- Өсімдіктердің өнімділігіне әсер ететін процестері. (Лекция 9, 10)

Содержание
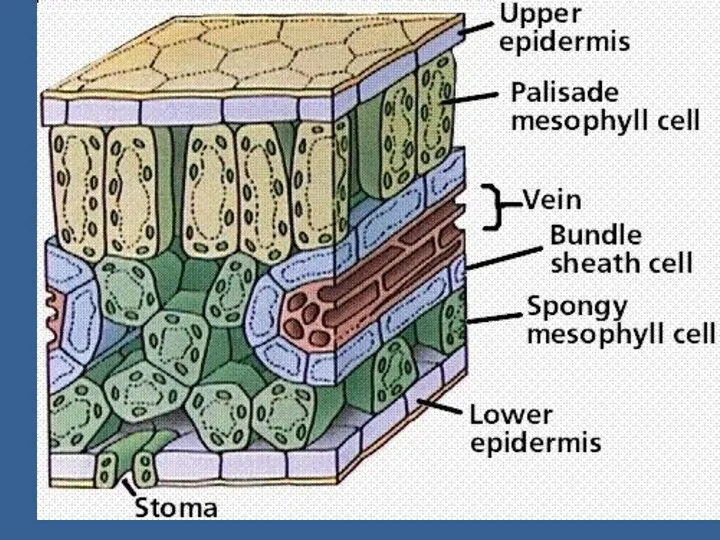
- 2. Өсімдіктердің өнімділікке әсер ететін процестер Фотосинтездің жарық сатысындағы процестердің өнімділікке маңызы Фотожүйе 1 және фотожүйе 2
- 7. Гендік деңгейінде ФС жоғарлағанда өсімдік экстенсивті жолды таңдайды – жапырақтың көлемі үлкейеді және хлорофилл мөлшерін жоғарылатады,
- 8. Қолайсыз жағдайда сахарозаның түзілуі төмендейді, АҚ мен орг. Қышқылдардың синтезі төмендейді. Жапыраққа енген CO2нің көміртегі тыныс
- 10. Өсімдікте масақ, жеміс пайда болғанда олар тамырмен ассимиляттар үшін бәсекелеседі. Бидайда масақ пайда болғанда оларға ассимиляттадың
- 11. Үлкен масақта 18 кішкене масақтың ішінде 3 жоғарғы масақтарды /колоска/ алып тастағанда ассимиляттардың 15% босап, олар
- 12. ФС өнімділігі өсімдіктің, жапырақтың бетінің функциясынмен байланысты, ал өнімділік өсімдіктің , массасының үлкеюімен байланысты. Біріншіге СО2нің
- 13. Фотосинтездеуші және ассимиляттарды қолданатын мүшелер дің донорлық-акцепторлық қарым қатынасы ФСдің белсенділігін ассимиляттарды қолданатын мүшелер бақылайды. ФСдік
- 15. ФС энергиялы процесс болғандықтан ол орындаушы роль атқарады. Сондықтан ФСА тын жақсартатын жұмыстар ассимиляттарды қолданатын мүшелердің
- 16. Эффективность продукции – отношение поглощенной энергии к единицу попавшей энергии Прежде Эффективность продукции - сух. Биомасса
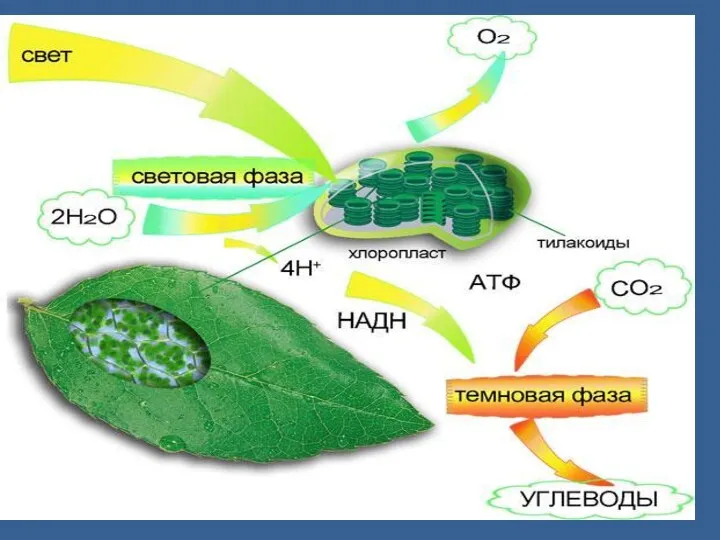
- 17. 1. Фотосинтездiк жуйелердегi энергия ауысу процестерi Фотосинтездiң жарық сатысы фотосинтездiк жүйелер құрамындары пигменттердiң жарық квантын (фотон)
- 20. 1 және 1. 2 фотожүйелер. Циклдік және циклсіз электрон тасмалдау жолдары. Фотосинтездің фотохимиялық реакциялары –күн сәулесінің
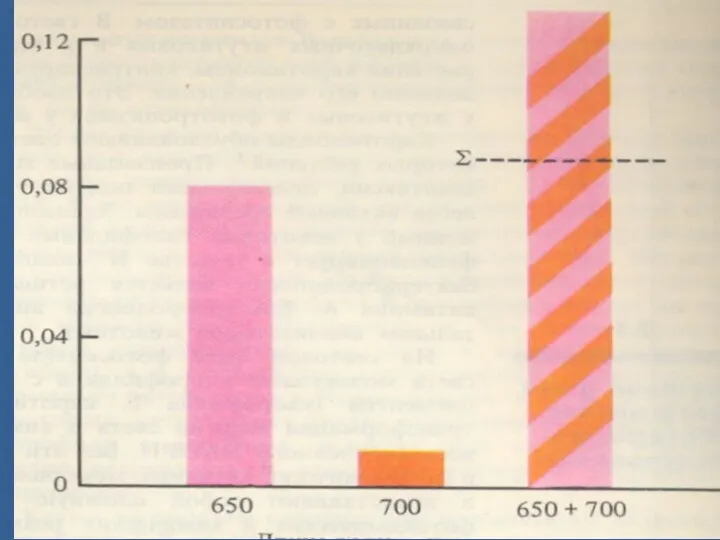
- 21. Эмерсон эффектісі. 680—700 нм ұзындығы бар толқындардың тиімділігін ұлғайту үшін ұзыныдығы қысқа (650—660 им) толқындарды қосу
- 25. Сіңірілген сәуле кванттардың энергиясы бірнеше жүз пигменттерден фотохимиялық реакциялық орталыққа жиналады Онда белгілі хлорофилл а молекуласының
- 26. Сіңірілген сәуле кванттар энергиясы Сәуле жинайтын кешен /бірнеше жүз пигменттер/ Фотохимиялық реакциялық орталығы хлорофилл а молекуласының
- 28. СЖК - сәулелерді жинайтын кешен. Хлорофилл а молекуласының жұпы - СЖК- пигменттерінен өткен қозу энергияны ұстайтын
- 30. Фотожүйе – фотосинтез аппаратының маңызды құрылымды-функциональды бөлімі. Ф/с процеске 2 фотожүйе қатысады
- 31. Фотожүйе Сәуле жинайтын комплекс Фотохимиялық реакциялардың орталығы Электрон тасмалдағыштар жиынтығы
- 34. ФЖ1 сәуленің әсерінен суда еритін FeS белок ферредоксинді (Фд) тотықсыздандырады және құрамында мыс бар, суда еритін
- 35. ФЖ 2 Реакциалық орталығы - хлорофилл Р 680 Антеналық пигменттер - хлорофиллдер А 670-68З Феофетин а
- 36. Суды тотықтыратын белок комплексі S, S пен байланысқан және П680-ге электрон доноры болып келетін электрон тасмалдағыш
- 37. ФЖ1 – НАДРН ФЖ2 – О2 Электрон тасмалдағыш тізбек екі фотожүйені байланыстырады - белок цитохромдық -
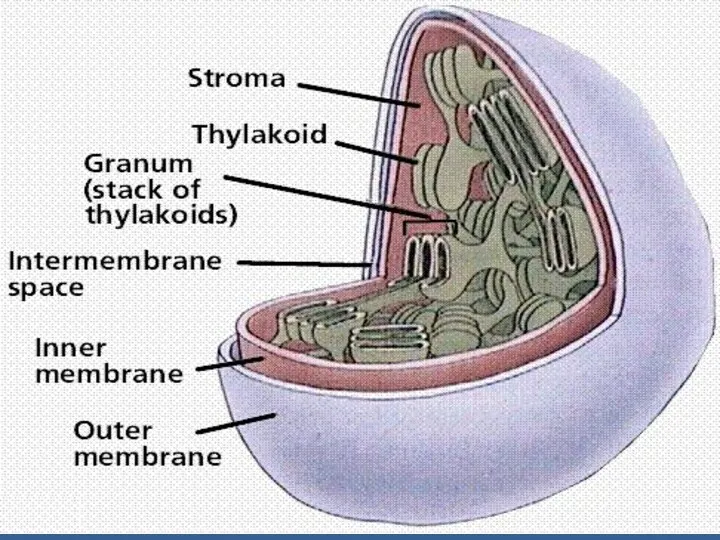
- 38. ФЖ 2-нің сәуле жинайтын комплексі және пигмент-белоктық комплексі тилакоидтардың жақын тұрған мембраналарында - грана тилакоидтарда -
- 39. Сәуле жинайтын кешен Пигмент-белоктық кешен ФЖ 2 Грана тилакоидтарында
- 40. Сәуле жинайтын кешен АТРазалық комплексі (CF 1 + CF о) ФЖ 1 Строма тилакоидтарында
- 41. Цитохромдар комплексі б - f строманың және граналардың тилакоидтарында орналасқан. Олар бір бірімен қалай қатынасады? Олар
- 42. Олар протондар мен электондарды тасмалдайды, немесе тек протондарды. Протон немесе электрон беретін молекула тотығады, алатын –
- 43. Айналымсыз тасымалдауда су фотототығады да электрондар судан НАДФ-қа тасмалданады. Тотығу-тотықсыздану реакцияларда бөлінетін энергия АТФтің синтезіне қолданылады.
- 44. Айналымсыз тасмалдануға екі ФЖ қатысады. П680-дегі электрондық тесік күшті тотықтырғыш болып істейді, ол бірнеше тасмалдағыш арқылы
