- Структура и функции нуклеиновых кислот. Репликация ДНК

Содержание
- 2. История изучения нуклеиновых кислот 1868 г. – открытие нуклеиновых кислот (Ф. Мишер) 1889 г. – введение
- 3. 1928 г. – эксперимент по трансформации бактерий Ф. Гриффита 1944 г. - О. Эвери, К. Маклеод
- 4. Функции нуклеиновых кислот Хранение наследственной информации Передача наследственной информации Реализация наследственной информации
- 5. Нуклеиновая кислота – это биополимер, мономерами которого являются нуклеотиды НК ДНК РНК дезоксирибонуклеиновая рибонуклеиновая кислота кислота
- 6. Пентоза ДНК РНК
- 7. АЗОТИСТЫЕ ОСНОВАНИЯ
- 9. Первичная структура НК Первичная структура нуклеиновых кислот – это последовательность нуклеотидов, соединенных ковалентными 3’-,5’- фосфодиэфирными связями.
- 10. Правила Чаргаффа (Э. Чаргафф, 1950 г.) [А] = [T] [G] = [C] [A + G] =
- 11. Рентгенограмма ДНК (Р. Франклин, 1953 г.)
- 12. Вторичная структура ДНК двойная спираль (Д. Уотсон и Ф. Крик, 1953 г.) Это две антипараллельные, комплементарные
- 14. Двойная спираль (В-тип) Большая бороздка Малая бороздка 1 оборот спирали = 3,4 нм (10 пар нуклеотидов)
- 15. Типы двойных спиралей
- 16. Третичная структура ДНК 1 – линейная, 2 – кольцевая одноцепочечная, 3 – кольцевая двухцепочечная молекулы.
- 17. Типы РНК м(и)РНК несут информацию о последовательности аминокислот в полипептидной цепочке. рРНК входят в состав рибосом.
- 18. Структура тРНК Вторичная Третичная «клеверный лист» L-форма
- 19. Репликация ДНК Репликация ДНК – это синтез ДНК на ДНК-матрице (удвоение ДНК) (М. Мезелсон, Ф. Сталь,
- 20. Подготовка ДНК-матрицы Ori-сайт - точка начала репликации (богатый АТ-парами участок ДНК, состоящий из 250-300 п.н.) +
- 21. Репликативная вилка 3’ 5’ 5’ 3’ топоизомераза геликаза SSB-белки
- 22. ДНК-полимеразы прокариот: ДНК-полимераза I полимераза (С-конец полипептидной цепи, или фрагмент Кленова) 3’-экзонуклеаза (С-конец полипептидной цепи, или
- 23. Особенности работы полимераз Катализируют реакцию (dNMP)n + dNTP → (dNMP)n+1 + PP Не могут осуществлять синтез
- 24. Ферменты репликации (этап синтеза) Праймаза относится к РНК-полимеразам, синтезирует праймер (РНК-затравку). ДНК-полимераза III синтезирует ведущую цепь
- 25. Особенности репликации у эукариот ДНК-полимеразы: ДНКП α участвует в синтезе праймеров. Процессивность низкая. ДНКП β –
- 26. Особенности репликации у эукариот Молекулы ДНК эукариот полирепликонные (у прокариот - монорепликонные). Длина фрагментов Оказаки –
- 27. Теломеры Теломеры - концевые участки хромосом, содержащие многократные повторы последовательности TTAGGG Теломераза – фермент, удлиняющий теломерную
- 29. Скачать презентацию

История изучения нуклеиновых кислот
1868 г. – открытие нуклеиновых кислот (Ф.
История изучения нуклеиновых кислот
1868 г. – открытие нуклеиновых кислот (Ф.

