- Синтез нуклеотидов. Происхождение хиральной чистоты

Содержание
- 2. ЧТО НАМ СТОИТ РНК ПОСТРОИТЬ ? Азотистые основания (аденин, гуанин, цитозин, урацил) Рибоза Фосфат Соединить их
- 3. РЕАКЦИЯ БУТЛЕРОВА: СИНТЕЗ САХАРОВ Водный раствор формальдегида (СН2О), Са(ОН)2, небольшое нагревание Образуются все возможные сахара одновременно
- 4. СИНТЕЗ АЗОТИСТЫХ ОСНОВАНИЙ Аденин и гуанин - из HCN при замерзании водного раствора, УФ-облучении, нагревании. При
- 5. ВОССТАНОВЛЕНИЕ СО2 В ГЕОТЕРМАЛЬНЫХ СИСТЕМАХ: ЩЕЛОЧНЫЕ ИСТОЧНИКИ Температуры 40-100 'C, pH 10-11 Осаждение Mg(OH)2 и CaCO3
- 6. ИСТОЧНИКИ ФОРМАЛЬДЕГИДА И ЦИАНИДА: АТМОСФЕРНАЯ ФОТОХИМИЯ
- 7. СИНТЕЗ НУКЛЕОТИДОВ Аденин при УФ-облучении водного раствора присоединяет рибозу и до 3 фосфатных групп Гуанин присоединяет
- 8. ОБХОДНОЙ ПУТЬ СИНТЕЗА ЦИТИДИНОВЫХ И ТИМИДИНОВЫХ НУКЛЕОТИДОВ Фосфат катализирует нужные реакции Побочные продукты первых реакций катализируют
- 9. СИНТЕЗ АКТИВИРОВАННЫХ АДЕНИНОВЫХ И ГУАНИНОВЫХ НУКЛЕОТИДОВ Для получения пуринов в той же системе достаточно добавить синильную
- 10. СИНТЕЗ ГЛИКОАЛЬДЕГИДА И ГЛИЦЕРАЛЬДЕГИДА: РЕАКЦИЯ КИЛИАНИ-ФИШЕРА На древней Земле не было Pd / BaSO4 Подходящий природный
- 11. ЦИАНОСУЛЬФИДНЫЙ ПРОТОМЕТАБОЛИЗМ Фосфат вызывает побочные реакции: Глицеральдегид → диоксиацетон → Val, Leu Цианоацетилен → малеонитрил →