- 45. Ол электрон П680 молекуласындағы тесікті бітейді. Су фотототығады да оттегінің молекуласы бөлінеді. 4 һν 2Н2О→4Н+ +
- 46. П680 Фф бірінші акцептор Пх /ол электрондарды және протондарды тасмалдайды/ б/f-комплекс / темір-күкірт белоктан Цх f-ке
- 47. ФЖ2- тотыққан П680 - күшті тотықтырғыш. Соның арқасында ФЖ2 су ыдырайды, оттегі бөлінеді ФЖ2-ге кіретін су
- 48. Су ыдырататын комплекстен П680-ге 4 электрон тасмалданғаннан кейін судың 2 молекуласы ыдырайды, оттегінің 1 молекуласы және
- 49. Айналымсыз фотофосфорланудың теңдігі : 2НАДФ + 2Н20 + 2АДФ + 2Н3Р04 → 2НАДФН + 2Н+ +
- 50. Айналымсыз фотофосфорланудың өнімі: 1. Тотықсызданған никотинамидадениндинрнуклеотидфосфат (НАДФН + Н) 2. АТФ. Олар ФС-дің қараңғы сатысында қолданылады.
- 51. Айналмалы фотофосфорлануға тек ФЖ1 қатысады. П700 бірінші тасмалдағыш Фд П700 Электронтасмалдағыш тізбекте цитохром б мен цитохром
- 52. Бұл жағдайда П700 донор да, акцептор да болып келеді. Айналмалы фотосинтездік фотофосфорланудың суммардық теңдігі: АДФ+Н3РО4+ һν
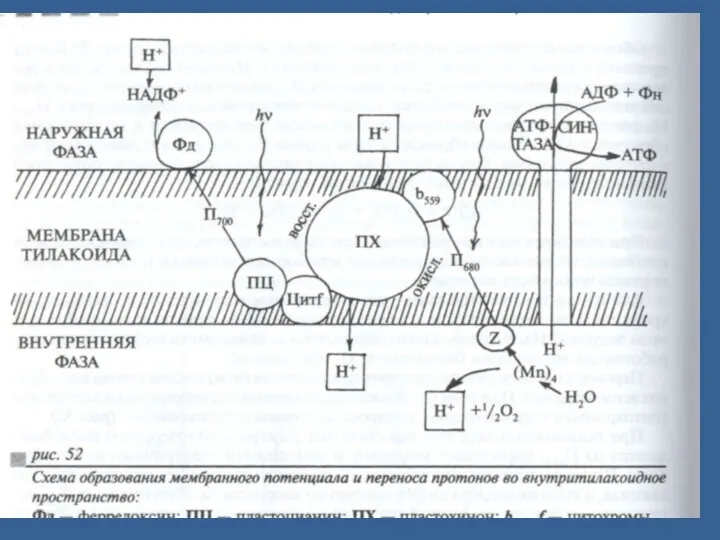
- 53. П.Митчеллдің хемиосмостық теориясы (ағылшын биохимик). Электрон тасмалдағыштар (электронтасмалдағыш тізбек) мембраналарда ассиметриялық орналасқан. Сонда тек электрондарды тасмалдайтын
- 58. Сәуленің 2 квантын сіңіргенде 2 эл-н (кезекпен) П680-нен шығып, мембранадан өтеді, орынына электрондар судан келеді. Онда
- 59. Электрондар П680-нен мембрананың сыртқы жағындағы акцепторға тасмалданады. Ол акцептордың аты - пластохинон (ПХ). Пластохинон — хинонның
- 60. Пх молекуласы басында 2 электрон қабылдайды: ПХ + 2е→ ПХ2-. ПХ2- протонға химиялық өте жақын -
- 61. Сонымен, Хл680-тен Хл.700-ге 2 электрон тасмалданғанда тилакоидтардың ішкі кеңістігінде протондар жиналады. Протондардың стромадан тилакоидтардың ішкі кеңістігіне
- 62. Протондар мембрананың бір жағында жиналған арқасында мембранада түзелген энергия 2 компоненттен тұрады - химиялық және электрлік.
- 63. Протондардың мембранадан диффузиясы АТФ-синтаза деген ферменттік комплекс арқылы жүреді /сопрягающий фактор/. Бұл комплекстің түрі саңырауқұлаққа ұқсайды,
- 64. Белок F1 – ол белоктық комплекс, 5 субъединицалардан тұрады: α, β, γ, δ, ε. Белоктық комплексті
- 66. Фосфорланудың механизмі туралы 2 гипотеза бар (тікелей және жанама). 1-ші гипотеза – фосфаттық топ және АДФ
- 67. Эффективность продукции – отношение поглощенной энергии к единицу попавшей энергии Прежде Эффективность продукции - сух. Биомасса
- 69. По этой формуле рост увеличивается с увеличением ежедневного PPF – PHOTOSYNTHETIC PHOTON FLUX и повышением СО2.
- 70. Эффект температуры значительно меньше с 15 до 250С Отличие от контролируемых условий в полевых условиях цель-
- 71. Проблема с оптимизацией эффектив. Продуктивности используя единственныое измерение урожая в конце жизненного цикла не говорит ничегго
- 73. Успех в оптимизации продуктивности в контролир. Условиях полагается на модели, предсказывающие ответные реакции растений на действие
- 74. Измерение в режиме реального времени эффективности продукции через поток СО2 Измерение ФС в течение светового периода
- 75. Измеряется раздельно потоки углерода в корнях и надзем. Органах Нетто фотосинтез /на квад. М горизонт. Площади/
- 79. Это полезно для интеграции мгновенных измерений, показ на рис. 4, развить Среднее значение СО2 газообмена Рез.
- 80. Измерение СО2 является недеструктивным методом измерения ежедневного роста. 90% сух. Биомассы берется из ФС. Было измерено
- 82. Измерение детерминантов роста Вдобавок к измерению эффективности продукции измерение газообмена обеспечивает данные для отдельного измерения детерминантов
- 83. Gross ФС опред-ся как уровень нетто ФС в течение светового периода + урвень дыхания в течение
- 84. Эффективность использования углерода. – отношение значения нетто ФС /чистая продукция ФС/ к валовому ФС, полагая ,
- 86. Скачать презентацию