1928 г. – эксперимент по трансформации бактерий Ф. Гриффита
1944 г. -
1944 г. -

Функции нуклеиновых кислот
Хранение наследственной информации
Передача наследственной информации
Реализация наследственной информации
Функции нуклеиновых кислот
Хранение наследственной информации
Передача наследственной информации
Реализация наследственной информации
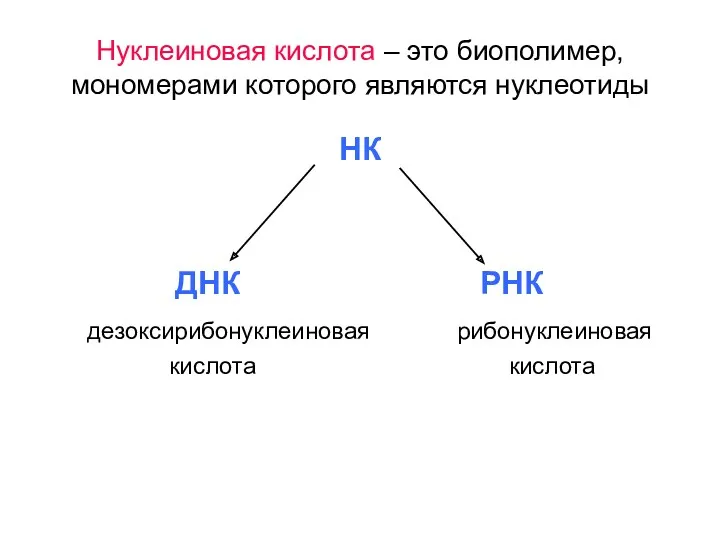
Нуклеиновая кислота – это биополимер, мономерами которого являются нуклеотиды
НК
ДНК РНК
дезоксирибонуклеиновая
Нуклеиновая кислота – это биополимер, мономерами которого являются нуклеотиды
НК
ДНК РНК
дезоксирибонуклеиновая
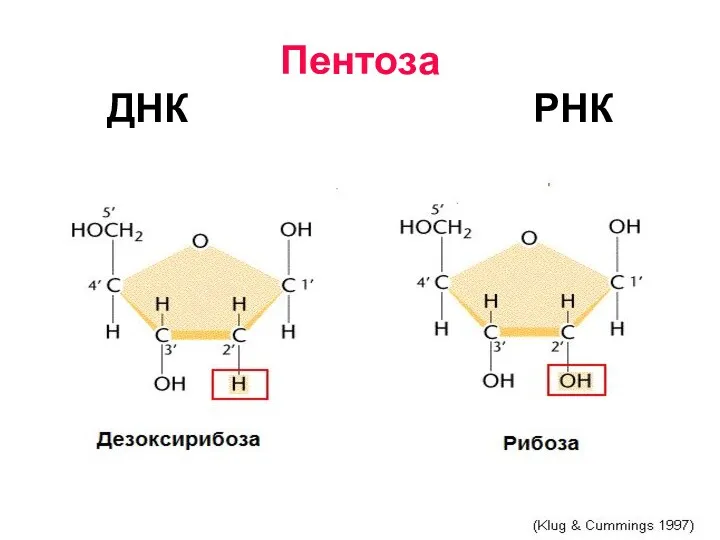
Пентоза
ДНК РНК
Пентоза
ДНК РНК
