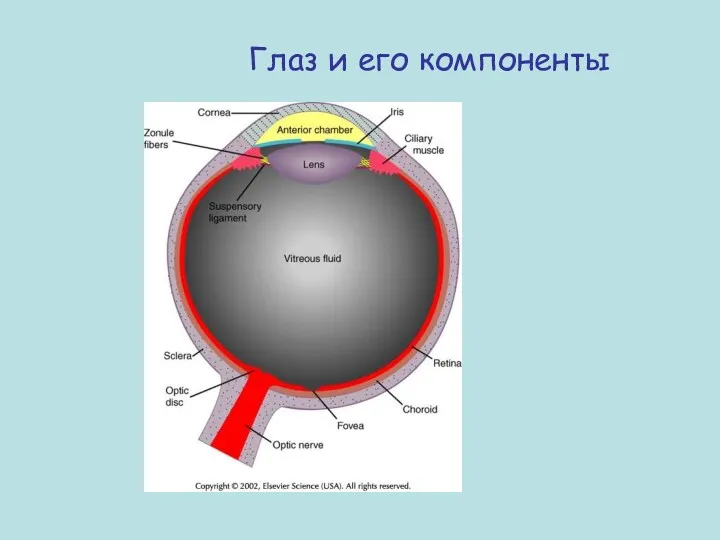
Глаз и его компоненты
Глаз и его компоненты
 Разнообразие природы родного края
Разнообразие природы родного края Большие пищеварительные железы. Функции печени
Большие пищеварительные железы. Функции печени Функциональная система процесса пищеварения
Функциональная система процесса пищеварения Синапс. Классификация синапсов
Синапс. Классификация синапсов Овощи и фрукты
Овощи и фрукты Биотические факторы в биоценозе
Биотические факторы в биоценозе Отдаленные последствия радиационного облучения
Отдаленные последствия радиационного облучения Живой мир Австралии
Живой мир Австралии Введение в курс физиологии. Возбудимые ткани. Биопотенциалы
Введение в курс физиологии. Возбудимые ткани. Биопотенциалы Тип Членистоногие, класс Ракообразные
Тип Членистоногие, класс Ракообразные Отряд Китообразные
Отряд Китообразные Взаимодействие генов
Взаимодействие генов Группы животных
Группы животных Все о кошках
Все о кошках Домашний доктор
Домашний доктор Сосуды малого круга кровообращения. Кровообращение плода
Сосуды малого круга кровообращения. Кровообращение плода Иглокожие. Лекция 15
Иглокожие. Лекция 15 Почва. Основа образования почвы
Почва. Основа образования почвы Өсімдіктер әлемінің жүйесі
Өсімдіктер әлемінің жүйесі Пейте дети молоко - будите здоровыми. Исследовательская работа
Пейте дети молоко - будите здоровыми. Исследовательская работа головоногия моллюски
головоногия моллюски 28 интересных фактов из жизни синих китов
28 интересных фактов из жизни синих китов Генетика. Первый закон Менделя
Генетика. Первый закон Менделя Обмен веществ и энергии
Обмен веществ и энергии Соединительные ткани
Соединительные ткани Покрытосеменные растения. 5 класс
Покрытосеменные растения. 5 класс Кора надпочечников
Кора надпочечников NGS - Секвенирование нового поколения
NGS - Секвенирование нового поколения