- Симбиоз и симбиогенез организмов

Содержание
- 2. Симбиоз это совместная жизнь, сосуществование, систематически отдалённых друг от друга организмов. Часто партнёры симбиоза относятся к
- 3. В начале двадцатого века и на протяжении более полувека эта теория не принималась биологами. Мережковский утверждал,
- 4. На основании собственных наблюдений и данных литературы К.С. Мережковский утверждал, что хроматофоры (хлоропласты) нельзя рассматривать как
- 5. В одной из своих публикаций Мережковский писал: «Царство микоидов (теперь прокариотов), единственное царство, которое не является
- 6. Как было отмечено выше, выводы Маргулис нельзя считать пионерскими, поскольку к аналогичным заключениям русские учёные пришли
- 7. 2). Способ деления пластид и митохондрий в клетке прокариот подобен бинарному делению бактерий. Невозможность образования пластид
- 8. Формы проявления симбиоза Учение о симбиозе при более широком взгляде можно рассматривать как составную часть экологии,
- 9. Симбиоз – это явно установленные связи представителей двух или более видов, предполагающие тесное или факультативное сожительство
- 10. Конкуренция. Эта форма симбиоза весьма широко распространена в природе. Она происходит там, где пищевые ресурсы или
- 11. Межвидовая конкуренция. В мире растений такая форма конкуренции с первого взгляда не кажется очевидной, но она
- 12. Мутуали́зм (англ. mutual – взаимный) – широко распространённая форма взаимополезного сожительства, когда присутствие партнёра становится обязательным
- 13. Высоко в кронах деревьев в тропическом лесу Юго-Восточной Азии произрастают виды мирмекодии (Myrmecodia). Род растений семейства
- 14. Самая тесная форма мутуализма – когда один организм живёт внутри другого. Поразительным примером этого служит система
- 15. Примером комменсализма у растений могут служить бобовые (например, клевер) и злаки, совместно произрастающие на почвах, бедных
- 16. Паразити́зм (от др.-греч. παράσιτος – нахлебник) – один из типов сосуществования организмов. Это явление, при котором
- 17. Факультативные паразиты, как правило, ведут свободный образ жизни и лишь при особых условиях переходят к паразитическому
- 18. Отношения между паразитом и хозяином в растительном и животном мире на популяционном и видовом уровнях определённым
- 19. Хищничество. Хищничество это форма трофических взаимоотношений между организмами разных видов, при которых один из них (хищник)
- 20. Примеры симбиоза Микориза Некоторые грибы играют ключевую роль в минеральном питании сосудистых растений. Всходы многих видов
- 21. Эктомикориза на корне проростка лиственницы сибирской. Сами, ещё слабо развитые, корни практически не видны в массе
- 22. Грибной компонент относится в основном к зигомицетам, причем во всем мире этот симбиоз образует менее 100
- 23. Типы микоризы: эктомикориза у дуба (Quercus sp) (1, 2); бука (Fagus sp.) (3) – гифы гриба
- 24. Микориза вересковых, по-видимому, необходима для снабжения растения скорее азотом, чем фосфором, что особо важно на кислых
- 25. Лишайники Лиша́йники (лат. Lichenes) – симбиотические ассоциации грибов (микобионт) и микроскопических зелёных водорослей и/или цианобактерий (фотобионт,
- 26. Первым на симбиотическую природу одного из видов лишайника в 1866 году указал врач и миколог Антон
- 27. Иллюстрация из книги Эрнста Геккеля «Kunstformen der Natur», 1904 год
- 28. Микобионт лишайника имеет полифилетическое происхождение от различных ветвей грибов. Лишайники, состоящие из гриба одного вида и
- 29. Лишайники окрашены в широком диапазоне цветов: от белого до ярко-жёлтого, коричневого, сиреневого, оранжевого, розового, зелёного, синего,
- 30. Строение гетеромерного лишайника на примере Sticta fuliginosa: a – корковый слой, b – гонидиальный слой, c
- 31. Лишайников с гетеромерным талломом большинство. В гетеромерном талломе верхний слой – корковый, сложенный гифами гриба. Он
- 32. Время, которое лишайник может провести высушенным, зависит от вида, известны случаи «воскрешения» после 40 лет в
- 33. Микобионт размножается всеми способами и в то время, когда фотобионт не размножается или размножается вегетативно. Микобионт
- 34. Плодовые тела (апотеции) у накипного лишайника: 1 – внешний вид; 2 – поперечный разрез через апотеций
- 36. Клубеньковые бактерии Клубеньковые бактерии это группа бактерий порядка Rhizobiales, способных связывать неорганический атмосферный азот, продуцируя органические
- 37. В целинных и окультуренных почвах присутствуют обычно в больших количествах клубеньковые бактерии тех видов бобовых растений,
- 38. Клубеньки на корнях ольхи чёрной (Alnus glutinosa) Клубеньки в корневой системе Lotus pedunculatus сем. бобовые Клубеньковые
- 39. Клубеньковые бактерии – микроаэрофилы (развиваются при незначительных количествах кислорода в среде), предпочитающие, однако, аэробные условия. Клубеньковые
- 40. Costasiella kuroshimae (морская овечка) Elysia-chlorotica-body На рисунках показаны яркие, но не часто обсуждаемые в научной литературе,
- 41. Costasiella kuroshimae (морская овечка) – брюхонгий моллюск. Этот слизняк, обитающий в Юго-Восточной Азиина глубине 10 метров.
- 43. Скачать презентацию

