- Строение и скелетных и гладких мышц. физиологические свойства

Содержание
- 2. План лекции Лекция 2. Строение и физиологические свойства скелетных и гладких мышц. 1. Строение мышцы 2.
- 3. Типы мышц
- 4. Типы мышц
- 5. 1. Строение скелетной мышцы состоят из цилиндрических мышечных волокон (клеток), каждый конец мышцы соединен посредством сухожилий
- 6. Сарколеммой называют клеточную мембрану МВ Она состоит из плазматической мембраны, и наружного покрытия из тонкого слоя
- 7. (изотропная, светлая) (анизотропная, более темная) 2. Структура саркомера При изучении с помощью светового микроскопа, основной характеристикой
- 8. 3. Механизм мышечного сокращения В состоянии покоя молекулы тропомиозина располагаются поверх активных участков актиновых нитей, препятствуя
- 9. Теория скользящих нитей При укорочении волокна каждый поперечный мостик, прикрепившийся к тонкому (актиновому) филаменту, совершает поворот
- 10. В цикле поперечных мостиков АТФ выполняет две разные роли: 1) гидролиз АТФ поставляет энергию для движения
- 11. 5. Электромеханическое сопряжение Это последовательность процессов, в результате которых потенциал действия плазматической мембраны мышечного волокна приводит
- 12. Электромеханическое сопряжение ПД распространяется по мембране МВ и по мембране Т-трубочек вглубь клетки. Достигнув области Т-трубочки,
- 13. 6. Нейромоторная единица (двигательная) Нервные импульсы, идущие от мотонейрона, вызывают сокращения определенной группы мышечных волокон. Все
- 15. 7. ОМС, суммация, тетанус - регуляция силы сокращения частотой стимуляции одиночные сокращения суммация зубчатый тетанус (неполный)
- 16. 8. Гладкие мышцы
- 21. В унитарных гладких мышцах различают Спайковые ПД. Такие ПД возникают под действием электрической стимуляции, гормонов, медиаторов,
- 22. Вместо тропонина ГМК содержат регуляторный белок – кальмодулин. 1. 4 иона Са2+ связываются с кальмодулином. 2.
- 23. Механизм удаления Са2+ в гладких мышцах
- 25. В гладкой мышце изменения уровней внутриклеточного Са2+ могут происходить как на фоне изменений мембранного потенциала, (электромеханическое
- 26. При фармако-механическом сопряжении химические факторы (например, нейромедиаторы) могут индуцировать сокращение гладкой мышцы без изменения мембранного потенциала.
- 28. агонист рецептор Гладкая мышца Комплекс G-белка Фосфолипаза С Потенциал- управляемый Са2+-канал Са2+ входит в цитоплазму через
- 31. Скачать презентацию

План лекции
Лекция 2. Строение и физиологические свойства скелетных и гладких мышц.
1.
План лекции
Лекция 2. Строение и физиологические свойства скелетных и гладких мышц.
1.
