- Өсімдіктерге жоғары температураның әсері

Содержание
- 2. Лекция 10 Өсімдіктерге сыртқы ортаның қолайсыз жағдайлардың әсері Қоршаған ортаның температурасының өзгеруі. Температура өзгерген жағдайда биополимерлердің
- 3. Эктотермалық организмдер, соның қатарында өсімдіктер, температураның өзгеруімен байланысты метаболизмнің өзгергуінің орынын толтыру үшін биополимерлерді модификациаландырады. Өзгерген
- 4. Ферменттік катализдің ырықтығының өзгеруі температура тәуелді реакциялардың жылдамдылығы өзгеруінің орынын толтыруға бағытталған. Реакциялардың жылдамдылығының өзгермеуі тек
- 5. 2. Ферменттердің қасиеттерінің өзгеруі Температура көбейген сайын ферменттік реациялардың жылдамдылығы ұлғайады. Реакциялардың жылдамдылығының температураға тәуелділігі Аррениус
- 6. Майысқақ құрылымдық ферменттерге конформациялық өзгерістер үшін аз Е қажет. Қатты құрылымдық ферменттерге конформациялық өзгерістер тән емес.
- 7. Бұдан басқа бейімділікке Михаэлистің Константасының (Кm) және субстраттың концентрациясының [S] қатынасының тұрақтығын сақтау жатады. Михаэлистің Константасы
- 8. Субстараттың Km физиологиялық концентрациясының диапазонының жартысына тең болып ұсталу керек. Бұл К мен [S]-тің қатынасында ферменттің
- 9. Т-ның өзгергуі Кm-нің өзгеруіне келтіреді. Михаэлистің константасы Т жоғарыланғанда көтеріледі. Бірақ өсімдіктерде және жануарлда ферменттердің Кm-і
- 10. Ферменттердің жаңа формалары акклимация процестерде стресс гендердің индукция нәтижесінде пайда болуы мүмкін. Ферменттердің түрлері изоферменттер деп
- 11. 3. Ферментердің мөлшерінің өзгеру арқылы Т-ның әсерінің орынын толтыру. Бұндай ферменттерге басты метаболизм процестерді шектейтін ферменттер
- 12. 4. Термофильдік бактериялар - термотұрақшылықты зерттеу механизмі. Термальдық бұлақтардағы- қайнарлардағы -Термофильдік бактериалар сыртқы Т- 100°С жақын
- 13. Термофилдерге тұрақтылықты протекторлық қосындылар да береді. Thermus thermophilus –тің белоктар синтезінің аппаратын ыдыраудан ПА сақтап қалады.
- 14. Сонымен жоғары температураға тұрақшылығын қамтамасыз ететін процестер: 1. Эл/статикалық байланыстардың көбейуі.. 2. Белоктардың байланыстардың көбейуі, алифатикалық
- 15. Жоғары Т-ға акклимациялануғы жылыту шоктың белоктарының маңызы. Өсімдіктер жылыту шокқа (ЖШ) ұшырағанда олардың ұлпаларымен мүшелері зақымданады,
- 16. Температураға тәуелді мембраналардың липидтік қосқабаттың модификациялары. Белоктың майсқақтығы мен тұрақтылығының арасында теңдік болса, Т-ға қарай фермент
- 17. Сұйық-кристалдық күйі липидтік бислойдың белгілі тығыздығы сәйкес. Қалыпты жағдайда липидтік бислойдың тығыздығы қатты сұйық күйден жоғары
- 18. Тасмалдаушы АТФазалар липидтердің тығыздығына өте сезімтал болады. Сонымен бислойдың сұйық-кристалдық күйінде сақтау - өте маңызды суыққа
- 20. Май қышқылдардың десатуразалары. МҚ-дың десатуразалары көміртек атомдар арасындағы жалғыз байланыстарды—СН2—СН2— қос байланыстарға айналдыруды катализдейді. ДСлар 3-валенттік
- 21. ДС-я көп күрделі биохимиялық реакциялар арқылы жүреді, ДСазалар тек соңғы сатысында қосылады. Осы реакциялардың маңызы -
- 22. ДСцияға қатысатын электрон тасмалдаушылар ЭНДР мен пластидтарада орналасады. ДСазалар бос және мембранамен байланысқан түрінде болады
- 23. Бос май қышқылдарды субстрат түрінде пайдаланатын ДСазалар мембраналардың липидтік құрамын реттеуге қатысады. Сонымен олар мембрананың сұйықтықтығын
- 24. Кейбір ДСалардың гендерінің экспрессиясы Т-ға тәуелді. Бұл құбылыс ДСазалардың акклимация, адаптация процестерге қатысады деген ұғарымға әкеледі.
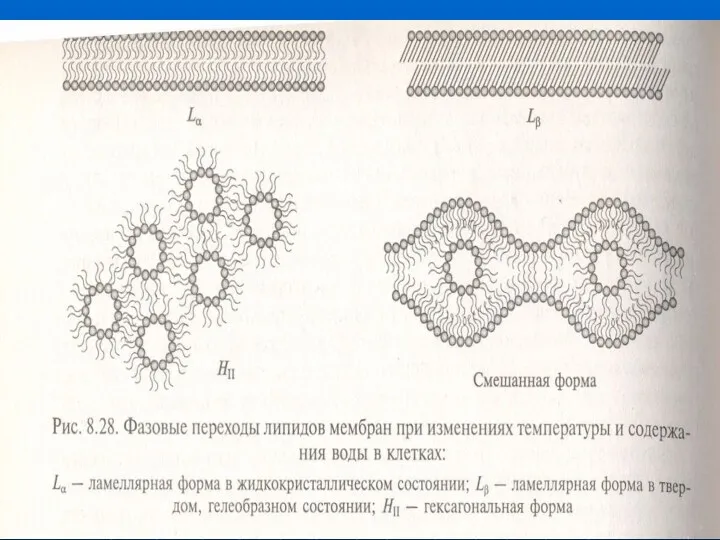
- 25. Липидтердің фазалық өтуінің Т-сы углеводородтық тізбегінің ұзындығына және қос байланысттарға тәуельді. Қос байланыстар айналмалар құрайды, олар
- 26. Т төмендегенде мембрана қатып қалмау үшін липидтердің май қышқылдарының көміртек атомы аз болып, қос байланыстар көп
- 27. Реакциялардың жылдамдылығы Т-мен қатар активацияның энергиясымен белгілінеді. Ферменттердің функциясы - активацияның энергиясын төмендету. Ферментің эффектитігі не
- 29. Скачать презентацию