- 12. ФОРМАМИДНЫЙ МИР Из формамида легко образуются азотистые основания Из формамида на TiO2 образуются N-формил-производные азотистых оснований,
- 13. МЕХАНИЗМЫ ПОЯВЛЕНИЯ ХИРАЛЬНОЙ ЧИСТОТЫ В метеоритной органике есть преобладание левых (L) аминокислот над правыми (D) Избирательное
- 14. МЕХАНИЗМЫ УСИЛЕНИЯ ХИРАЛЬНОЙ ЧИСТОТЫ Аминокислоты обычно выпадают в осадок в соотношении L:D 1:1 – раствор обогащается
- 15. СИНТЕЗ ХИРАЛЬНО ЧИСТЫХ НУКЛЕОТИДОВ L-аминокислоты связывают L-глицеральдегид в побочную реакцию Из D-глицеральдегида образуются нуклеотиды с D-рибозой
- 16. КОПИРОВАНИЕ РНК НАЧАЛО РНК-МИРА Лекция 6
- 17. БЕЗМАТРИЧНАЯ ПОЛИМЕРИЗАЦИЯ НУКЛЕОТИДОВ В РНК На поверхности минералов (глины, сульфиды металлов) из имидазол-активированных нуклеотидов — до
- 18. РИБОЗИМЫ — РНК-ПОЛИМЕРАЗЫ Активные рибозимы-полимеразы состоят из 150-200 нуклеотидов Лучшая полимераза tC19Z во льду удлиняет праймер
- 19. РИБОЗИМЫ — РНК-ЛИГАЗА Гораздо короче полимераз, от 29 нуклеотидов → вероятнее случайное возникновение из абиогенных шпилек
- 20. АБИОГЕННАЯ РЕПЛИКАЦИЯ РНК При высокой концентрации Mg++ короткие РНК реплицируются сами Копируется до 15 нуклеотидов 10%
- 21. НАКОПЛЕНИЕ РНК В ТЕПЛОВОЙ ЛОВУШКЕ Конвекция и термофорез в капиллярах концентрируют РНК и нуклеотиды на много
- 22. МИР ПАЛИНДРОМОВ Дарвиновская эволюция начинается до первого рибозима — отбор идет на самопрайминг Палиндромные РНК богаты