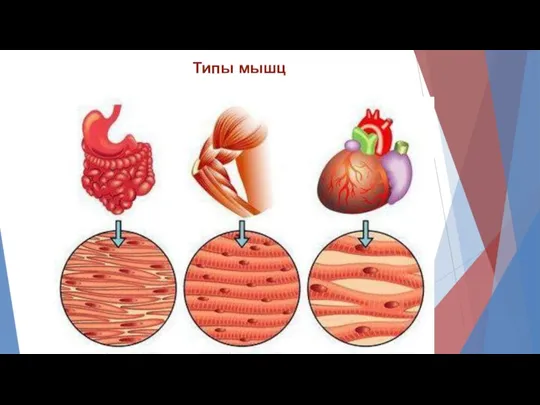
Типы мышц
Типы мышц
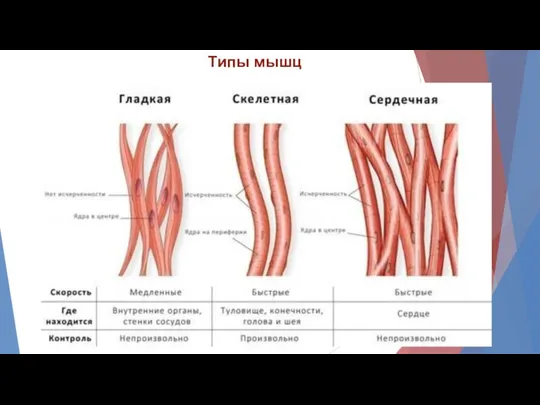
Типы мышц
Типы мышц
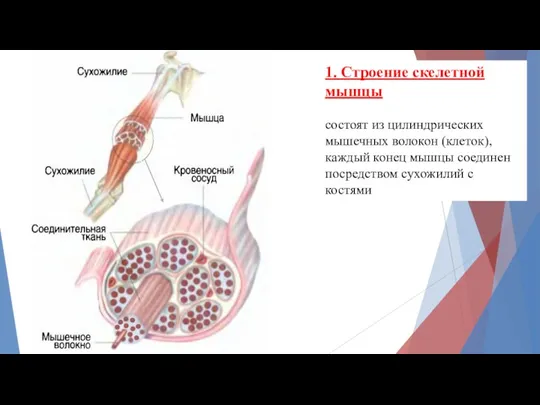
1. Строение скелетной мышцы
состоят из цилиндрических мышечных волокон (клеток),
каждый
1. Строение скелетной мышцы
состоят из цилиндрических мышечных волокон (клеток),
каждый
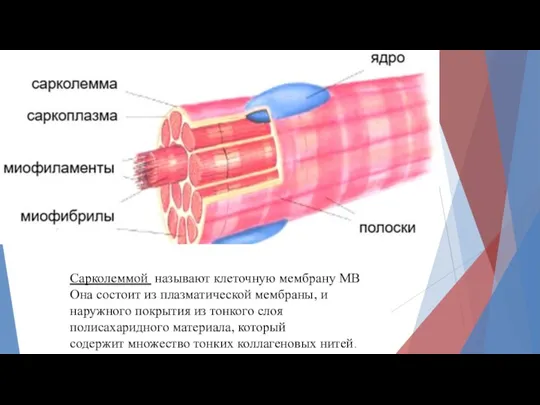
Сарколеммой называют клеточную мембрану МВ
Она состоит из плазматической мембраны, и
наружного покрытия
Сарколеммой называют клеточную мембрану МВ
Она состоит из плазматической мембраны, и
наружного покрытия

(изотропная, светлая)
(анизотропная, более темная)
2. Структура саркомера
При изучении с помощью светового микроскопа,
(изотропная, светлая)
(анизотропная, более темная)
2. Структура саркомера
При изучении с помощью светового микроскопа,
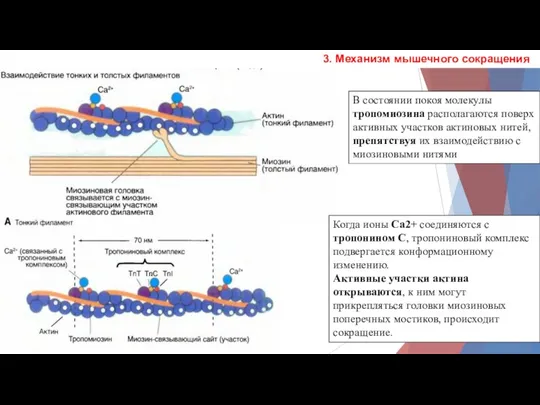
3. Механизм мышечного сокращения
В состоянии покоя молекулы тропомиозина располагаются поверх активных