Өсімдіктердің өнімділікке әсер ететін процестер
Фотосинтездің жарық сатысындағы процестердің өнімділікке маңызы
Фотожүйе
Өсімдіктердің өнімділікке әсер ететін процестер
Фотосинтездің жарық сатысындағы процестердің өнімділікке маңызы
Фотожүйе
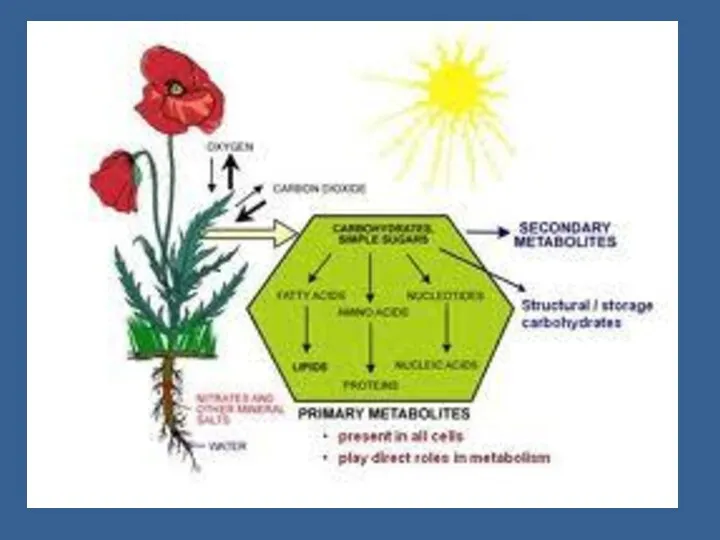

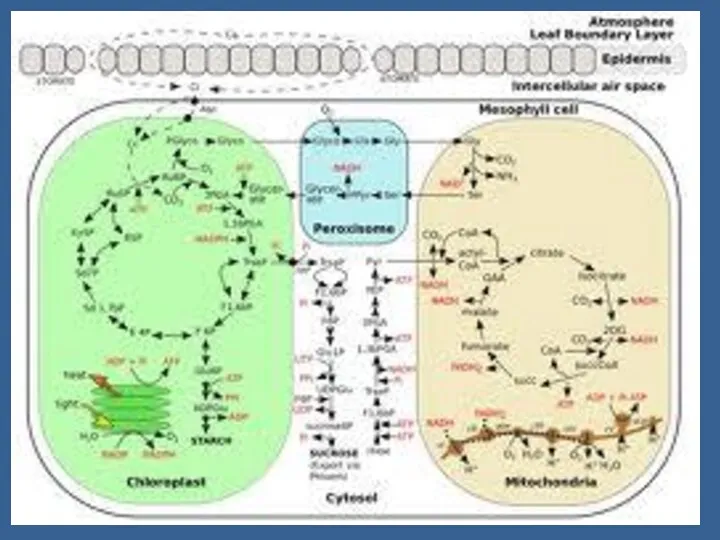


Гендік деңгейінде ФС жоғарлағанда өсімдік экстенсивті жолды таңдайды – жапырақтың көлемі
Гендік деңгейінде ФС жоғарлағанда өсімдік экстенсивті жолды таңдайды – жапырақтың көлемі

Қолайсыз жағдайда сахарозаның түзілуі төмендейді, АҚ мен орг. Қышқылдардың синтезі төмендейді.
Жапыраққа
Қолайсыз жағдайда сахарозаның түзілуі төмендейді, АҚ мен орг. Қышқылдардың синтезі төмендейді.
Жапыраққа
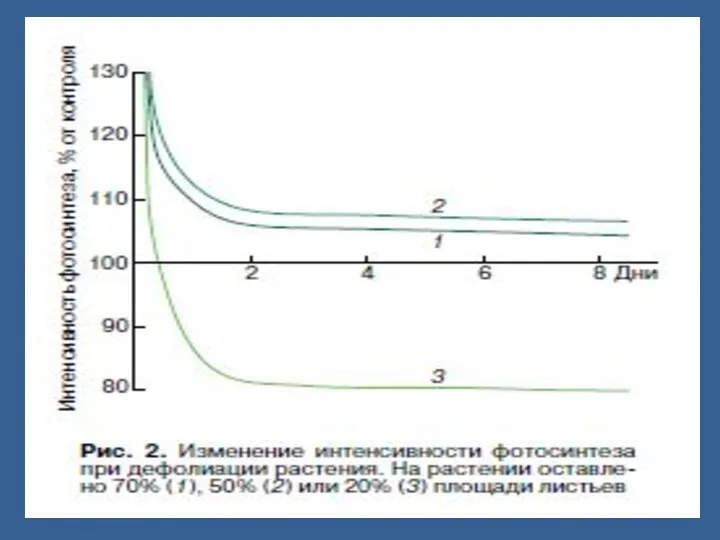

Өсімдікте масақ, жеміс пайда болғанда олар тамырмен ассимиляттар үшін бәсекелеседі.
Бидайда масақ
Өсімдікте масақ, жеміс пайда болғанда олар тамырмен ассимиляттар үшін бәсекелеседі.
Бидайда масақ

Үлкен масақта 18 кішкене масақтың ішінде 3 жоғарғы масақтарды /колоска/ алып
Үлкен масақта 18 кішкене масақтың ішінде 3 жоғарғы масақтарды /колоска/ алып

ФС өнімділігі өсімдіктің, жапырақтың бетінің функциясынмен байланысты, ал өнімділік өсімдіктің ,
ФС өнімділігі өсімдіктің, жапырақтың бетінің функциясынмен байланысты, ал өнімділік өсімдіктің ,