- 24. Скачать презентацию
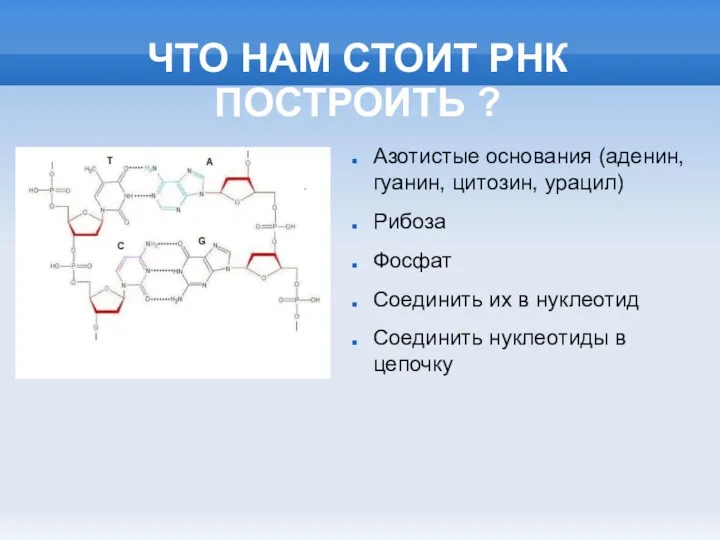
ЧТО НАМ СТОИТ РНК ПОСТРОИТЬ ?
Азотистые основания (аденин, гуанин, цитозин, урацил)
Рибоза
Фосфат
Соединить
ЧТО НАМ СТОИТ РНК ПОСТРОИТЬ ?
Азотистые основания (аденин, гуанин, цитозин, урацил)
Рибоза
Фосфат
Соединить
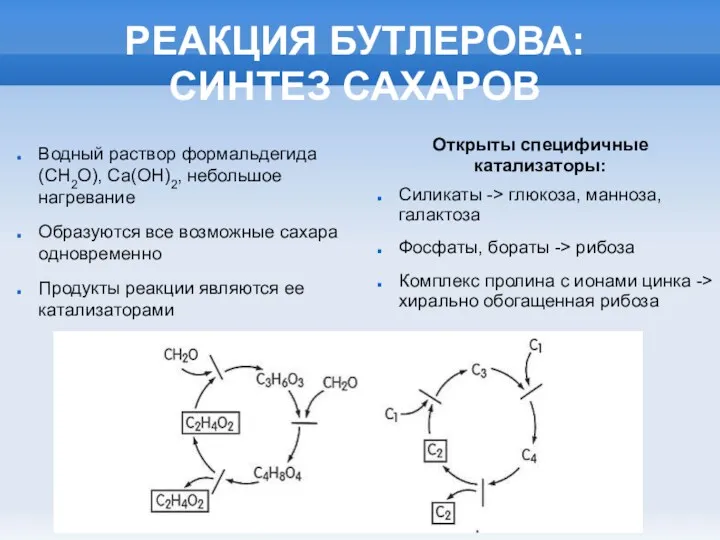
РЕАКЦИЯ БУТЛЕРОВА: СИНТЕЗ САХАРОВ
Водный раствор формальдегида (СН2О), Са(ОН)2, небольшое нагревание
Образуются все
РЕАКЦИЯ БУТЛЕРОВА: СИНТЕЗ САХАРОВ
Водный раствор формальдегида (СН2О), Са(ОН)2, небольшое нагревание
Образуются все

СИНТЕЗ АЗОТИСТЫХ ОСНОВАНИЙ
Аденин и гуанин - из HCN при замерзании водного
СИНТЕЗ АЗОТИСТЫХ ОСНОВАНИЙ
Аденин и гуанин - из HCN при замерзании водного
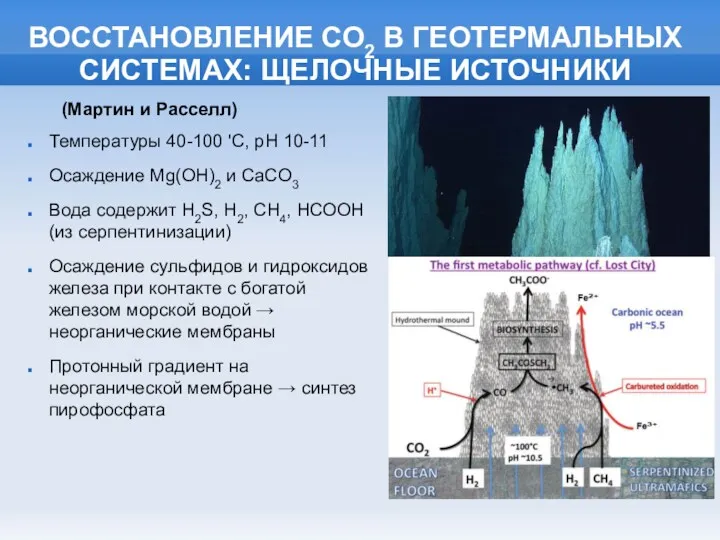
ВОССТАНОВЛЕНИЕ СО2 В ГЕОТЕРМАЛЬНЫХ СИСТЕМАХ: ЩЕЛОЧНЫЕ ИСТОЧНИКИ
Температуры 40-100 'C, pH 10-11
Осаждение
ВОССТАНОВЛЕНИЕ СО2 В ГЕОТЕРМАЛЬНЫХ СИСТЕМАХ: ЩЕЛОЧНЫЕ ИСТОЧНИКИ
Температуры 40-100 'C, pH 10-11
Осаждение
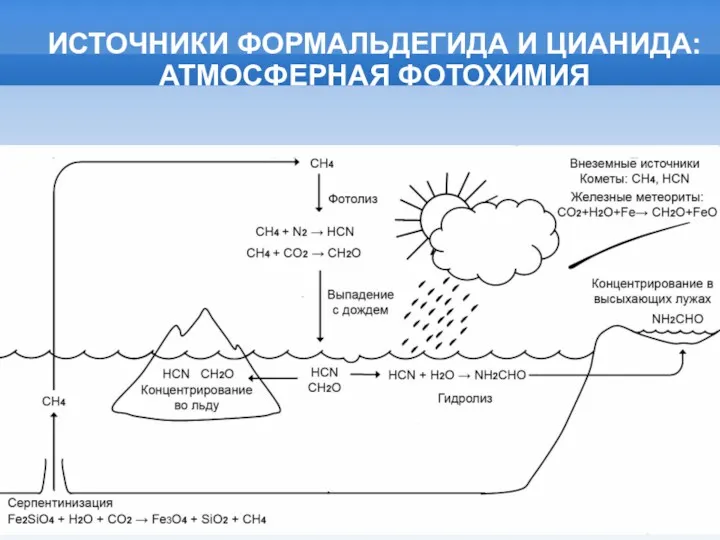
ИСТОЧНИКИ ФОРМАЛЬДЕГИДА И ЦИАНИДА:
АТМОСФЕРНАЯ ФОТОХИМИЯ
ИСТОЧНИКИ ФОРМАЛЬДЕГИДА И ЦИАНИДА:
АТМОСФЕРНАЯ ФОТОХИМИЯ

СИНТЕЗ НУКЛЕОТИДОВ
Аденин при УФ-облучении водного раствора присоединяет рибозу и до 3
СИНТЕЗ НУКЛЕОТИДОВ
Аденин при УФ-облучении водного раствора присоединяет рибозу и до 3