3. Механизм мышечного сокращения
В состоянии покоя молекулы тропомиозина располагаются поверх активных
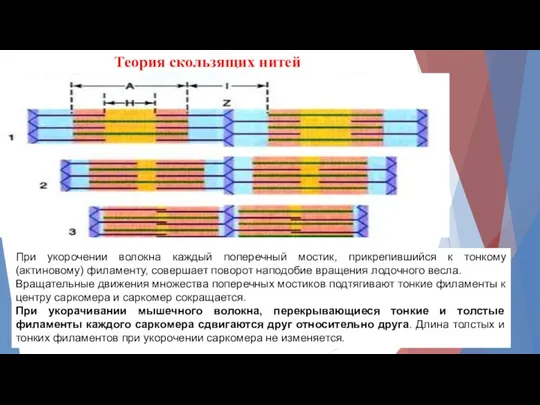
Теория скользящих нитей
При укорочении волокна каждый поперечный мостик, прикрепившийся к тонкому
Теория скользящих нитей
При укорочении волокна каждый поперечный мостик, прикрепившийся к тонкому
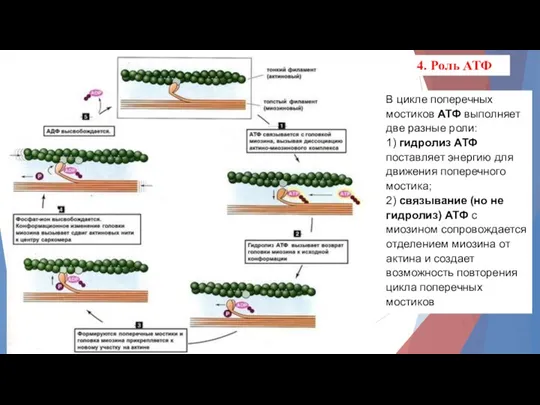
В цикле поперечных мостиков АТФ выполняет две разные роли:
1) гидролиз АТФ
В цикле поперечных мостиков АТФ выполняет две разные роли:
1) гидролиз АТФ
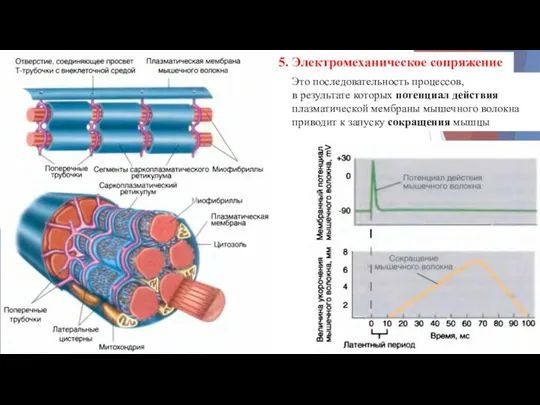
5. Электромеханическое сопряжение
Это последовательность процессов,
в результате которых потенциал действия
плазматической мембраны
5. Электромеханическое сопряжение
Это последовательность процессов,
в результате которых потенциал действия
плазматической мембраны
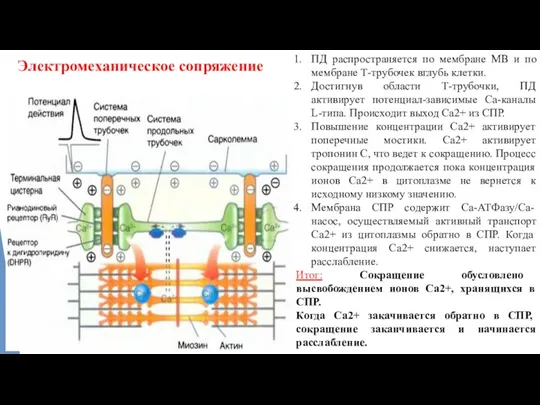
Электромеханическое сопряжение
ПД распространяется по мембране МВ и по мембране Т-трубочек вглубь