Симбиоз это совместная жизнь, сосуществование, систематически отдалённых друг от друга организмов.
Симбиоз это совместная жизнь, сосуществование, систематически отдалённых друг от друга организмов.

В начале двадцатого века и на протяжении более полувека эта теория
В начале двадцатого века и на протяжении более полувека эта теория

На основании собственных наблюдений и данных литературы К.С. Мережковский утверждал, что
На основании собственных наблюдений и данных литературы К.С. Мережковский утверждал, что

В одной из своих публикаций Мережковский писал: «Царство микоидов (теперь прокариотов),
В одной из своих публикаций Мережковский писал: «Царство микоидов (теперь прокариотов),

Как было отмечено выше, выводы Маргулис нельзя считать пионерскими, поскольку к
Как было отмечено выше, выводы Маргулис нельзя считать пионерскими, поскольку к

2). Способ деления пластид и митохондрий в клетке прокариот подобен бинарному
2). Способ деления пластид и митохондрий в клетке прокариот подобен бинарному

Формы проявления симбиоза
Учение о симбиозе при более широком взгляде можно рассматривать
Формы проявления симбиоза
Учение о симбиозе при более широком взгляде можно рассматривать

Симбиоз – это явно установленные связи представителей двух или более видов,
Симбиоз – это явно установленные связи представителей двух или более видов,

Конкуренция. Эта форма симбиоза весьма широко распространена в природе. Она происходит
Конкуренция. Эта форма симбиоза весьма широко распространена в природе. Она происходит

Межвидовая конкуренция. В мире растений такая форма конкуренции с первого взгляда
Межвидовая конкуренция. В мире растений такая форма конкуренции с первого взгляда

Мутуали́зм (англ. mutual – взаимный) – широко распространённая форма взаимополезного сожительства,
Мутуали́зм (англ. mutual – взаимный) – широко распространённая форма взаимополезного сожительства,

Высоко в кронах деревьев в тропическом лесу Юго-Восточной Азии произрастают виды
Высоко в кронах деревьев в тропическом лесу Юго-Восточной Азии произрастают виды

Самая тесная форма мутуализма – когда один организм живёт внутри другого.
Самая тесная форма мутуализма – когда один организм живёт внутри другого.

Примером комменсализма у растений могут служить бобовые (например, клевер) и злаки,
Примером комменсализма у растений могут служить бобовые (например, клевер) и злаки,

Паразити́зм (от др.-греч. παράσιτος – нахлебник) – один из типов сосуществования
Паразити́зм (от др.-греч. παράσιτος – нахлебник) – один из типов сосуществования

Факультативные паразиты, как правило, ведут свободный образ жизни и лишь при
Факультативные паразиты, как правило, ведут свободный образ жизни и лишь при

Отношения между паразитом и хозяином в растительном и животном мире на
Отношения между паразитом и хозяином в растительном и животном мире на

Хищничество. Хищничество это форма трофических взаимоотношений между организмами разных видов, при
Хищничество. Хищничество это форма трофических взаимоотношений между организмами разных видов, при

Примеры симбиоза
Микориза
Некоторые грибы играют ключевую роль в минеральном питании сосудистых растений.
Примеры симбиоза
Микориза
Некоторые грибы играют ключевую роль в минеральном питании сосудистых растений.