Фотосинтездеуші және ассимиляттарды қолданатын мүшелер дің донорлық-акцепторлық қарым қатынасы
ФСдің белсенділігін ассимиляттарды
Фотосинтездеуші және ассимиляттарды қолданатын мүшелер дің донорлық-акцепторлық қарым қатынасы
ФСдің белсенділігін ассимиляттарды
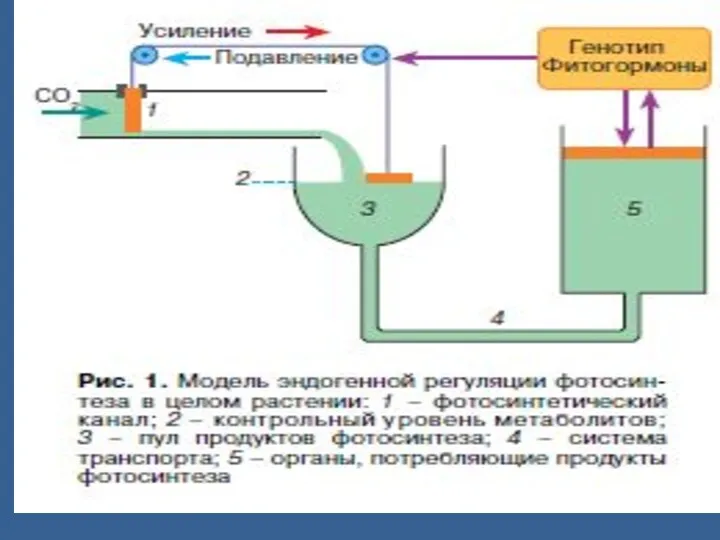

ФС энергиялы процесс болғандықтан ол орындаушы роль атқарады.
Сондықтан ФСА тын
ФС энергиялы процесс болғандықтан ол орындаушы роль атқарады.
Сондықтан ФСА тын

Эффективность продукции – отношение поглощенной энергии к единицу попавшей энергии
Прежде
Эффективность
Эффективность продукции – отношение поглощенной энергии к единицу попавшей энергии
Прежде
Эффективность

1. Фотосинтездiк жуйелердегi энергия ауысу процестерi
Фотосинтездiң жарық сатысы фотосинтездiк жүйелер құрамындары
1. Фотосинтездiк жуйелердегi энергия ауысу процестерi
Фотосинтездiң жарық сатысы фотосинтездiк жүйелер құрамындары

1 және 1. 2 фотожүйелер. Циклдік және циклсіз электрон тасмалдау жолдары.
1 және 1. 2 фотожүйелер. Циклдік және циклсіз электрон тасмалдау жолдары.

Эмерсон эффектісі.
680—700 нм ұзындығы бар толқындардың тиімділігін ұлғайту үшін ұзыныдығы қысқа
Эмерсон эффектісі.
680—700 нм ұзындығы бар толқындардың тиімділігін ұлғайту үшін ұзыныдығы қысқа
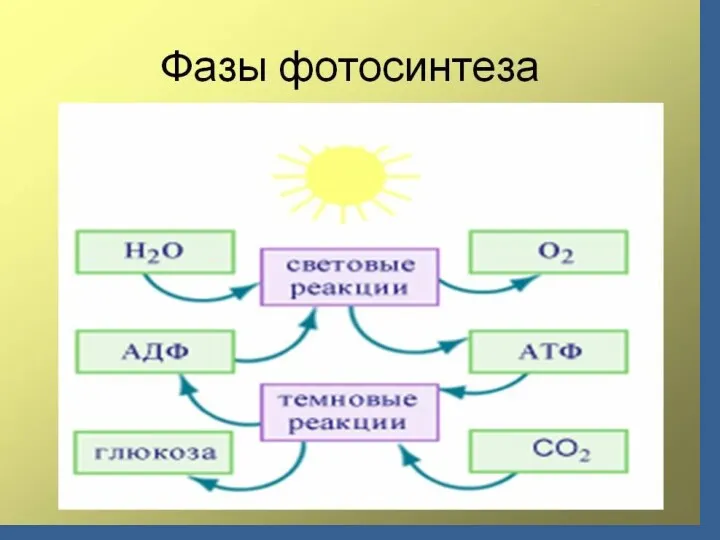

Сіңірілген сәуле кванттардың энергиясы бірнеше жүз пигменттерден фотохимиялық реакциялық орталыққа жиналады
Онда
Сіңірілген сәуле кванттардың энергиясы бірнеше жүз пигменттерден фотохимиялық реакциялық орталыққа жиналады
Онда

Сіңірілген сәуле кванттар
энергиясы
Сәуле жинайтын кешен
/бірнеше жүз пигменттер/
Фотохимиялық реакциялық
орталығы
хлорофилл а
Сіңірілген сәуле кванттар
энергиясы
Сәуле жинайтын кешен
/бірнеше жүз пигменттер/
Фотохимиялық реакциялық
орталығы
хлорофилл а
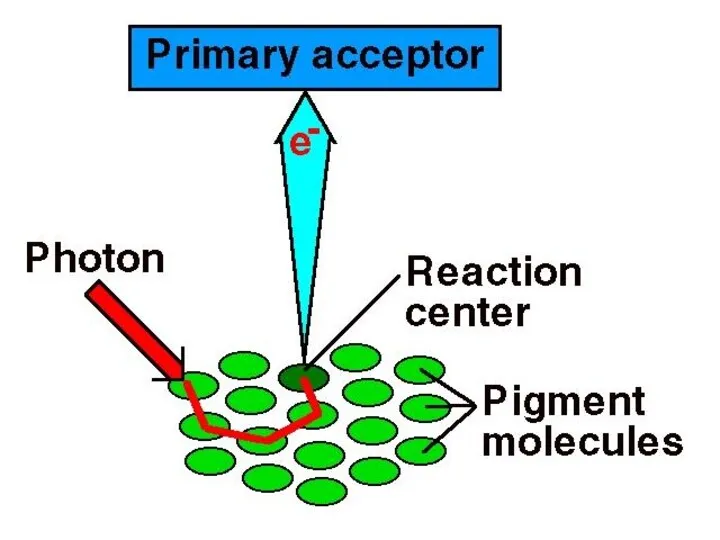

СЖК - сәулелерді жинайтын кешен.
Хлорофилл а молекуласының жұпы - СЖК-
СЖК - сәулелерді жинайтын кешен.
Хлорофилл а молекуласының жұпы - СЖК-
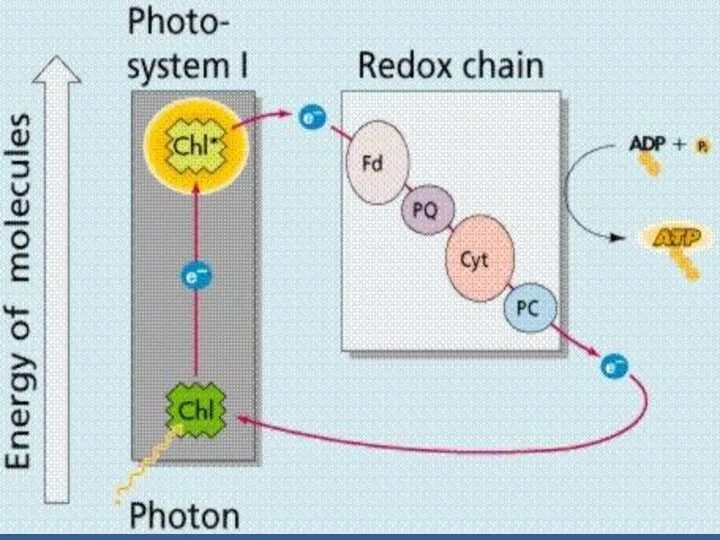