Электромеханическое сопряжение
ПД распространяется по мембране МВ и по мембране Т-трубочек вглубь
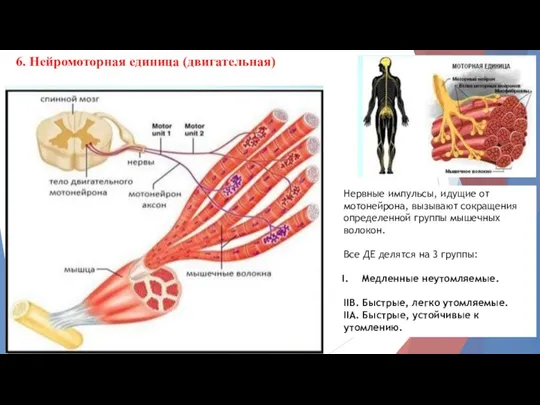
6. Нейромоторная единица (двигательная)
Нервные импульсы, идущие от мотонейрона, вызывают сокращения определенной
6. Нейромоторная единица (двигательная)
Нервные импульсы, идущие от мотонейрона, вызывают сокращения определенной
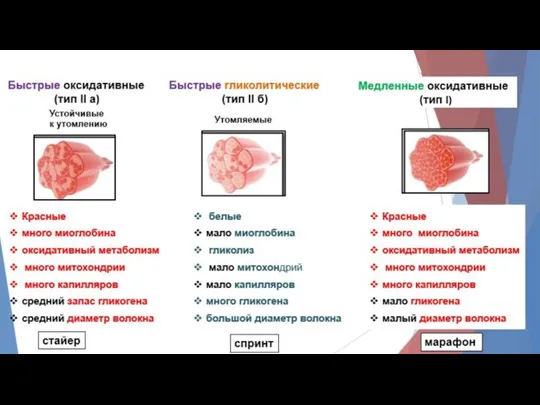
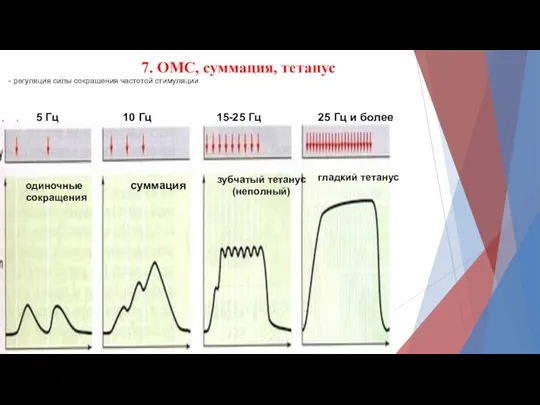
7. ОМС, суммация, тетанус
- регуляция силы сокращения частотой стимуляции
одиночные
сокращения
суммация
зубчатый
7. ОМС, суммация, тетанус
- регуляция силы сокращения частотой стимуляции
одиночные
сокращения
суммация
зубчатый
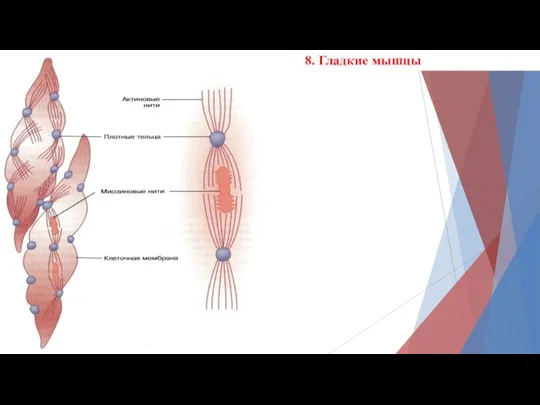
8. Гладкие мышцы
8. Гладкие мышцы


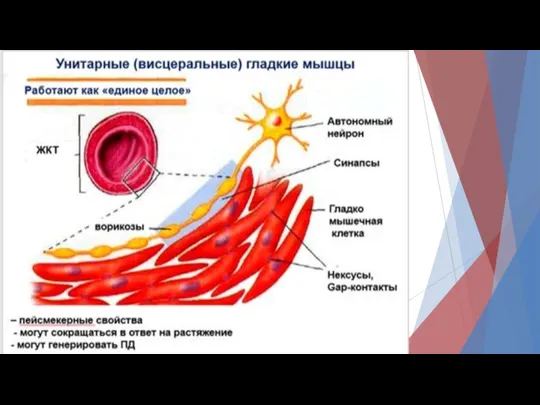
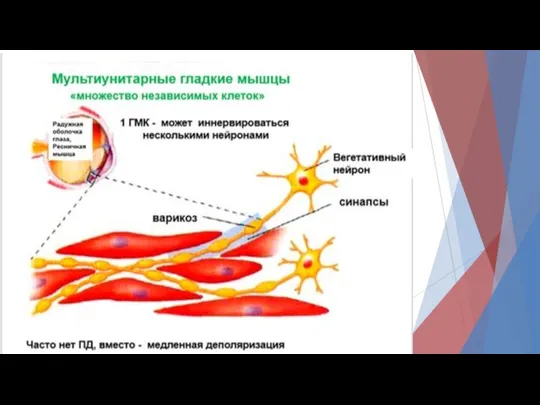
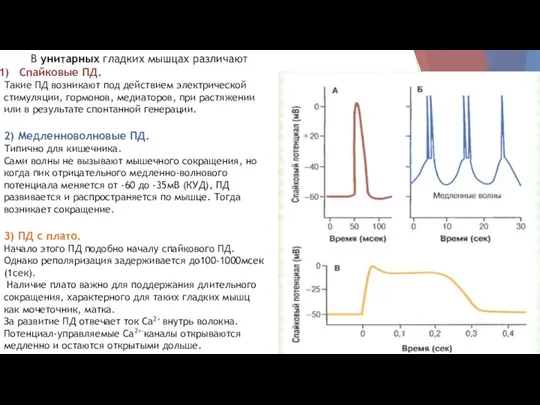
В унитарных гладких мышцах различают
Спайковые ПД.