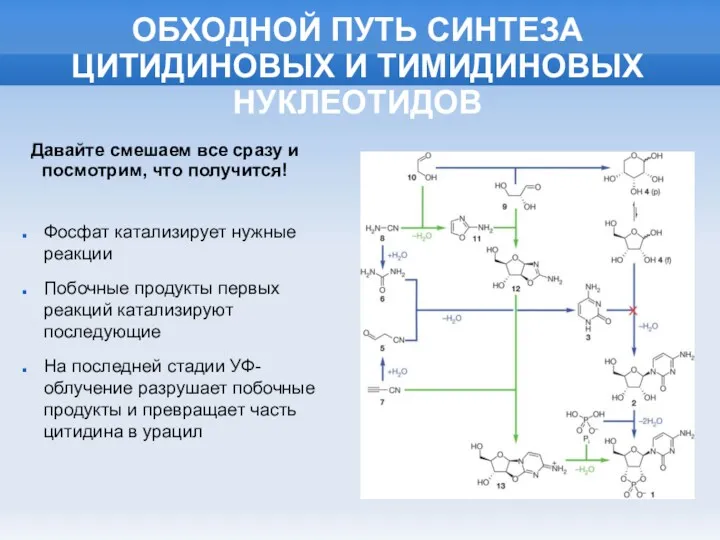
ОБХОДНОЙ ПУТЬ СИНТЕЗА ЦИТИДИНОВЫХ И ТИМИДИНОВЫХ НУКЛЕОТИДОВ
Фосфат катализирует нужные реакции
Побочные
ОБХОДНОЙ ПУТЬ СИНТЕЗА ЦИТИДИНОВЫХ И ТИМИДИНОВЫХ НУКЛЕОТИДОВ
Фосфат катализирует нужные реакции
Побочные

СИНТЕЗ АКТИВИРОВАННЫХ АДЕНИНОВЫХ И ГУАНИНОВЫХ НУКЛЕОТИДОВ
Для получения пуринов в той же
СИНТЕЗ АКТИВИРОВАННЫХ АДЕНИНОВЫХ И ГУАНИНОВЫХ НУКЛЕОТИДОВ
Для получения пуринов в той же
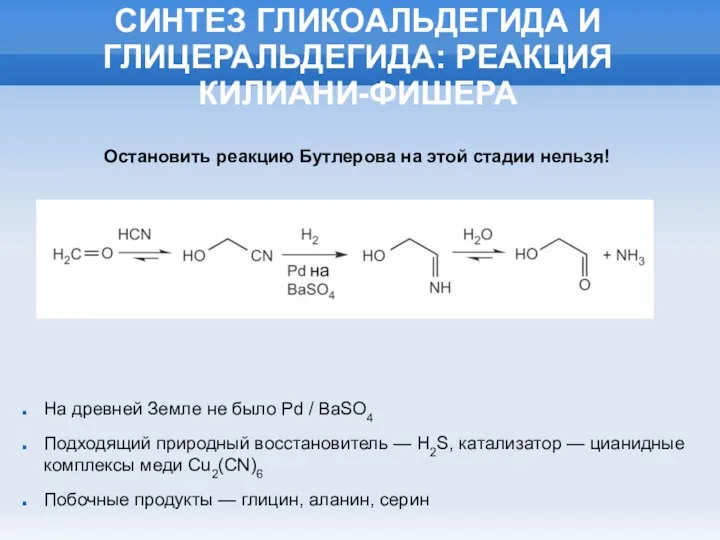
СИНТЕЗ ГЛИКОАЛЬДЕГИДА И ГЛИЦЕРАЛЬДЕГИДА: РЕАКЦИЯ КИЛИАНИ-ФИШЕРА
На древней Земле не было Pd
СИНТЕЗ ГЛИКОАЛЬДЕГИДА И ГЛИЦЕРАЛЬДЕГИДА: РЕАКЦИЯ КИЛИАНИ-ФИШЕРА
На древней Земле не было Pd
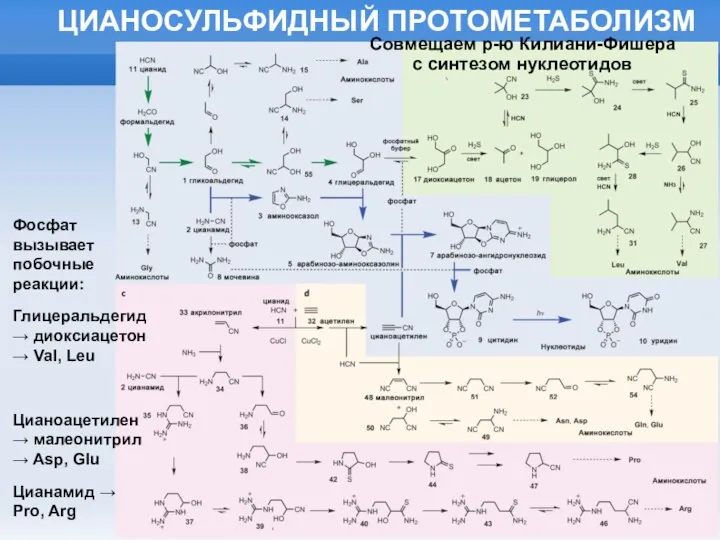
ЦИАНОСУЛЬФИДНЫЙ ПРОТОМЕТАБОЛИЗМ
Фосфат вызывает побочные реакции:
Глицеральдегид → диоксиацетон → Val, Leu
Цианоацетилен
ЦИАНОСУЛЬФИДНЫЙ ПРОТОМЕТАБОЛИЗМ
Фосфат вызывает побочные реакции:
Глицеральдегид → диоксиацетон → Val, Leu
Цианоацетилен

ФОРМАМИДНЫЙ МИР
Из формамида легко образуются азотистые основания
Из формамида на TiO2 образуются
ФОРМАМИДНЫЙ МИР
Из формамида легко образуются азотистые основания
Из формамида на TiO2 образуются

МЕХАНИЗМЫ ПОЯВЛЕНИЯ ХИРАЛЬНОЙ ЧИСТОТЫ
В метеоритной органике есть преобладание левых (L) аминокислот
МЕХАНИЗМЫ ПОЯВЛЕНИЯ ХИРАЛЬНОЙ ЧИСТОТЫ
В метеоритной органике есть преобладание левых (L) аминокислот