Фотожүйе – фотосинтез аппаратының маңызды құрылымды-функциональды бөлімі.
Ф/с процеске 2 фотожүйе
Ф/с процеске 2 фотожүйе
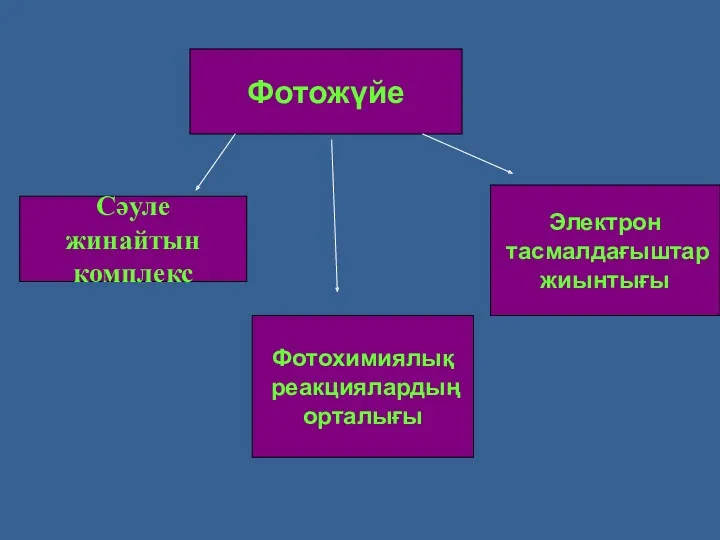
Фотожүйе
Сәуле жинайтын
комплекс
Фотохимиялық
реакциялардың
орталығы
Электрон
тасмалдағыштар
жиынтығы
Фотожүйе
Сәуле жинайтын
комплекс
Фотохимиялық
реакциялардың
орталығы
Электрон
тасмалдағыштар
жиынтығы



ФЖ1 сәуленің әсерінен суда еритін FeS белок ферредоксинді (Фд) тотықсыздандырады
ФЖ1 сәуленің әсерінен суда еритін FeS белок ферредоксинді (Фд) тотықсыздандырады

ФЖ 2
Реакциалық орталығы - хлорофилл Р 680
Антеналық пигменттер - хлорофиллдер
ФЖ 2
Реакциалық орталығы - хлорофилл Р 680
Антеналық пигменттер - хлорофиллдер

Суды тотықтыратын белок комплексі S,
S пен байланысқан және
Суды тотықтыратын белок комплексі S,
S пен байланысқан және

ФЖ1 – НАДРН
ФЖ2 – О2
Электрон тасмалдағыш тізбек екі фотожүйені байланыстырады -
ФЖ1 – НАДРН
ФЖ2 – О2
Электрон тасмалдағыш тізбек екі фотожүйені байланыстырады -

ФЖ 2-нің сәуле жинайтын комплексі және пигмент-белоктық комплексі тилакоидтардың жақын тұрған
ФЖ 2-нің сәуле жинайтын комплексі және пигмент-белоктық комплексі тилакоидтардың жақын тұрған
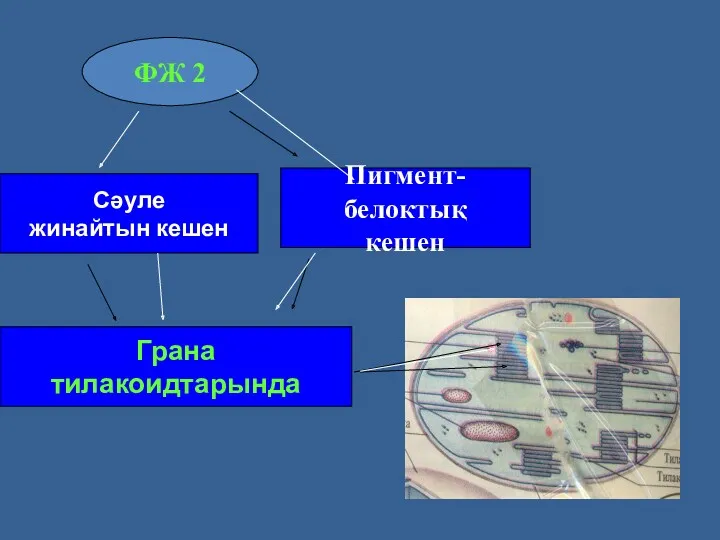
Сәуле
жинайтын кешен
Пигмент-белоктық
кешен
ФЖ 2
Грана
тилакоидтарында
Сәуле
жинайтын кешен
Пигмент-белоктық
кешен
ФЖ 2
Грана
тилакоидтарында
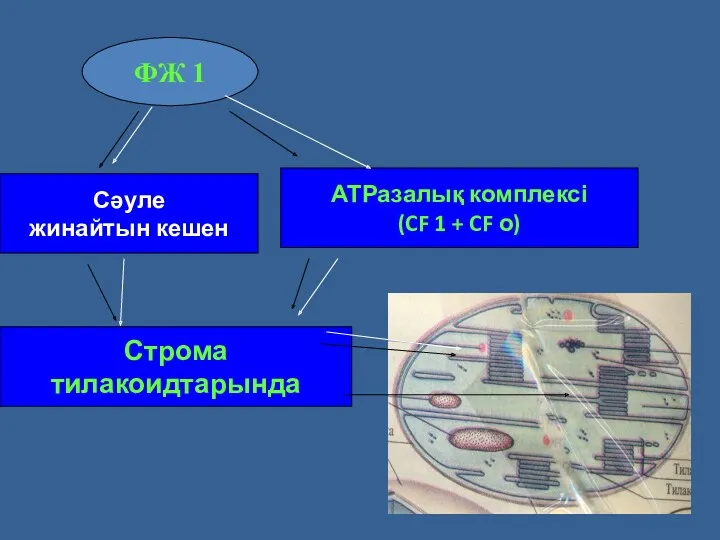
Сәуле
жинайтын кешен
АТРазалық комплексі
(CF 1 + CF о)
ФЖ 1
Строма
тилакоидтарында
Сәуле
жинайтын кешен
АТРазалық комплексі
(CF 1 + CF о)
ФЖ 1
Строма
тилакоидтарында

Цитохромдар комплексі б - f строманың және граналардың тилакоидтарында орналасқан.
Цитохромдар комплексі б - f строманың және граналардың тилакоидтарында орналасқан.