Такие ПД возникают под
В унитарных гладких мышцах различают
Спайковые ПД.
Такие ПД возникают под

Вместо тропонина ГМК содержат регуляторный белок – кальмодулин.
1. 4 иона Са2+
Вместо тропонина ГМК содержат регуляторный белок – кальмодулин.
1. 4 иона Са2+
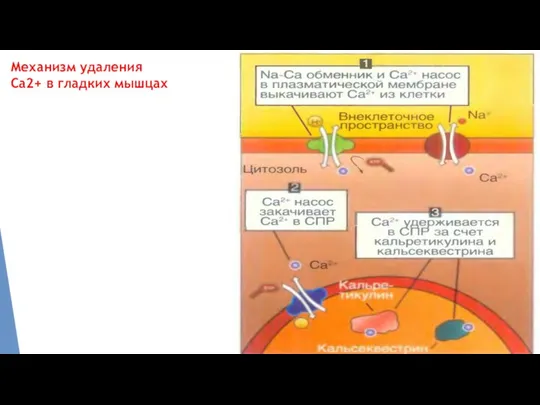
Механизм удаления Са2+ в гладких мышцах
Механизм удаления Са2+ в гладких мышцах


В гладкой мышце изменения уровней внутриклеточного Са2+ могут происходить как на фоне
В гладкой мышце изменения уровней внутриклеточного Са2+ могут происходить как на фоне

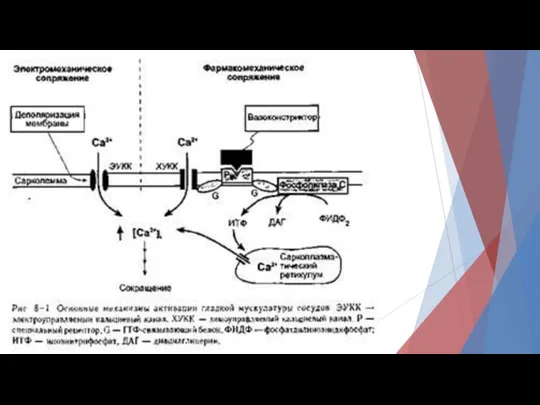
При фармако-механическом сопряжении химические факторы (например, нейромедиаторы) могут индуцировать сокращение гладкой
При фармако-механическом сопряжении химические факторы (например, нейромедиаторы) могут индуцировать сокращение гладкой
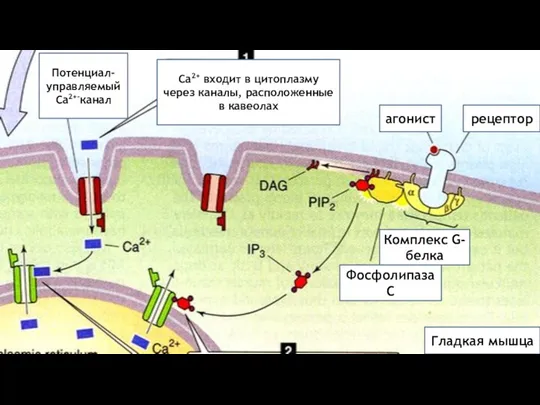
агонист
рецептор
Гладкая мышца
Комплекс G-белка
Фосфолипаза С
Потенциал-
управляемый
Са2+-канал
Са2+ входит в цитоплазму через каналы,
агонист
рецептор
Гладкая мышца
Комплекс G-белка
Фосфолипаза С
Потенциал-
управляемый
Са2+-канал
Са2+ входит в цитоплазму через каналы,

 Биосоциальная природа человека
Биосоциальная природа человека Виды животноводства
Виды животноводства Жасуша – тіршіліктің құрылымдық-функциялық бірлігі
Жасуша – тіршіліктің құрылымдық-функциялық бірлігі Использование в рационах кур-несушек высокобелковых растительных кормов взамен животных в ООО Птицефабрика Владимирская
Использование в рационах кур-несушек высокобелковых растительных кормов взамен животных в ООО Птицефабрика Владимирская Удобрения и их классификация
Удобрения и их классификация Корень. Внешнее и внутреннее строение корня