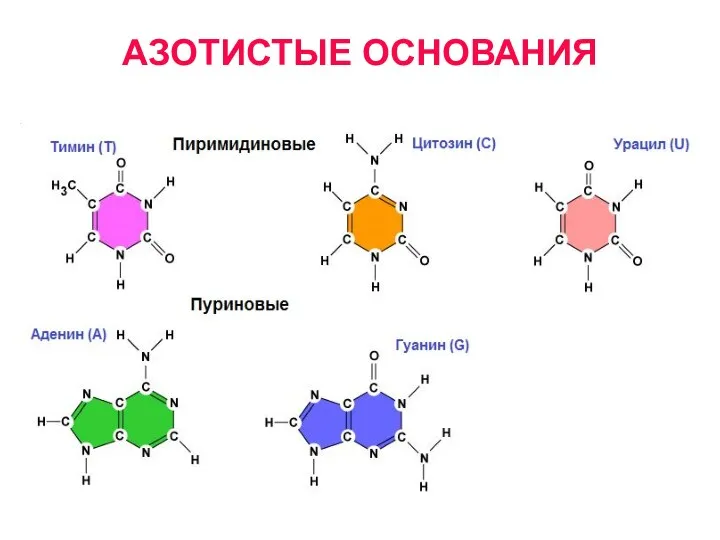
АЗОТИСТЫЕ ОСНОВАНИЯ
АЗОТИСТЫЕ ОСНОВАНИЯ
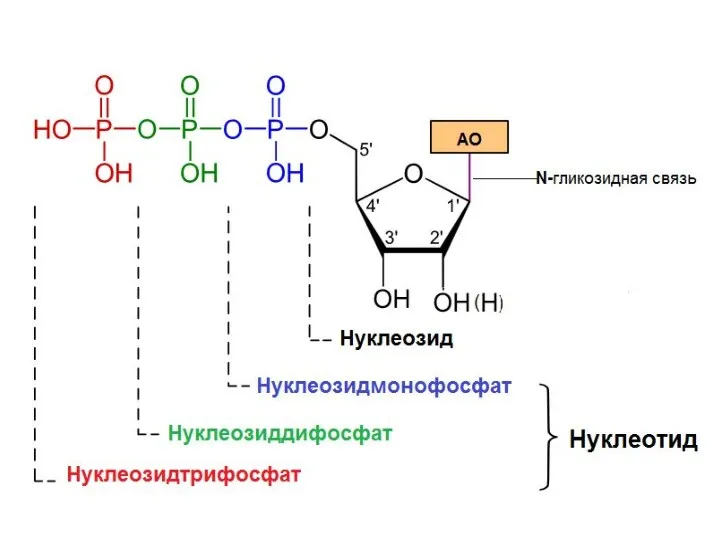
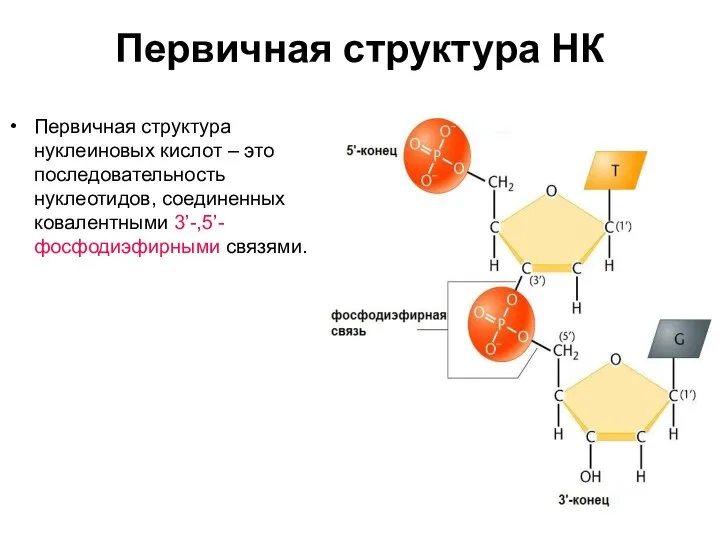
Первичная структура НК
Первичная структура нуклеиновых кислот – это последовательность нуклеотидов, соединенных
Первичная структура НК
Первичная структура нуклеиновых кислот – это последовательность нуклеотидов, соединенных
![Правила Чаргаффа (Э. Чаргафф, 1950 г.) [А] = [T] [G]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/399146/slide-9.jpg)
Правила Чаргаффа
(Э. Чаргафф, 1950 г.)
[А] = [T]
[G] = [C]
[A +
Правила Чаргаффа
(Э. Чаргафф, 1950 г.)
[А] = [T]
[G] = [C]
[A +
Рентгенограмма ДНК
(Р. Франклин, 1953 г.)
Рентгенограмма ДНК
(Р. Франклин, 1953 г.)