Лекция 10
Өсімдіктерге сыртқы ортаның қолайсыз жағдайлардың әсері
Қоршаған ортаның температурасының өзгеруі.
Температура
Лекция 10
Өсімдіктерге сыртқы ортаның қолайсыз жағдайлардың әсері
Қоршаған ортаның температурасының өзгеруі.
Температура

Эктотермалық организмдер, соның қатарында өсімдіктер, температураның өзгеруімен байланысты метаболизмнің өзгергуінің орынын
Эктотермалық организмдер, соның қатарында өсімдіктер, температураның өзгеруімен байланысты метаболизмнің өзгергуінің орынын

Ферменттік катализдің ырықтығының өзгеруі температура тәуелді реакциялардың жылдамдылығы өзгеруінің орынын толтыруға
Ферменттік катализдің ырықтығының өзгеруі температура тәуелді реакциялардың жылдамдылығы өзгеруінің орынын толтыруға

2. Ферменттердің қасиеттерінің өзгеруі
Температура көбейген сайын ферменттік реациялардың жылдамдылығы ұлғайады. Реакциялардың
2. Ферменттердің қасиеттерінің өзгеруі
Температура көбейген сайын ферменттік реациялардың жылдамдылығы ұлғайады. Реакциялардың

Майысқақ құрылымдық ферменттерге конформациялық өзгерістер үшін аз Е қажет.
Қатты құрылымдық
Майысқақ құрылымдық ферменттерге конформациялық өзгерістер үшін аз Е қажет.
Қатты құрылымдық
![Бұдан басқа бейімділікке Михаэлистің Константасының (Кm) және субстраттың концентрациясының [S]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/203851/slide-6.jpg)
Бұдан басқа бейімділікке Михаэлистің Константасының (Кm) және субстраттың концентрациясының [S] қатынасының
Бұдан басқа бейімділікке Михаэлистің Константасының (Кm) және субстраттың концентрациясының [S] қатынасының

Субстараттың Km физиологиялық концентрациясының диапазонының жартысына тең болып ұсталу керек.
Бұл
Субстараттың Km физиологиялық концентрациясының диапазонының жартысына тең болып ұсталу керек.
Бұл