Олар протондар мен электондарды тасмалдайды, немесе тек протондарды.
Протон немесе электрон
Олар протондар мен электондарды тасмалдайды, немесе тек протондарды.
Протон немесе электрон

Айналымсыз тасымалдауда су фотототығады да электрондар судан НАДФ-қа тасмалданады.
Тотығу-тотықсыздану реакцияларда
Айналымсыз тасымалдауда су фотототығады да электрондар судан НАДФ-қа тасмалданады.
Тотығу-тотықсыздану реакцияларда

Айналымсыз тасмалдануға екі ФЖ қатысады.
П680-дегі электрондық тесік күшті тотықтырғыш болып
Айналымсыз тасмалдануға екі ФЖ қатысады.
П680-дегі электрондық тесік күшті тотықтырғыш болып

Ол электрон П680 молекуласындағы тесікті бітейді. Су фотототығады да оттегінің молекуласы
Ол электрон П680 молекуласындағы тесікті бітейді. Су фотототығады да оттегінің молекуласы

П680 Фф бірінші акцептор Пх /ол электрондарды және протондарды тасмалдайды/ б/f-комплекс
П680 Фф бірінші акцептор Пх /ол электрондарды және протондарды тасмалдайды/ б/f-комплекс

ФЖ2- тотыққан П680 - күшті тотықтырғыш.
Соның арқасында ФЖ2 су ыдырайды,
ФЖ2- тотыққан П680 - күшті тотықтырғыш.
Соның арқасында ФЖ2 су ыдырайды,

Су ыдырататын комплекстен П680-ге 4 электрон тасмалданғаннан кейін судың 2 молекуласы
Су ыдырататын комплекстен П680-ге 4 электрон тасмалданғаннан кейін судың 2 молекуласы

Айналымсыз фотофосфорланудың теңдігі :
2НАДФ + 2Н20 + 2АДФ + 2Н3Р04 →
Айналымсыз фотофосфорланудың теңдігі :
2НАДФ + 2Н20 + 2АДФ + 2Н3Р04 →

Айналымсыз фотофосфорланудың өнімі:
1. Тотықсызданған никотинамидадениндинрнуклеотидфосфат (НАДФН + Н)
2. АТФ.
Олар
Айналымсыз фотофосфорланудың өнімі:
1. Тотықсызданған никотинамидадениндинрнуклеотидфосфат (НАДФН + Н)
2. АТФ.
Олар

Айналмалы фотофосфорлануға тек ФЖ1 қатысады.
П700 бірінші тасмалдағыш Фд П700
Электронтасмалдағыш
Айналмалы фотофосфорлануға тек ФЖ1 қатысады.
П700 бірінші тасмалдағыш Фд П700
Электронтасмалдағыш

Бұл жағдайда П700 донор да, акцептор да болып келеді.
Айналмалы фотосинтездік
Бұл жағдайда П700 донор да, акцептор да болып келеді.
Айналмалы фотосинтездік

П.Митчеллдің хемиосмостық теориясы (ағылшын биохимик).
Электрон тасмалдағыштар (электронтасмалдағыш тізбек) мембраналарда ассиметриялық
П.Митчеллдің хемиосмостық теориясы (ағылшын биохимик).
Электрон тасмалдағыштар (электронтасмалдағыш тізбек) мембраналарда ассиметриялық
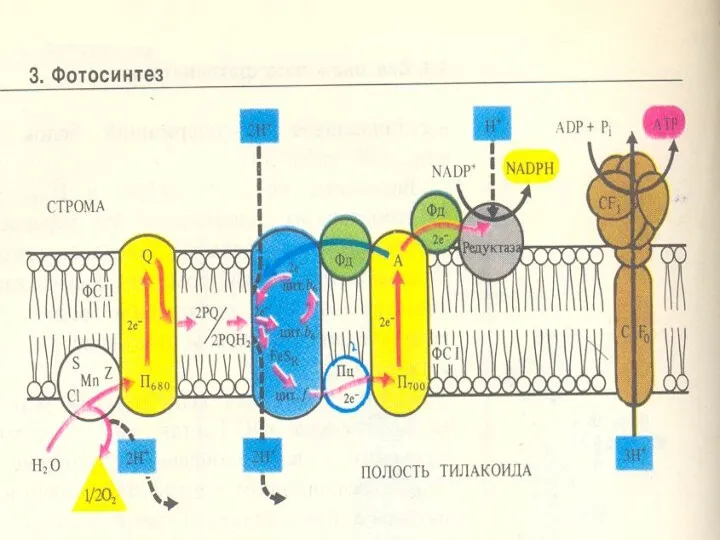
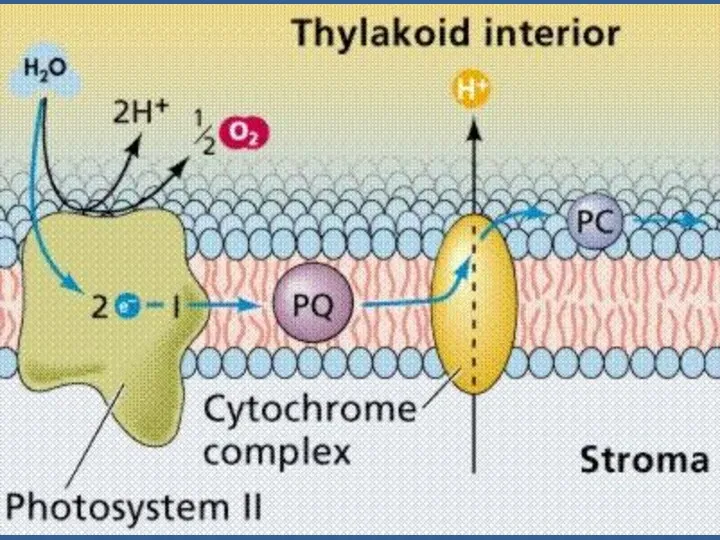
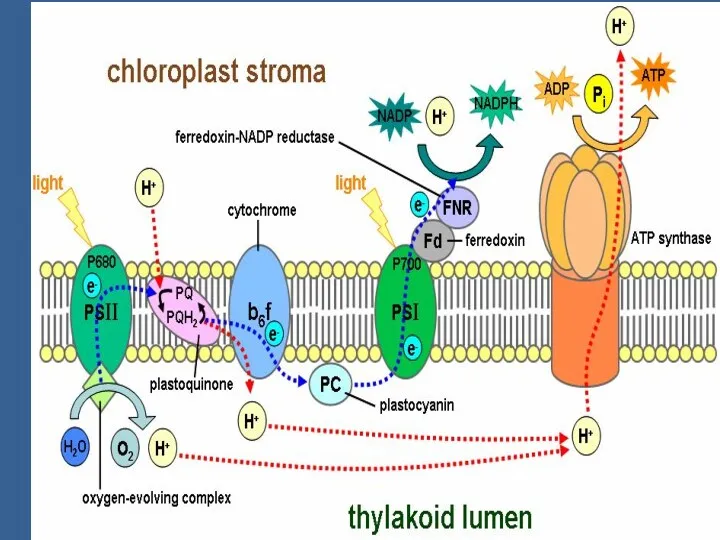

Сәуленің 2 квантын сіңіргенде 2 эл-н (кезекпен) П680-нен шығып, мембранадан өтеді,
Сәуленің 2 квантын сіңіргенде 2 эл-н (кезекпен) П680-нен шығып, мембранадан өтеді,

Электрондар П680-нен мембрананың сыртқы жағындағы акцепторға тасмалданады. Ол акцептордың аты -
Электрондар П680-нен мембрананың сыртқы жағындағы акцепторға тасмалданады. Ол акцептордың аты -

Пх молекуласы басында 2 электрон қабылдайды: ПХ + 2е→ ПХ2-.
ПХ2-
Пх молекуласы басында 2 электрон қабылдайды: ПХ + 2е→ ПХ2-.
ПХ2-

Сонымен, Хл680-тен Хл.700-ге 2 электрон тасмалданғанда тилакоидтардың ішкі кеңістігінде протондар жиналады.
Сонымен, Хл680-тен Хл.700-ге 2 электрон тасмалданғанда тилакоидтардың ішкі кеңістігінде протондар жиналады.