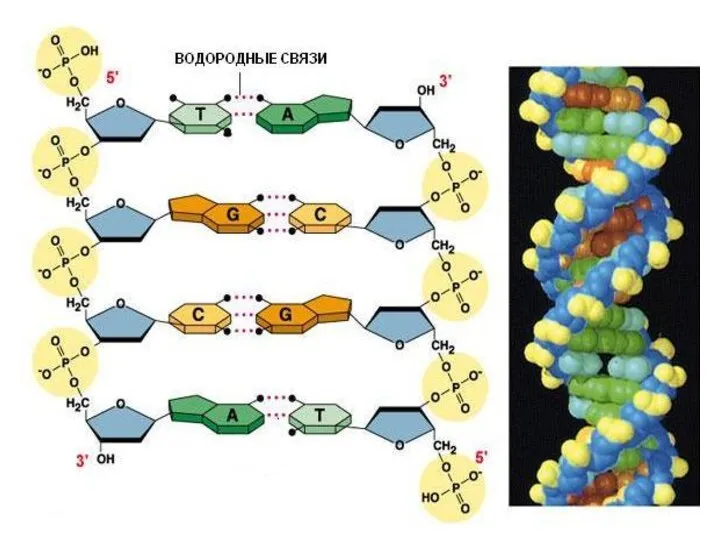
Вторичная структура ДНК
двойная спираль
(Д. Уотсон и Ф. Крик, 1953 г.)
Вторичная структура ДНК
двойная спираль
(Д. Уотсон и Ф. Крик, 1953 г.)
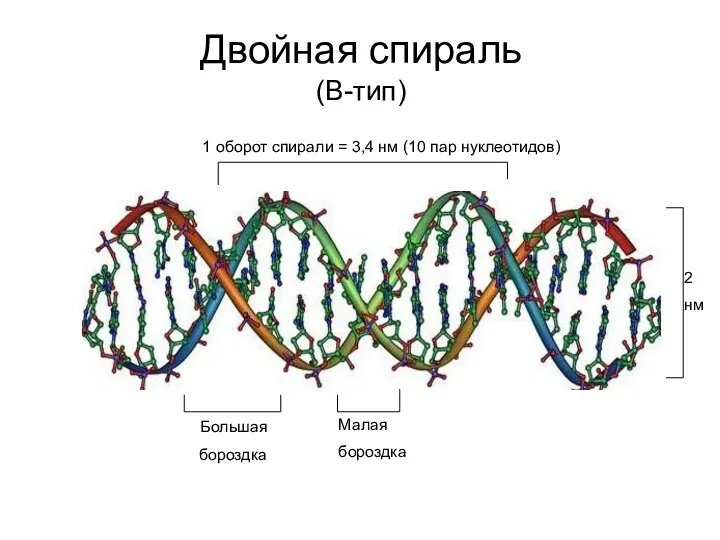
Двойная спираль
(В-тип)
Большая
бороздка
Малая
бороздка
1 оборот спирали = 3,4 нм (10
Двойная спираль
(В-тип)
Большая
бороздка
Малая
бороздка
1 оборот спирали = 3,4 нм (10
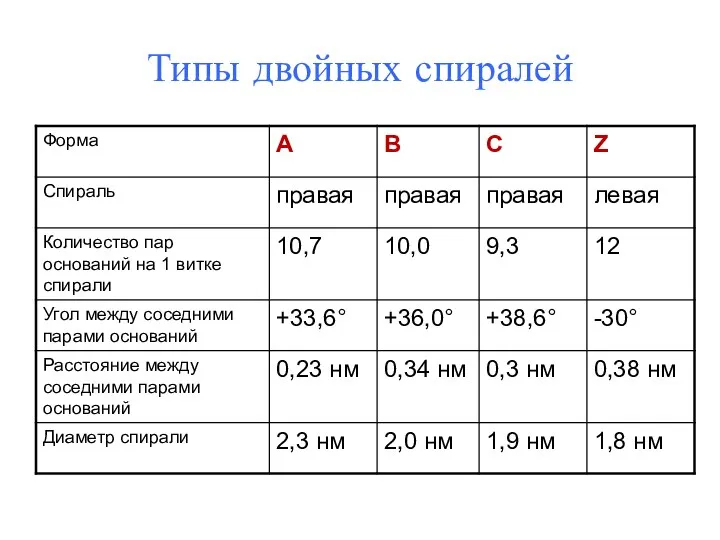
Типы двойных спиралей
Типы двойных спиралей
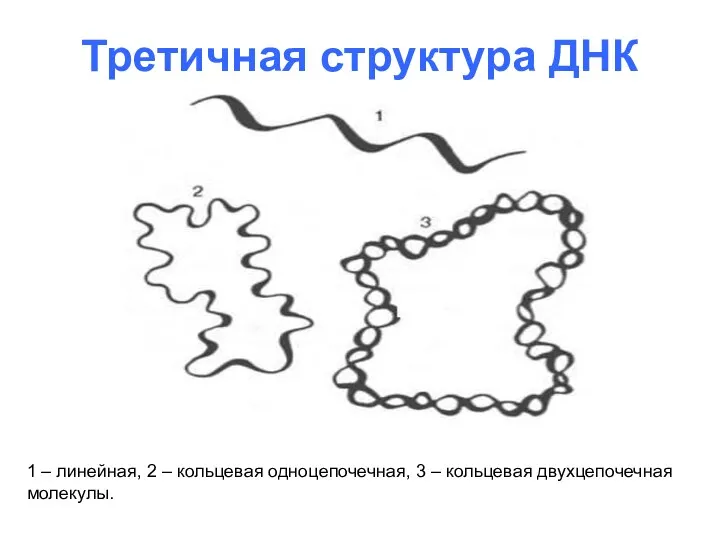
Третичная структура ДНК
1 – линейная, 2 – кольцевая одноцепочечная, 3 –
Третичная структура ДНК
1 – линейная, 2 – кольцевая одноцепочечная, 3 –