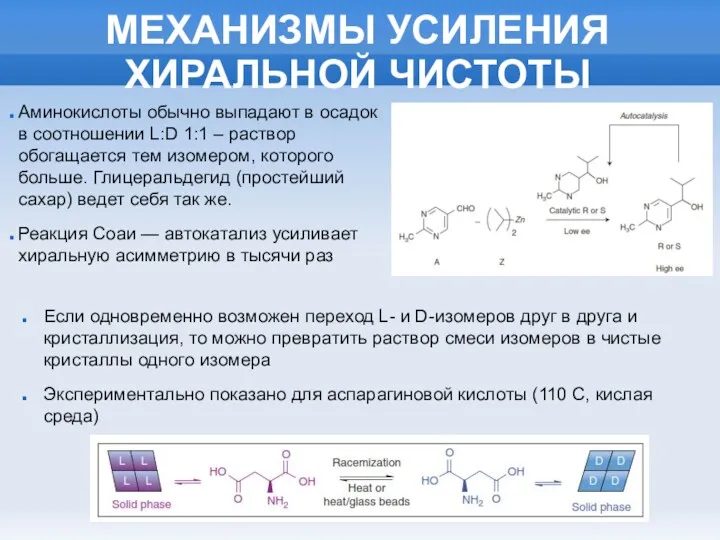
МЕХАНИЗМЫ УСИЛЕНИЯ ХИРАЛЬНОЙ ЧИСТОТЫ
Аминокислоты обычно выпадают в осадок в соотношении L:D
МЕХАНИЗМЫ УСИЛЕНИЯ ХИРАЛЬНОЙ ЧИСТОТЫ
Аминокислоты обычно выпадают в осадок в соотношении L:D
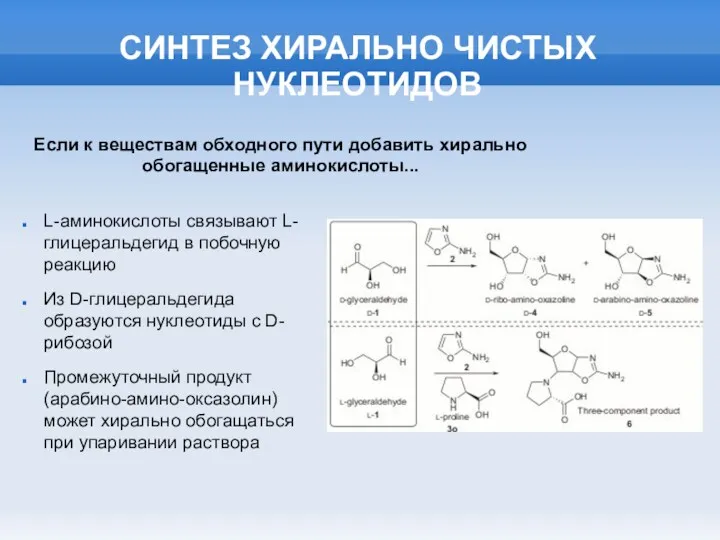
СИНТЕЗ ХИРАЛЬНО ЧИСТЫХ НУКЛЕОТИДОВ
L-аминокислоты связывают L-глицеральдегид в побочную реакцию
Из D-глицеральдегида
СИНТЕЗ ХИРАЛЬНО ЧИСТЫХ НУКЛЕОТИДОВ
L-аминокислоты связывают L-глицеральдегид в побочную реакцию
Из D-глицеральдегида

КОПИРОВАНИЕ РНК
НАЧАЛО РНК-МИРА
Лекция 6
КОПИРОВАНИЕ РНК
НАЧАЛО РНК-МИРА
Лекция 6

БЕЗМАТРИЧНАЯ ПОЛИМЕРИЗАЦИЯ НУКЛЕОТИДОВ В РНК
На поверхности минералов (глины, сульфиды металлов) из
БЕЗМАТРИЧНАЯ ПОЛИМЕРИЗАЦИЯ НУКЛЕОТИДОВ В РНК
На поверхности минералов (глины, сульфиды металлов) из

РИБОЗИМЫ — РНК-ПОЛИМЕРАЗЫ
Активные рибозимы-полимеразы состоят из 150-200 нуклеотидов
Лучшая полимераза tC19Z во
РИБОЗИМЫ — РНК-ПОЛИМЕРАЗЫ
Активные рибозимы-полимеразы состоят из 150-200 нуклеотидов
Лучшая полимераза tC19Z во
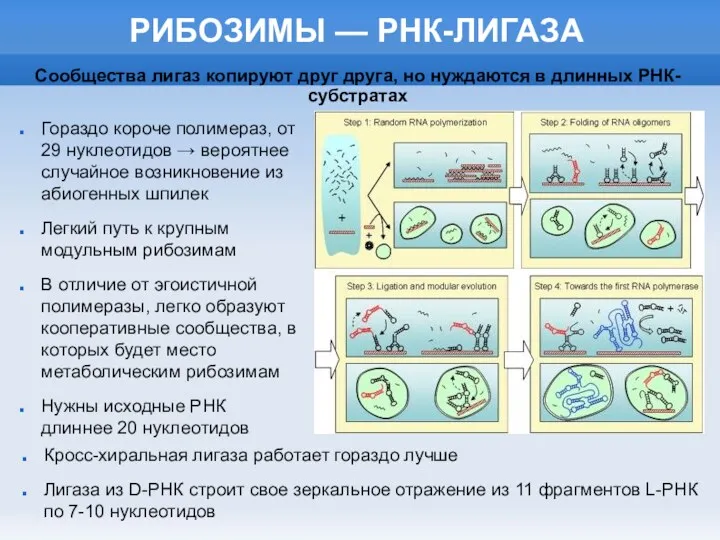
РИБОЗИМЫ — РНК-ЛИГАЗА
Гораздо короче полимераз, от 29 нуклеотидов → вероятнее случайное
РИБОЗИМЫ — РНК-ЛИГАЗА
Гораздо короче полимераз, от 29 нуклеотидов → вероятнее случайное

АБИОГЕННАЯ РЕПЛИКАЦИЯ РНК
При высокой концентрации Mg++ короткие РНК реплицируются сами
Копируется до
АБИОГЕННАЯ РЕПЛИКАЦИЯ РНК
