- Соединительная ткань. Биохимия межклеточного матрикса. (Тема 8)

Содержание
- 2. Межклеточный матрикс. Основные положения. Ткань, в которой межклеточный матрикс занимает более значительный объем , чем клетки
- 3. Основные компоненты межклеточного матрикса А. Структурные белки: коллаген, эластин; Б. Неколлагеновые адгезивные белки: фибронектин, остеонектин, ламинин,
- 4. Коллаген – основной структурный белок соед.ткани Коллаген - фибриллярный белок –гликопротеид- основной белок кожи, сухожилий, кровеносных
- 5. Коллаген. Строение Молекула коллагена (тропоколлаген) представляет спираль из 3 про-α-цепей, одинаковой длины. про-α-цепь – необычная полипептидная
- 6. Этапы синтеза созревания коллагена. Внутриклеточный этап Сложный, многоэтапный процесс начинается в клетке, заканчивается в матриксе. I.Внутриклеточный
- 7. Этапы синтеза созревания коллагена. Внутриклеточный этап 2. Гидроксилирование пролина и лизина в про- α-цепи (посттрансляционная реакция).
- 8. Особенности про-α-цепи Про-α-цепь имеет особенности, которые обеспечивают особенности пространственной структуры молекулы коллагена: Каждая третья аминок-та в
- 9. Этапы биосинтеза коллагена. Внутриклеточный этап ( продолжение) 3. Гликозилирование остатков аминокислот (аспарагиновой и оксилизина). Три про-α
- 10. Синтез коллагена. Внеклеточный этап. 1. Отщепляются концевые пептиды про-α-цепей в составе молекулы проколлагена. Образуются молекулы коллагена
- 11. Образование фибрилл в межклеточном матриксе Образование фибрилл из молекул коллагена Фибриллы представляют ступенчато расположенные ряды молекул
- 12. Коллаген. Обмен. Регуляция. Коллаген – медленно обменивающийся белок. Большинство протеолитических ферментов тканей и протеазы ЖКТ не
- 13. Эластин Эластин –гликопротеин - основной компонент многокомпонентной системы – эластичных волокон межклеточного вещества стенок кровеносных сосудов,
- 14. Эластин. Структура. Десмозины Десмозины (от гречес. связка) – структуры которые и обеспечивают волокну эластичные, резиноподобные свойства
- 15. Эластин. Образование связей между молекулами эластина (СН2)3 (СН2)3 N (СН2)3 лизин лиз лиз лиз В образовании
- 16. Катаболизм эластина. Нарушение структуры - причины и следствие Катаболизм эластина катализирует эластазы. Наиболее активная эластаза –
- 17. Протеогликаны. Глюкозаминогликаны (ГАГ). Состояние матрикса зависит от соотношения коллагена и протеогликанов. Протеогликаны - крупные молекулы, содержат
- 18. Глюкозаминогликаны. Протеогликаны. В настоящее время известна структура шести классов ГАГ: гиалуроновая кислота, два класса хондроитинсульфатов, дерматансульфаты,
- 19. Агрегаты протеогликанов. Гиалуроновая кислота В матриксе протеогликаны образуют агрегаты, занимающие большие объемы матрикса. Связующая молекула –
- 20. Функции протеогликанов и ГАГ Обеспечивают тургор тканей; в суставной жидкости стекловидном теле глаза - роль смазочного
- 21. Распад протеогликанов и ГАГ Распад – физиологический процесс обновления макромолекул. В распаде протеогликанов участвуют протеиназы и
- 22. Адгезивные белки – неколлагеновые гликопротеины Белки адгезии обеспечивают интеграцию («склеивание») клеток и компонентов матрикса. К ним
- 24. Скачать презентацию

Межклеточный матрикс. Основные положения.
Ткань, в которой межклеточный матрикс занимает более значительный
Межклеточный матрикс. Основные положения.
Ткань, в которой межклеточный матрикс занимает более значительный

Основные компоненты межклеточного матрикса
А. Структурные белки: коллаген, эластин;
Б. Неколлагеновые адгезивные белки:
Основные компоненты межклеточного матрикса
А. Структурные белки: коллаген, эластин;
Б. Неколлагеновые адгезивные белки:

Коллаген – основной структурный белок соед.ткани
Коллаген - фибриллярный белок –гликопротеид- основной
Коллаген – основной структурный белок соед.ткани
Коллаген - фибриллярный белок –гликопротеид- основной