Типы РНК
м(и)РНК несут информацию о последовательности аминокислот в полипептидной цепочке.
рРНК входят
Типы РНК
м(и)РНК несут информацию о последовательности аминокислот в полипептидной цепочке.
рРНК входят
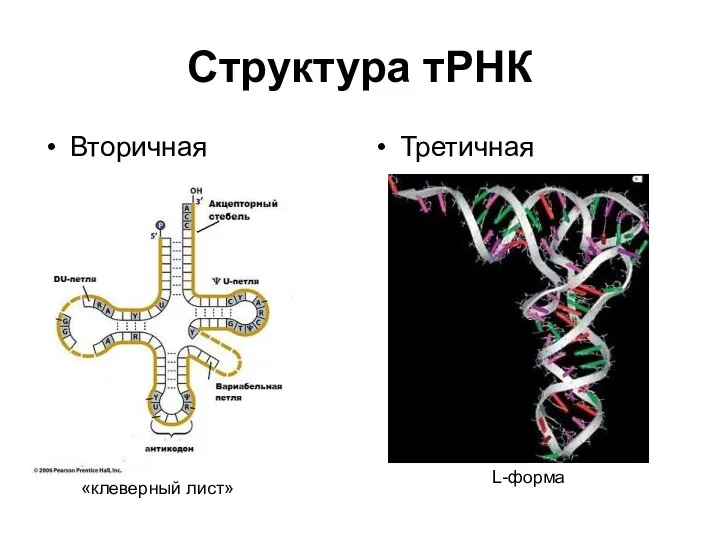
Структура тРНК
Вторичная
Третичная
«клеверный лист»
L-форма
Структура тРНК
Вторичная
Третичная
«клеверный лист»
L-форма