Протондар мембрананың бір жағында жиналған арқасында мембранада түзелген энергия 2 компоненттен
Протондар мембрананың бір жағында жиналған арқасында мембранада түзелген энергия 2 компоненттен

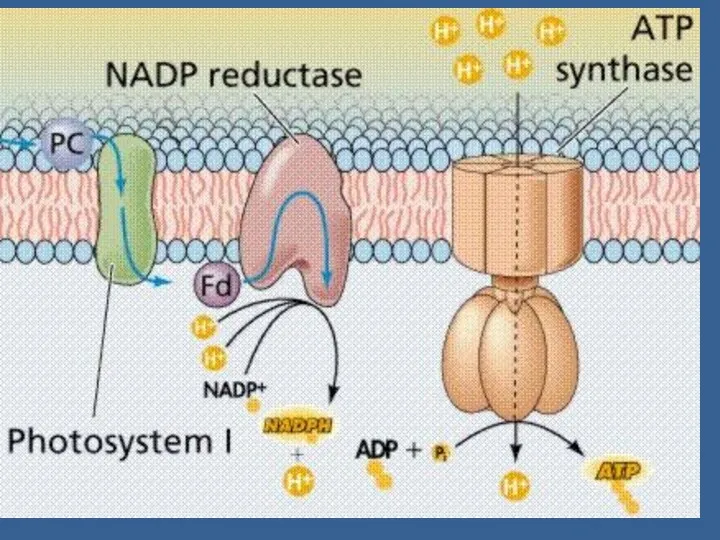
Протондардың мембранадан диффузиясы АТФ-синтаза деген ферменттік комплекс арқылы жүреді /сопрягающий фактор/.
Протондардың мембранадан диффузиясы АТФ-синтаза деген ферменттік комплекс арқылы жүреді /сопрягающий фактор/.

Белок F1 – ол белоктық комплекс, 5 субъединицалардан тұрады: α, β,
Белок F1 – ол белоктық комплекс, 5 субъединицалардан тұрады: α, β,

Фосфорланудың механизмі туралы 2 гипотеза бар (тікелей және жанама).
1-ші гипотеза
Фосфорланудың механизмі туралы 2 гипотеза бар (тікелей және жанама).
1-ші гипотеза

Эффективность продукции – отношение поглощенной энергии к единицу попавшей энергии
Прежде
Эффективность
Эффективность продукции – отношение поглощенной энергии к единицу попавшей энергии
Прежде
Эффективность
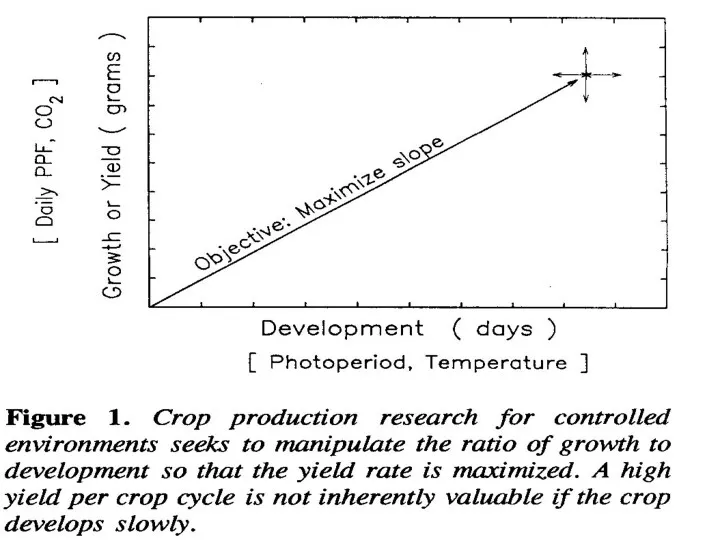

По этой формуле рост увеличивается с увеличением ежедневного PPF – PHOTOSYNTHETIC
По этой формуле рост увеличивается с увеличением ежедневного PPF – PHOTOSYNTHETIC

Эффект температуры значительно меньше с 15 до 250С
Отличие от контролируемых
Эффект температуры значительно меньше с 15 до 250С
Отличие от контролируемых

Проблема с оптимизацией эффектив. Продуктивности используя единственныое измерение урожая в конце
Проблема с оптимизацией эффектив. Продуктивности используя единственныое измерение урожая в конце
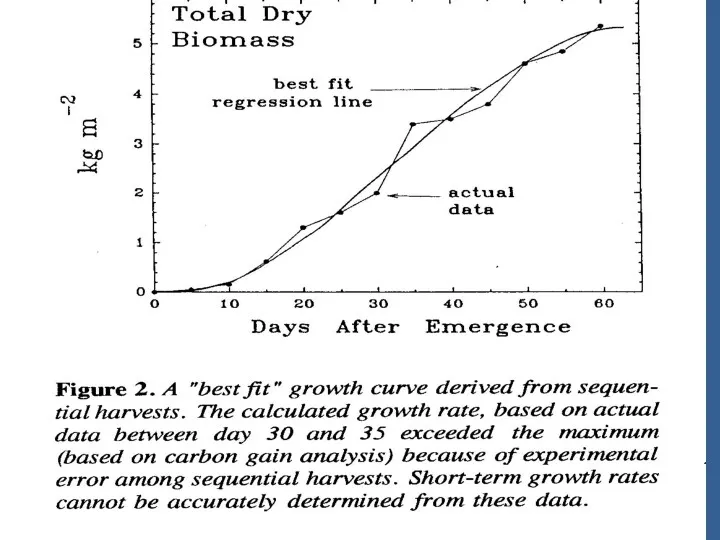

Успех в оптимизации продуктивности в контролир. Условиях полагается на модели, предсказывающие
Успех в оптимизации продуктивности в контролир. Условиях полагается на модели, предсказывающие

Измерение в режиме реального времени эффективности продукции через поток СО2
Измерение ФС
Измерение в режиме реального времени эффективности продукции через поток СО2
Измерение ФС