Т-ның өзгергуі Кm-нің өзгеруіне келтіреді.
Михаэлистің константасы Т жоғарыланғанда көтеріледі.
Бірақ
Т-ның өзгергуі Кm-нің өзгеруіне келтіреді.
Михаэлистің константасы Т жоғарыланғанда көтеріледі.
Бірақ

Ферменттердің жаңа формалары акклимация процестерде стресс гендердің индукция нәтижесінде пайда болуы
Ферменттердің жаңа формалары акклимация процестерде стресс гендердің индукция нәтижесінде пайда болуы

3. Ферментердің мөлшерінің өзгеру арқылы Т-ның әсерінің орынын толтыру.
Бұндай ферменттерге
3. Ферментердің мөлшерінің өзгеру арқылы Т-ның әсерінің орынын толтыру.
Бұндай ферменттерге

4. Термофильдік бактериялар - термотұрақшылықты зерттеу механизмі.
Термальдық бұлақтардағы- қайнарлардағы -Термофильдік
4. Термофильдік бактериялар - термотұрақшылықты зерттеу механизмі.
Термальдық бұлақтардағы- қайнарлардағы -Термофильдік

Термофилдерге тұрақтылықты протекторлық қосындылар да береді.
Thermus thermophilus –тің белоктар синтезінің
Термофилдерге тұрақтылықты протекторлық қосындылар да береді.
Thermus thermophilus –тің белоктар синтезінің

Сонымен жоғары температураға тұрақшылығын қамтамасыз ететін процестер:
1. Эл/статикалық байланыстардың көбейуі..
2. Белоктардың
Сонымен жоғары температураға тұрақшылығын қамтамасыз ететін процестер:
1. Эл/статикалық байланыстардың көбейуі..
2. Белоктардың

Жоғары Т-ға акклимациялануғы жылыту шоктың белоктарының маңызы.
Өсімдіктер жылыту шокқа (ЖШ)
Жоғары Т-ға акклимациялануғы жылыту шоктың белоктарының маңызы.
Өсімдіктер жылыту шокқа (ЖШ)

Температураға тәуелді мембраналардың липидтік қосқабаттың модификациялары.
Белоктың майсқақтығы мен тұрақтылығының арасында
Температураға тәуелді мембраналардың липидтік қосқабаттың модификациялары.
Белоктың майсқақтығы мен тұрақтылығының арасында

Сұйық-кристалдық күйі липидтік бислойдың белгілі тығыздығы сәйкес.
Қалыпты жағдайда липидтік бислойдың
Сұйық-кристалдық күйі липидтік бислойдың белгілі тығыздығы сәйкес.
Қалыпты жағдайда липидтік бислойдың

Тасмалдаушы АТФазалар липидтердің тығыздығына өте сезімтал болады. Сонымен бислойдың сұйық-кристалдық күйінде
Тасмалдаушы АТФазалар липидтердің тығыздығына өте сезімтал болады. Сонымен бислойдың сұйық-кристалдық күйінде

Май қышқылдардың десатуразалары.
МҚ-дың десатуразалары көміртек атомдар арасындағы жалғыз байланыстарды—СН2—СН2—
Май қышқылдардың десатуразалары.
МҚ-дың десатуразалары көміртек атомдар арасындағы жалғыз байланыстарды—СН2—СН2—

ДС-я көп күрделі биохимиялық реакциялар арқылы жүреді,
ДСазалар тек соңғы сатысында
ДС-я көп күрделі биохимиялық реакциялар арқылы жүреді,
ДСазалар тек соңғы сатысында
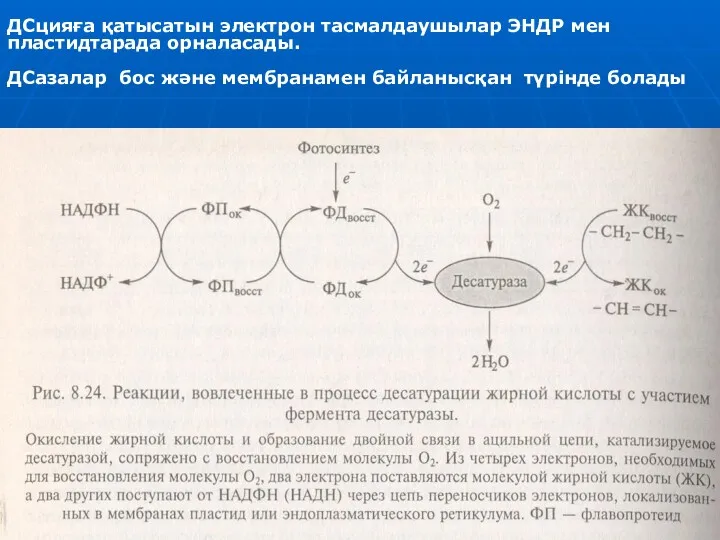
ДСцияға қатысатын электрон тасмалдаушылар ЭНДР мен пластидтарада орналасады.
ДСазалар бос және
ДСцияға қатысатын электрон тасмалдаушылар ЭНДР мен пластидтарада орналасады.
ДСазалар бос және