Репликация ДНК
Репликация ДНК – это синтез ДНК на ДНК-матрице (удвоение ДНК)
Репликация ДНК
Репликация ДНК – это синтез ДНК на ДНК-матрице (удвоение ДНК)

Подготовка ДНК-матрицы
Ori-сайт - точка начала репликации (богатый АТ-парами участок ДНК, состоящий
Подготовка ДНК-матрицы
Ori-сайт - точка начала репликации (богатый АТ-парами участок ДНК, состоящий
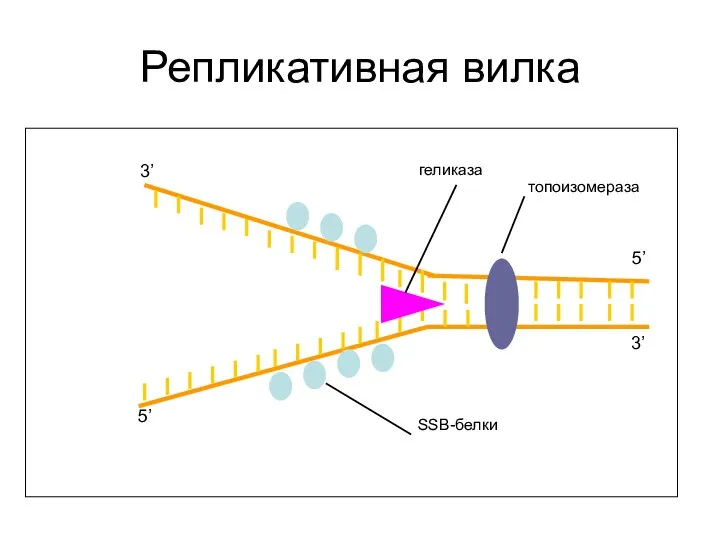
Репликативная вилка
3’
5’
5’
3’
топоизомераза
геликаза
SSB-белки
Репликативная вилка
3’
5’
5’
3’
топоизомераза
геликаза
SSB-белки

ДНК-полимеразы прокариот:
ДНК-полимераза I
полимераза (С-конец полипептидной цепи, или фрагмент Кленова)
3’-экзонуклеаза (С-конец
ДНК-полимеразы прокариот:
ДНК-полимераза I
полимераза (С-конец полипептидной цепи, или фрагмент Кленова)
3’-экзонуклеаза (С-конец

Особенности работы полимераз
Катализируют реакцию
(dNMP)n + dNTP → (dNMP)n+1 + PP
Не могут
Особенности работы полимераз
Катализируют реакцию
(dNMP)n + dNTP → (dNMP)n+1 + PP
Не могут

Ферменты репликации
(этап синтеза)
Праймаза относится к РНК-полимеразам, синтезирует праймер (РНК-затравку).
ДНК-полимераза III
Ферменты репликации
(этап синтеза)
Праймаза относится к РНК-полимеразам, синтезирует праймер (РНК-затравку).
ДНК-полимераза III

Особенности репликации у эукариот
ДНК-полимеразы:
ДНКП α участвует в синтезе праймеров. Процессивность низкая.
ДНКП
Особенности репликации у эукариот
ДНК-полимеразы:
ДНКП α участвует в синтезе праймеров. Процессивность низкая.
ДНКП

Особенности репликации у эукариот
Молекулы ДНК эукариот полирепликонные (у прокариот - монорепликонные).
Длина
Особенности репликации у эукариот
Молекулы ДНК эукариот полирепликонные (у прокариот - монорепликонные).
Длина