При высокой концентрации Mg++ короткие РНК реплицируются сами
Копируется до

НАКОПЛЕНИЕ РНК В ТЕПЛОВОЙ ЛОВУШКЕ
Конвекция и термофорез в капиллярах концентрируют РНК
НАКОПЛЕНИЕ РНК В ТЕПЛОВОЙ ЛОВУШКЕ
Конвекция и термофорез в капиллярах концентрируют РНК
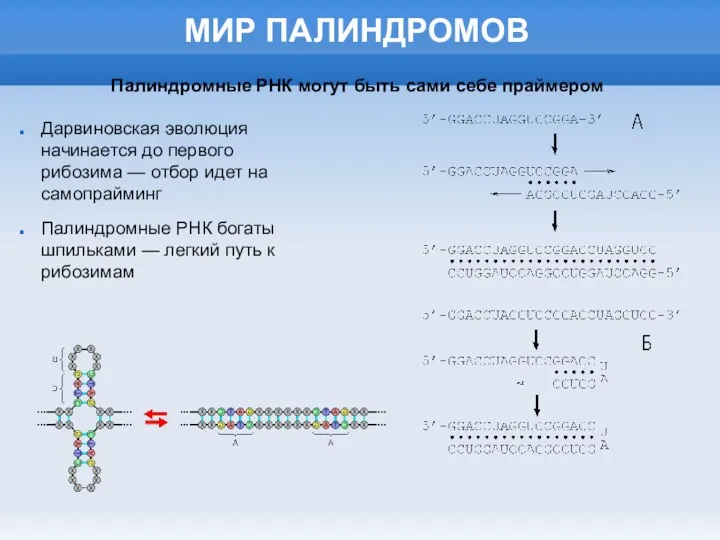
МИР ПАЛИНДРОМОВ
Дарвиновская эволюция начинается до первого рибозима — отбор идет на
МИР ПАЛИНДРОМОВ
Дарвиновская эволюция начинается до первого рибозима — отбор идет на
 Биоэлектрогенез. Строение и физические свойства биологических мембран
Биоэлектрогенез. Строение и физические свойства биологических мембран Пищеварительная система
Пищеварительная система Структурная организация микробной клетки
Структурная организация микробной клетки Биотехнология как источник био-риска
Биотехнология как источник био-риска Фізіологія. Механізми регуляції
Фізіологія. Механізми регуляції Систематика Покритонасінних рослин. Відділ Магноліофіти, Квіткові або Покритонасінні
Систематика Покритонасінних рослин. Відділ Магноліофіти, Квіткові або Покритонасінні Общая физиология сенсорных систем
Общая физиология сенсорных систем Отряд голенастые
Отряд голенастые Белгілерді ң тұқым қуалауының негізгі зандары, т ұқым қуалайтын белгілерді ңберілу ережелері
Белгілерді ң тұқым қуалауының негізгі зандары, т ұқым қуалайтын белгілерді ңберілу ережелері Правила техники безопасности в кабинете биологии