Бос май қышқылдарды субстрат түрінде пайдаланатын ДСазалар мембраналардың липидтік құрамын реттеуге
Бос май қышқылдарды субстрат түрінде пайдаланатын ДСазалар мембраналардың липидтік құрамын реттеуге

Кейбір ДСалардың гендерінің экспрессиясы Т-ға тәуелді.
Бұл құбылыс ДСазалардың акклимация, адаптация
Кейбір ДСалардың гендерінің экспрессиясы Т-ға тәуелді.
Бұл құбылыс ДСазалардың акклимация, адаптация

Липидтердің фазалық өтуінің Т-сы углеводородтық тізбегінің ұзындығына және қос байланысттарға тәуельді.
Липидтердің фазалық өтуінің Т-сы углеводородтық тізбегінің ұзындығына және қос байланысттарға тәуельді.

Т төмендегенде мембрана қатып қалмау үшін липидтердің май қышқылдарының көміртек атомы
Т төмендегенде мембрана қатып қалмау үшін липидтердің май қышқылдарының көміртек атомы

Реакциялардың жылдамдылығы Т-мен қатар активацияның энергиясымен белгілінеді. Ферменттердің функциясы - активацияның
Реакциялардың жылдамдылығы Т-мен қатар активацияның энергиясымен белгілінеді. Ферменттердің функциясы - активацияның
 История биологии и ее дисциплины
История биологии и ее дисциплины Анализаторы в системной организации поведения. Строение и функции анализаторов с позиции учения И.П. Павлова
Анализаторы в системной организации поведения. Строение и функции анализаторов с позиции учения И.П. Павлова Буынаяқтылар типі - arthropoda
Буынаяқтылар типі - arthropoda Органические вещества клетки. Белки
Органические вещества клетки. Белки Сад и его обитатели
Сад и его обитатели Шапинкові гриби
Шапинкові гриби Тема уроку: Інтенсивна технологія вирощування картоплі
Тема уроку: Інтенсивна технологія вирощування картоплі Морфология пищеварительной системы
Морфология пищеварительной системы Ткани растений
Ткани растений Професія дощового чер`яка
Професія дощового чер`яка Эволюция организмов
Эволюция организмов Направления эволюции
Направления эволюции Основные типы экологических взаимодействий
Основные типы экологических взаимодействий Строение организмов. Животные: морфология и анатомия. Хордовые
Строение организмов. Животные: морфология и анатомия. Хордовые Строение слизистой оболочки носа
Строение слизистой оболочки носа Презентация к уроку Строении функции головного мозга
Презентация к уроку Строении функции головного мозга The requirement of growing tomatoes in the greenhouse
The requirement of growing tomatoes in the greenhouse Жеміс дақылдарының бактериялы күйігіerwinia amilovora (burrill) winston etбактериальный ожог плодовых культур
Жеміс дақылдарының бактериялы күйігіerwinia amilovora (burrill) winston etбактериальный ожог плодовых культур Семейство паслёновые
Семейство паслёновые Процессы видообразование
Процессы видообразование Genome annotation
Genome annotation Экзотические домашние животные
Экзотические домашние животные Путешествие маленьких икринок
Путешествие маленьких икринок Екіншілік метаболиттер алу биотехнологиясы
Екіншілік метаболиттер алу биотехнологиясы Культура горох посевной
Культура горох посевной Мордовский государственный заповедник имени П.Г.Смидовича
Мордовский государственный заповедник имени П.Г.Смидовича Оценка динамики численности ресурсных видов Рассказовского и Пичаевского районов Тамбовской области
Оценка динамики численности ресурсных видов Рассказовского и Пичаевского районов Тамбовской области Мезозойская эра
Мезозойская эра