Теломеры
Теломеры - концевые участки хромосом, содержащие многократные повторы последовательности TTAGGG
Теломераза
Теломеры
Теломеры - концевые участки хромосом, содержащие многократные повторы последовательности TTAGGG
Теломераза
 Клеточная оболочка. Оболочка животных и растительных клеток
Клеточная оболочка. Оболочка животных и растительных клеток Вид, критерии вида. Популяция, подвид
Вид, критерии вида. Популяция, подвид Презентация по биологии для учащихся 7 класса по теме Класс Земноводные
Презентация по биологии для учащихся 7 класса по теме Класс Земноводные ПрезентацияОсновы учения о биосфере
ПрезентацияОсновы учения о биосфере Молочнокислые бактерии. 5 класс
Молочнокислые бактерии. 5 класс Устройство микроскопа и приёмы работы с ним. 5 класс
Устройство микроскопа и приёмы работы с ним. 5 класс Спектр белков и ферментов ротовой жидкости
Спектр белков и ферментов ротовой жидкости Дополнительная презентация Декоративные растения в интерьерек элективному курсу Основы фитодизайна, 9 класс
Дополнительная презентация Декоративные растения в интерьерек элективному курсу Основы фитодизайна, 9 класс Фармацевтична ботаніка. Аналіз буклету 2015 року
Фармацевтична ботаніка. Аналіз буклету 2015 року Открытый урок биологии по теме Движение 6 класс (УМК Н.И.Сонин) , автор учитель биологии Сизова С.В.
Открытый урок биологии по теме Движение 6 класс (УМК Н.И.Сонин) , автор учитель биологии Сизова С.В. Основные понятия генетики
Основные понятия генетики Тип Плоские черви
Тип Плоские черви Анатомия, физиология и патология органа слуха
Анатомия, физиология и патология органа слуха Игра Насекомые
Игра Насекомые Бактерии в организме человека
Бактерии в организме человека Схема - строение светового микроскопа
Схема - строение светового микроскопа Метаболизмнің гормондық реттелуі. Қоршаған және өндірістік орта факторларының әсер ету бағасының биохимиялық критерийлері
Метаболизмнің гормондық реттелуі. Қоршаған және өндірістік орта факторларының әсер ету бағасының биохимиялық критерийлері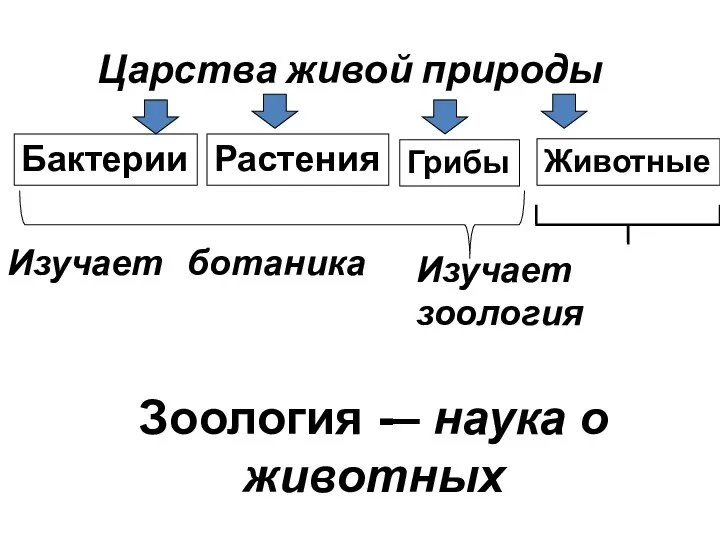 Царства живой природы
Царства живой природы Генетикалық инженерия негіздері
Генетикалық инженерия негіздері Бионика органов чувств человека (9 класс)
Бионика органов чувств человека (9 класс) Красная книга Самарской области
Красная книга Самарской области მცენარეების გავრცელების გზები
მცენარეების გავრცელების გზები Условия прорастания семян растений
Условия прорастания семян растений Кошки и собаки
Кошки и собаки Оболочки головного и спинного мозга
Оболочки головного и спинного мозга Общая характеристика пресмыкающихся
Общая характеристика пресмыкающихся Собака в нашем доме (часть 2)
Собака в нашем доме (часть 2) Protein denatu-ration
Protein denatu-ration