Коллаген. Строение
Молекула коллагена (тропоколлаген) представляет спираль из 3 про-α-цепей, одинаковой
Коллаген. Строение
Молекула коллагена (тропоколлаген) представляет спираль из 3 про-α-цепей, одинаковой

Этапы синтеза созревания коллагена.
Внутриклеточный этап
Сложный, многоэтапный процесс начинается в клетке,
Этапы синтеза созревания коллагена.
Внутриклеточный этап
Сложный, многоэтапный процесс начинается в клетке,

Этапы синтеза созревания коллагена.
Внутриклеточный этап
2. Гидроксилирование пролина и лизина в
Этапы синтеза созревания коллагена.
Внутриклеточный этап
2. Гидроксилирование пролина и лизина в

Особенности про-α-цепи
Про-α-цепь имеет особенности, которые обеспечивают особенности пространственной структуры молекулы коллагена:
Каждая
Особенности про-α-цепи
Про-α-цепь имеет особенности, которые обеспечивают особенности пространственной структуры молекулы коллагена:
Каждая

Этапы биосинтеза коллагена.
Внутриклеточный этап ( продолжение)
3. Гликозилирование остатков аминокислот
(аспарагиновой
Этапы биосинтеза коллагена.
Внутриклеточный этап ( продолжение)
3. Гликозилирование остатков аминокислот
(аспарагиновой

Синтез коллагена. Внеклеточный этап.
1. Отщепляются концевые пептиды про-α-цепей в составе молекулы
Синтез коллагена. Внеклеточный этап.
1. Отщепляются концевые пептиды про-α-цепей в составе молекулы

Образование фибрилл в межклеточном матриксе
Образование фибрилл из молекул коллагена
Фибриллы представляют ступенчато
Образование фибрилл в межклеточном матриксе
Образование фибрилл из молекул коллагена
Фибриллы представляют ступенчато

Коллаген. Обмен. Регуляция.
Коллаген – медленно обменивающийся белок. Большинство протеолитических ферментов тканей
Коллаген. Обмен. Регуляция.
Коллаген – медленно обменивающийся белок. Большинство протеолитических ферментов тканей

Эластин
Эластин –гликопротеин - основной компонент многокомпонентной системы – эластичных волокон межклеточного
Эластин
Эластин –гликопротеин - основной компонент многокомпонентной системы – эластичных волокон межклеточного

Эластин. Структура. Десмозины
Десмозины (от гречес. связка) – структуры которые и обеспечивают
Эластин. Структура. Десмозины
Десмозины (от гречес. связка) – структуры которые и обеспечивают
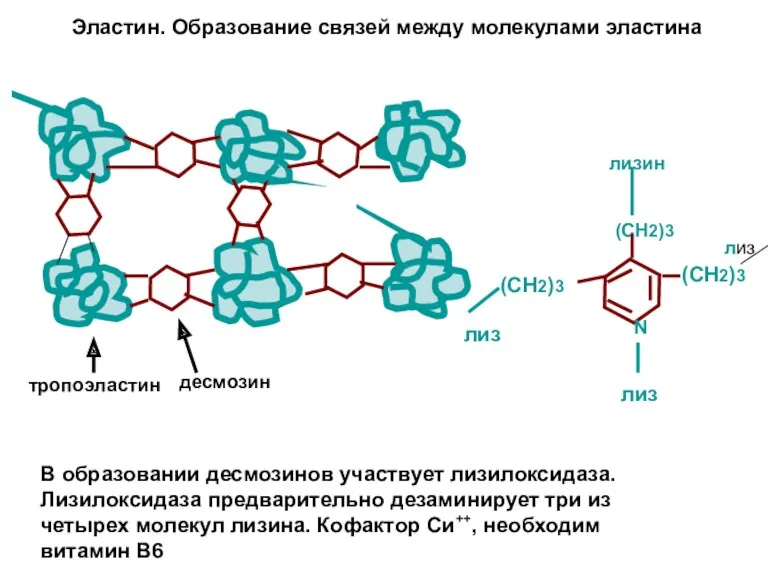
Эластин. Образование связей между молекулами эластина
(СН2)3
(СН2)3
N
(СН2)3
лизин
лиз
лиз
лиз
В образовании десмозинов участвует лизилоксидаза. Лизилоксидаза
Эластин. Образование связей между молекулами эластина
(СН2)3
(СН2)3
N
(СН2)3
лизин
лиз
лиз
лиз
В образовании десмозинов участвует лизилоксидаза. Лизилоксидаза

Катаболизм эластина. Нарушение структуры - причины и следствие
Катаболизм эластина катализирует эластазы.
Катаболизм эластина. Нарушение структуры - причины и следствие
Катаболизм эластина катализирует эластазы.