Эктомикориза на корне проростка лиственницы сибирской. Сами, ещё слабо развитые, корни
Эктомикориза на корне проростка лиственницы сибирской. Сами, ещё слабо развитые, корни

Грибной компонент относится в основном к зигомицетам, причем во всем мире
Грибной компонент относится в основном к зигомицетам, причем во всем мире

Типы микоризы: эктомикориза у дуба (Quercus sp) (1, 2); бука (Fagus
Типы микоризы: эктомикориза у дуба (Quercus sp) (1, 2); бука (Fagus

Микориза вересковых, по-видимому, необходима для снабжения растения скорее азотом, чем фосфором,
Микориза вересковых, по-видимому, необходима для снабжения растения скорее азотом, чем фосфором,

Лишайники
Лиша́йники (лат. Lichenes) – симбиотические ассоциации грибов (микобионт) и микроскопических зелёных
Лишайники
Лиша́йники (лат. Lichenes) – симбиотические ассоциации грибов (микобионт) и микроскопических зелёных

Первым на симбиотическую природу одного из видов лишайника в 1866 году
Первым на симбиотическую природу одного из видов лишайника в 1866 году
Иллюстрация из книги Эрнста Геккеля «Kunstformen der Natur», 1904 год
Иллюстрация из книги Эрнста Геккеля «Kunstformen der Natur», 1904 год

Микобионт лишайника имеет полифилетическое происхождение от различных ветвей грибов.
Лишайники, состоящие из
Микобионт лишайника имеет полифилетическое происхождение от различных ветвей грибов.
Лишайники, состоящие из

Лишайники окрашены в широком диапазоне цветов: от белого до ярко-жёлтого, коричневого,
Лишайники окрашены в широком диапазоне цветов: от белого до ярко-жёлтого, коричневого,
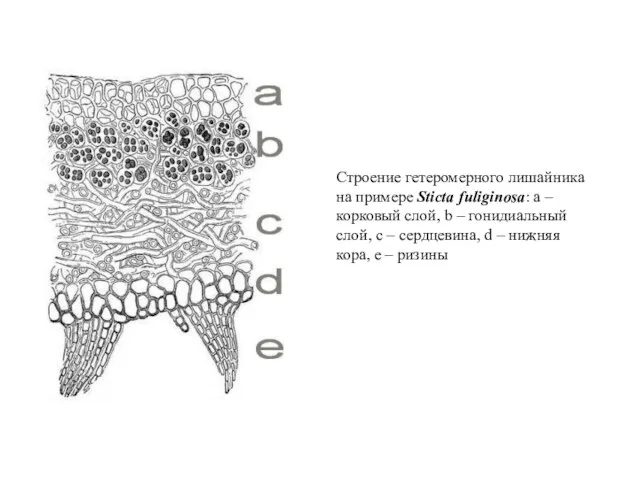
Строение гетеромерного лишайника на примере Sticta fuliginosa: a – корковый слой,
Строение гетеромерного лишайника на примере Sticta fuliginosa: a – корковый слой,

Лишайников с гетеромерным талломом большинство. В гетеромерном талломе верхний слой –
Лишайников с гетеромерным талломом большинство. В гетеромерном талломе верхний слой –

Время, которое лишайник может провести высушенным, зависит от вида, известны случаи
Время, которое лишайник может провести высушенным, зависит от вида, известны случаи

Микобионт размножается всеми способами и в то время, когда фотобионт не
Микобионт размножается всеми способами и в то время, когда фотобионт не
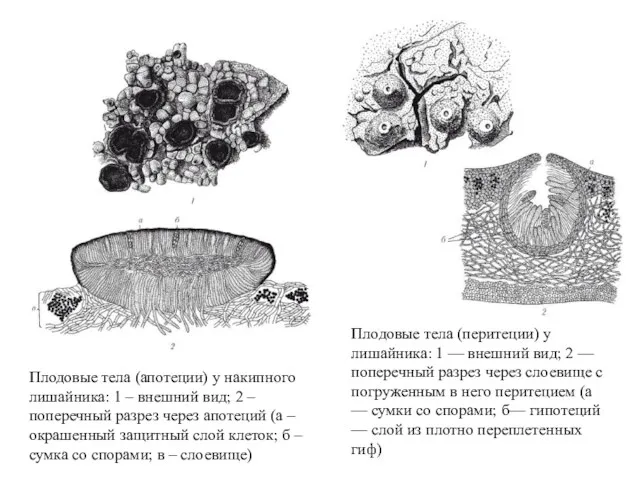
Плодовые тела (апотеции) у накипного лишайника: 1 – внешний вид; 2
Плодовые тела (апотеции) у накипного лишайника: 1 – внешний вид; 2


Клубеньковые бактерии
Клубеньковые бактерии это группа бактерий порядка Rhizobiales, способных связывать неорганический
Клубеньковые бактерии
Клубеньковые бактерии это группа бактерий порядка Rhizobiales, способных связывать неорганический