Корень. Внешнее и внутреннее строение корня Жизнь луга
Жизнь луга Влияние музыки на растения и животных
Влияние музыки на растения и животных Экологические факторы и их влияние на живые организмы
Экологические факторы и их влияние на живые организмы Подцарство многоклеточные. Тип Губки
Подцарство многоклеточные. Тип Губки Подготовка к ЕГЭ по биологии. 10 класс
Подготовка к ЕГЭ по биологии. 10 класс Понятия старения и старости. Теории старения. Организация гериатрической службы в Российской Федерации
Понятия старения и старости. Теории старения. Организация гериатрической службы в Российской Федерации Экологическая ниша и взаимоотношения организмов в биоценозе
Экологическая ниша и взаимоотношения организмов в биоценозе Энергетический обмен
Энергетический обмен Гербарии растений, содержащих эфирные масла. Часть 2
Гербарии растений, содержащих эфирные масла. Часть 2 Животные в зимнем лесу
Животные в зимнем лесу Радиопрограмма Неизвестная природа на уроках биологии в 6 классе.
Радиопрограмма Неизвестная природа на уроках биологии в 6 классе. Класс Двустворчатые моллюски
Класс Двустворчатые моллюски Размножение мхов
Размножение мхов Як вчаться пташенята
Як вчаться пташенята Царство животные. Тип моллюски. Тип членистоногие
Царство животные. Тип моллюски. Тип членистоногие Головний мозок
Головний мозок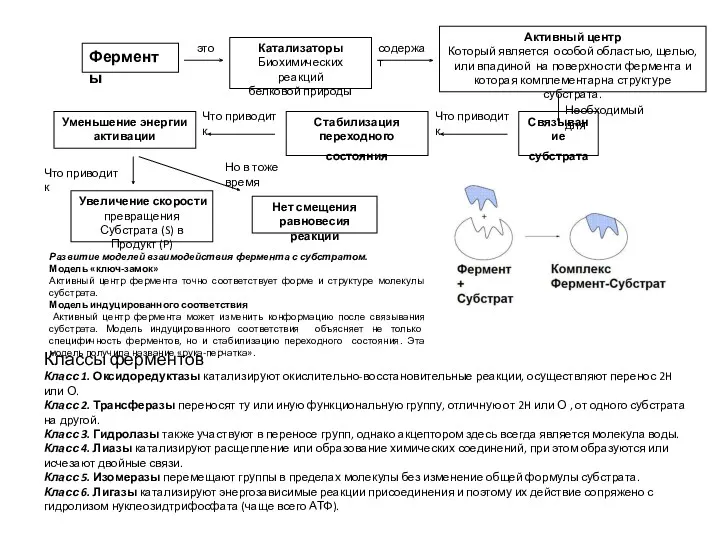 Ферменты. Тема 2.1
Ферменты. Тема 2.1 Методы селекции
Методы селекции Удивительный мир растений. 5 класс
Удивительный мир растений. 5 класс Приспособленность организмов и её относительность
Приспособленность организмов и её относительность Олень Карибу (Северный)
Олень Карибу (Северный) Орган зрения
Орган зрения