Протеогликаны. Глюкозаминогликаны (ГАГ).
Состояние матрикса зависит от соотношения коллагена и
Протеогликаны. Глюкозаминогликаны (ГАГ).
Состояние матрикса зависит от соотношения коллагена и

Глюкозаминогликаны. Протеогликаны.
В настоящее время известна структура шести классов ГАГ: гиалуроновая кислота,
Глюкозаминогликаны. Протеогликаны.
В настоящее время известна структура шести классов ГАГ: гиалуроновая кислота,

Агрегаты протеогликанов. Гиалуроновая кислота
В матриксе протеогликаны образуют агрегаты, занимающие большие объемы
Агрегаты протеогликанов. Гиалуроновая кислота
В матриксе протеогликаны образуют агрегаты, занимающие большие объемы

Функции протеогликанов и ГАГ
Обеспечивают тургор тканей; в суставной жидкости стекловидном теле
Функции протеогликанов и ГАГ
Обеспечивают тургор тканей; в суставной жидкости стекловидном теле

Распад протеогликанов и ГАГ
Распад – физиологический процесс обновления макромолекул.
В распаде
Распад протеогликанов и ГАГ
Распад – физиологический процесс обновления макромолекул.
В распаде

Адгезивные белки – неколлагеновые гликопротеины
Белки адгезии обеспечивают интеграцию («склеивание») клеток
Адгезивные белки – неколлагеновые гликопротеины
Белки адгезии обеспечивают интеграцию («склеивание») клеток
 Генетика пола. Наследование, сцепленное с полом
Генетика пола. Наследование, сцепленное с полом урок биологии в 7 классе Внешнее и внутреннее строение насекомых
урок биологии в 7 классе Внешнее и внутреннее строение насекомых Породы кошек
Породы кошек Происхождение человека. Расы и расогенез. Адаптивные типы людей
Происхождение человека. Расы и расогенез. Адаптивные типы людей Цветковые (покрытосемянные) растения
Цветковые (покрытосемянные) растения Многообразие организмов. Вирусы
Многообразие организмов. Вирусы Ядовитые грибы
Ядовитые грибы Птицы Ненецкого Автономного округа
Птицы Ненецкого Автономного округа Структура и динамика популяций. Внутривидовые и межвидовые взаимоотношения в популяциях, гомеостаз и экологические стратегии
Структура и динамика популяций. Внутривидовые и межвидовые взаимоотношения в популяциях, гомеостаз и экологические стратегии Психология и социобиология половых различий
Психология и социобиология половых различий Органы чувств. Строение и функции аппаратов глаза. Строение и гистофизиология органа обоняния
Органы чувств. Строение и функции аппаратов глаза. Строение и гистофизиология органа обоняния Факты о генетике
Факты о генетике Разнообразие зеленых водорослей
Разнообразие зеленых водорослей Животный мир Мещёры
Животный мир Мещёры Анатомия и морфология рыб
Анатомия и морфология рыб Обмен веществ и энергии
Обмен веществ и энергии Проблемы Антропогенеза
Проблемы Антропогенеза Мхи, хвощи, папоротники, плауны
Мхи, хвощи, папоротники, плауны Физиология гладких мышц. (Лекция 7)
Физиология гладких мышц. (Лекция 7) Морфология сельскохозяйственных животных. Аппарат движения. Остеология, миология
Морфология сельскохозяйственных животных. Аппарат движения. Остеология, миология Высшая нервная деятельность
Высшая нервная деятельность Презентация Лист. внешнее и внутреннее строение.
Презентация Лист. внешнее и внутреннее строение. Промежуточный мозг
Промежуточный мозг Углеводы. Классификация углеводов
Углеводы. Классификация углеводов Эмбриогенез печени и желчевыводящих путей
Эмбриогенез печени и желчевыводящих путей Транскрипция у эукариот
Транскрипция у эукариот Первая и вторая сигнальные системы человека
Первая и вторая сигнальные системы человека Презентация к внеклассному мероприятию Муравьи- общественные насекомые
Презентация к внеклассному мероприятию Муравьи- общественные насекомые