Измеряется раздельно потоки углерода в корнях и надзем. Органах
Нетто фотосинтез
Измеряется раздельно потоки углерода в корнях и надзем. Органах
Нетто фотосинтез
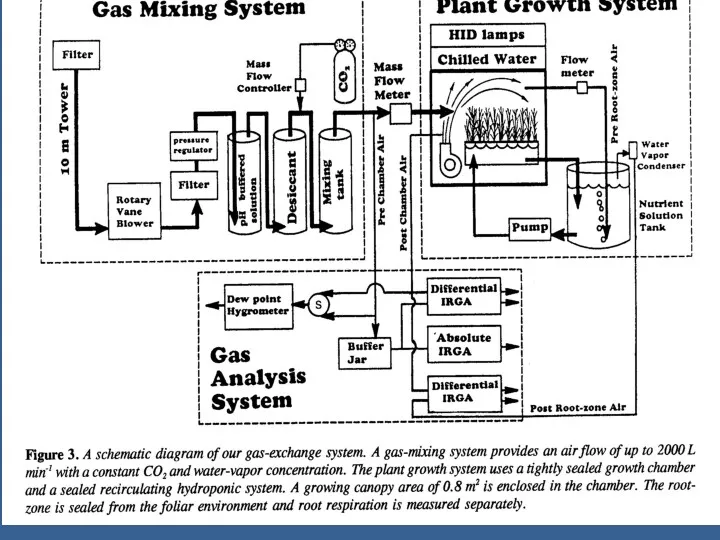
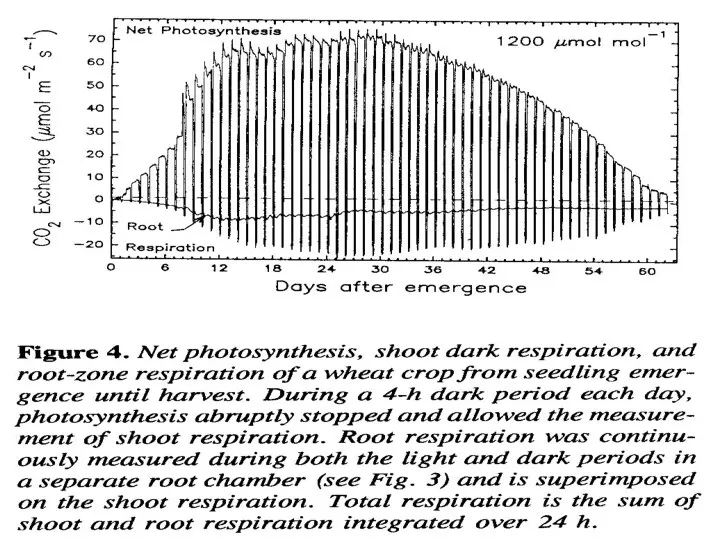
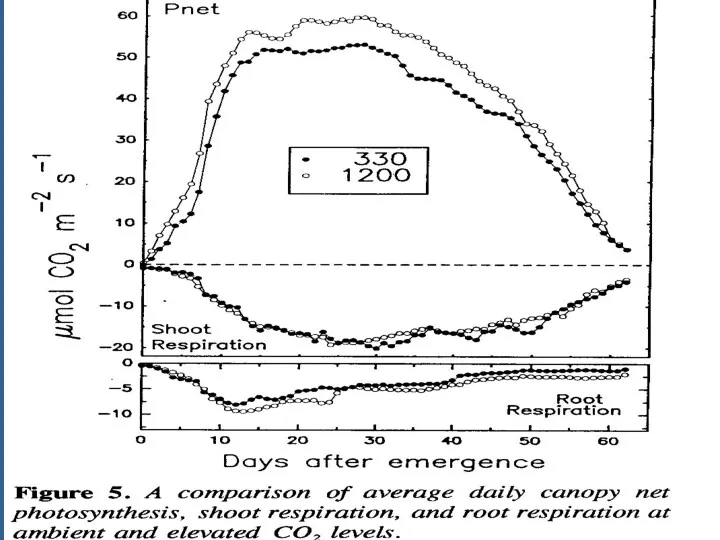

Это полезно для интеграции мгновенных измерений, показ на рис. 4, развить
Это полезно для интеграции мгновенных измерений, показ на рис. 4, развить

Измерение СО2 является недеструктивным методом измерения ежедневного роста.
90%
Измерение СО2 является недеструктивным методом измерения ежедневного роста.
90%
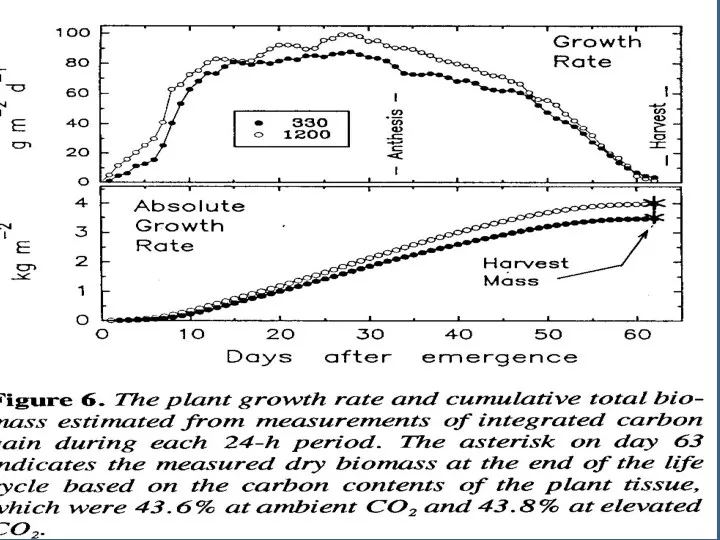

Измерение детерминантов роста
Вдобавок к измерению эффективности продукции измерение газообмена обеспечивает
Измерение детерминантов роста
Вдобавок к измерению эффективности продукции измерение газообмена обеспечивает

Gross ФС опред-ся как уровень нетто ФС в течение светового периода
Gross ФС опред-ся как уровень нетто ФС в течение светового периода

Эффективность использования углерода. – отношение значения нетто ФС /чистая продукция ФС/
Эффективность использования углерода. – отношение значения нетто ФС /чистая продукция ФС/
 Систематика растений. Многообразие растений
Систематика растений. Многообразие растений Пострансляционные модификации белков
Пострансляционные модификации белков Оплодотворение. Биологическое значение оплодотворения
Оплодотворение. Биологическое значение оплодотворения Основы цитологии. Неорганические вещества клетки
Основы цитологии. Неорганические вещества клетки Стационарная и динамическая организация функциональной межполушарной асимметрии. Введение в проблему
Стационарная и динамическая организация функциональной межполушарной асимметрии. Введение в проблему Биохимические основы скоростно-силовых качеств спортсмена
Биохимические основы скоростно-силовых качеств спортсмена Устройство речевого аппарата
Устройство речевого аппарата Ботаника – наука о растениях
Ботаника – наука о растениях презентация по теме МХИ биология 6 класс
презентация по теме МХИ биология 6 класс Процессы жизнеобеспечения в организме человека. Эндокринные железы (железы внутренней секреции)
Процессы жизнеобеспечения в организме человека. Эндокринные железы (железы внутренней секреции) Микроскопическое строение крови человека и лягушки. Лабораторная работа 3
Микроскопическое строение крови человека и лягушки. Лабораторная работа 3 Физиология возбудимых тканей (продолжение). Физиология нервов и нервных волокон. Физиология синапсов
Физиология возбудимых тканей (продолжение). Физиология нервов и нервных волокон. Физиология синапсов Орган зрения и зрительный анализатор
Орган зрения и зрительный анализатор Плод. Строение плода
Плод. Строение плода Лекция №4 Содержание и уход за собаками
Лекция №4 Содержание и уход за собаками Будова органів дихання
Будова органів дихання Движение крови и лимфы. Профилактика
Движение крови и лимфы. Профилактика Мини-презентация по теме Здоровое питание
Мини-презентация по теме Здоровое питание Экологическая ниша у разных видов растений и животных
Экологическая ниша у разных видов растений и животных Направления эволюции
Направления эволюции презентация Землетрясения
презентация Землетрясения Презентация к занятию по теме Аллергия
Презентация к занятию по теме Аллергия Органы цветковых растений. Корень
Органы цветковых растений. Корень Органы чувств. Регуляция деятельности
Органы чувств. Регуляция деятельности Растительность Челябинской области (часть 1)
Растительность Челябинской области (часть 1) Сенсорные системы. Анализаторы
Сенсорные системы. Анализаторы Птицы. Гнездование
Птицы. Гнездование Биологические ритмы человека
Биологические ритмы человека