В целинных и окультуренных почвах присутствуют обычно в больших количествах клубеньковые
В целинных и окультуренных почвах присутствуют обычно в больших количествах клубеньковые
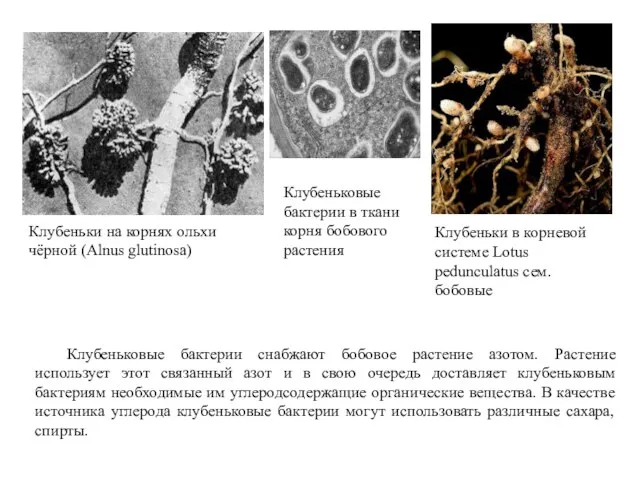
Клубеньки на корнях ольхи чёрной (Alnus glutinosa)
Клубеньки в корневой системе Lotus
Клубеньки на корнях ольхи чёрной (Alnus glutinosa)
Клубеньки в корневой системе Lotus

Клубеньковые бактерии – микроаэрофилы (развиваются при незначительных количествах кислорода в среде),
Клубеньковые бактерии – микроаэрофилы (развиваются при незначительных количествах кислорода в среде),

Costasiella kuroshimae (морская овечка)
Elysia-chlorotica-body
На рисунках показаны яркие, но не часто обсуждаемые
Costasiella kuroshimae (морская овечка)
Elysia-chlorotica-body
На рисунках показаны яркие, но не часто обсуждаемые

Costasiella kuroshimae (морская овечка) – брюхонгий моллюск. Этот слизняк, обитающий в
Costasiella kuroshimae (морская овечка) – брюхонгий моллюск. Этот слизняк, обитающий в
 Мутагенез. Механизмы защиты генома от мутагенных воздействий
Мутагенез. Механизмы защиты генома от мутагенных воздействий Метаболические основы физиологических функций. Лекция 28
Метаболические основы физиологических функций. Лекция 28 Опора і рух. Види скелета. Значення опорно-рухової системи
Опора і рух. Види скелета. Значення опорно-рухової системи Легенды и сказания о цветах
Легенды и сказания о цветах Рыба-клоун
Рыба-клоун Интересные факты о змеях
Интересные факты о змеях Минеральное питание растений
Минеральное питание растений Липиды. Классификация, физические и химические свойства
Липиды. Классификация, физические и химические свойства Презентация Грибы
Презентация Грибы Электронный атлас растений
Электронный атлас растений Домашние животные
Домашние животные Факторы среды
Факторы среды Вода - источник жизни на Земле
Вода - источник жизни на Земле Мутагены, их влияние на наследственный аппарат человека
Мутагены, их влияние на наследственный аппарат человека Животноводство
Животноводство Регуляция дыхания
Регуляция дыхания Презентация к уроку Анализаторы
Презентация к уроку Анализаторы Вроджені системні захворювання скелету
Вроджені системні захворювання скелету Акушерская физиология. Анатомия и физиология половых органов самцов животных
Акушерская физиология. Анатомия и физиология половых органов самцов животных Презентация по биологии для учащихся 9 класса по теме Деление клетки. Митоз
Презентация по биологии для учащихся 9 класса по теме Деление клетки. Митоз Углеводы и их роль в жизнедеятельности клетки
Углеводы и их роль в жизнедеятельности клетки Оплодотворение у цветковых растений
Оплодотворение у цветковых растений Путешествие по материкам. Животный мир Евразии
Путешествие по материкам. Животный мир Евразии Мозговая организация слухового восприятия
Мозговая организация слухового восприятия Основные оложения клеточной теории. 10 кл.
Основные оложения клеточной теории. 10 кл. Класс млекопитающие, отряд китообразные
Класс млекопитающие, отряд китообразные Пластиковая чума
Пластиковая чума Зміни в м᾽ясі при тепловій обробці. Лекція 10,11
Зміни в м᾽ясі при тепловій обробці. Лекція 10,11