Правила техники безопасности в кабинете биологии Анатомия нервной системы
Анатомия нервной системы Тип членистоногие. Класс насекомые
Тип членистоногие. Класс насекомые Тип Членистоногие
Тип Членистоногие Листериоз. Род Listeria
Листериоз. Род Listeria Интегрированный урок по биологии,химии и физике. Викторина Колосок
Интегрированный урок по биологии,химии и физике. Викторина Колосок Дыхательная система. Органы дыхания
Дыхательная система. Органы дыхания Поведение собак
Поведение собак Основы селекции. Работы Н.И. Вавилова
Основы селекции. Работы Н.И. Вавилова Краткая история изучения клетки
Краткая история изучения клетки Обмен веществ. Возможные превращения веществ
Обмен веществ. Возможные превращения веществ Тканевые элементы нервной системы
Тканевые элементы нервной системы Какие функции выполняет корень?
Какие функции выполняет корень? Лисиця. Представниця класу ссавці, ряду хижі звірі
Лисиця. Представниця класу ссавці, ряду хижі звірі Анатомия крысы
Анатомия крысы Зимующие птицы
Зимующие птицы ГМО в пищевой промышленности
ГМО в пищевой промышленности Зоология - наука о животных. Подготовка к ОГЭ и ЕГЭ по биологии
Зоология - наука о животных. Подготовка к ОГЭ и ЕГЭ по биологии Основные фосфолипиды и гликолипиды тканей человека
Основные фосфолипиды и гликолипиды тканей человека