- Структурная организация белка

Содержание
- 2. История изучения Белки были выделены в отдельный класс биологических молекулБелки были выделены в отдельный класс биологических
- 3. Мономерами белков являются аминокислоты карбоксильная группа радикал аминогруппа
- 4. Олигопептиды содержат до 20 амк (среди них различают ди-, три-, тетра- и т.д. пептиды); Полипептиды содержат
- 5. Образование пептидной связи Схематическое изображение образования пептидной связи. Подобная реакция происходит в молекулярной машине по образованию
- 6. Свойства белков Размер белка может измеряться в числе аминокислот или в дальтонахРазмер белка может измеряться в
- 7. - Амфотерность - Способность к ионизации в растворе - Характерной константой белков является рI – изоэлектическая
- 8. Классификация белков По химическому строению простые; сложные; По выполняемой функции белки-ферменты белки-гормоны белки-рецепторы структурные белки защитные
- 9. Классификация белков По молекулярной массе низкомолекулярные высокомолекулярные По локализации в клетке цитоплазматические лизосомальные ядерные и т.д.
- 10. Классификация белков По молекулярной массе низкомолекулярные Высокомолекулярные По локализации в клетке цитоплазматические лизосомальные ядерные и т.д.
- 11. Классификация белков По возможности адаптативно регулировать количество данных белков белки, синтезирующиеся с постоянной скоростью (конститутивные) белки,
- 12. Особенности первичной структуры В остове полипептидной цепи чередуются жесткие структуры (плоские пептидные группы) с относительно подвижными
- 13. Пептидная связь – образуется при взаимодействии аминогруппы одной аминокислоты с карбоксильной группой другой аминокислоты с выделением
- 14. Первичная структура белка Последовательность чередования аминокислотных остатков (все связи ковалентные – пептидные, прочные). Пептидные связи стабилизируют
- 15. Особенности первичной структуры В остове полипептидной цепи чередуются жесткие структуры (плоские пептидные группы) с относительно подвижными
- 16. Особенности пептидной связи Компланарность – все атомы, входящие в пептидную группу находятся в одной плоскости; Способность
- 17. Последовательность изучения первичной структуры белков. Расщепление полипептидной цепи белка на более короткие фрагменты по определенным положениям
- 18. К слабым взаимодействиям относят: Водородные связи – внутри-и межмолекулярные (эти связи формируют гидрофильные радикалы); Ионные связи
- 19. Вторичная структура Вторичная структура это пространственная конфигурация полипептидных цепей, которые стремятся уменьшить свободную энергию, то есть
- 20. α спирализация – это закручивание полипептидно остова вокруг мнимой оси цилиндра по ходу часовой стрелки, поскольку
- 21. ά-спираль Водородные связи формируют трехмерную вторичную структуру белка и образуются между карбонильной (>С=О) и иминогруппами (=NH)
- 22. Спираль-клубочек Содержание ά-спирали в белках неодинаково и является индивидуальной особенностью каждой белковой молекулы. Глобулярные белки имеют
- 23. Основные особенности ά-спирали Спиральная конфигурация полипептидной цепи имеет винтовую симметрию (напоминает растянутую спираль электрической плитки); Образование
- 24. Некоторые фибриллярные белки образуют конформацию β – структуры- структуры складчатого листа, то есть, последовательный ряд листков,
- 25. β-структура – Складчатого типа, водородные связи образуются менее системно, формируя гофрированную структуру из полипептидной цепи. При
- 26. N-конец С-конец Параллельная β-складчатая укладка β-поворот N-конец С-конец Антипараллельная β-складчатая укладка – боковые радикалы одного слоя
- 27. β – структура
- 29. Связи, поддерживающие вторичную структуру: водородные электростатические гидрофобные ван-дер-ваальсовые
- 30. Водородные: 1) N - H H - C - R R – C - H C=O
- 31. Домены Это более сложные уровни организации вторичной структуры. Они представляют собой обособленные глобулярные участки, соединенные др.
- 32. Доменная организация характерна для многих белков. В этих белках находится, как правило несколько структурных доменов, каждый
- 33. Третичная структура- Третичная структура- это способ укладки полипептидной цепи в определенном объеме пространства.
- 34. Третичная структура поддерживается большим числом слабых связей, энергия которых мала от 1 до 7 кал. Слабые
- 35. Третичная структура белка Это реальная трехмерная конфигурация, которую принимает в пространстве закрученная спираль (за счет гидрофобных
- 36. Связи, стабилизирующие третичную структуру белка Ковалентные - сильные: дисульфидные (-S-S-боковые радикалы цис); изопептидные или псевдопептидные –
- 37. 2. Полярные – слабые: водородные – между группой –NH2, -OH,-SH бокового радикала одной амк и СООН-группой
- 38. Особенности организации третичной структуры. Конформация третичной структуры полипептидной цепи определяется свойствами боковых радикалов входящих в нее
- 39. Уровни структурной организации белка
- 40. Четвертичная структура Четвертичная структура – это способ укладки в пространстве отдельных полипептидных цепей, обладающих одинаковой или
- 41. Стабилизация четвертичной структуры белков. В стабилизации четвертичной структуры принимают участие те же типы взаимодействий, что и
- 43. Структура белка Кроме последовательности аминокислот полипептида (первичной структуры), крайне важна трёхмерная структура белка, которая формируется в
- 44. Гемоглобин
- 45. Гемоглобин
- 46. Кооперативные изменения конформации протомеров О2 связывается с протомерами Нв через Fe2+, который соединен с 4 атомами
- 47. Кооперативные изменения конформации протомеров Нв при присоединении О2 Белки обладают конформационной лабильностью и после перемещения железа
- 48. Гем присоединяется к неполярным радикалам активного центра своими пиррольными циклами, а также к радикалу гистидина с
- 49. Эмбриональный и фетальный гемоглобин Эмбриональный гемоглобин синтезируется в эмбриональном желточном мешке через несколько недель после оплодотворения.
- 50. Формы гемоглобина Оксигемоглобин (НвО2) – полностью оксигенированный Нв; Дезоксигемоглобин (Нв) – незамещенная 6 координационная связь железа;
- 51. Строение протомеров гемоглобина Аллостерический центр Каждая субъединица имеет центр связывания, где располагается небелковая часть молекулы –
- 52. Таким образом: 1. в центре тетрамерной молекулы Нв находится полость, которую образуют амк остатки всех 4
- 53. 2,3-Бисфосоглицерат (БФГ) – вещество, синтезируемое в Эр из промежуточного продукта окисления глюкозы – 1,3-бисфосфоглицерата. В нормальных
- 54. Изменение концентрации БФГ – как механизм адаптации организма к гипоксии. Концентрация БФГ в Эр людей, живущих
- 55. Аллостерическими регуляторами активности Нв, которые присоединяются к аллостерическим центрам (пространственно удаленным от активного центра), помимо 2,3-БФГ
- 56. Перенос Н+ и СО2 из тканей в легкие с помощью гемоглобина. Эффект Бора. Увеличение освобождения О2
- 57. Оксигенирование дезоксигемоглобина в легких, образование и выделение СО2. Увеличение концентрации протонов в среде снижает сродство О2
- 59. Конформационные изменения гемоглобина при присоединении кислорода
- 60. Изменение конформации гемоглобина в результате связывания с кислородом
- 61. Кривая насыщения гемоглобина кислородом
- 62. Виды гемоглобина человека Гемоглобин А (НвА) - α 2β2 - основной тип гемоглобина у взрослых Гемоглобин
- 64. Скачать презентацию

История изучения
Белки были выделены в отдельный класс биологических молекулБелки были выделены
История изучения
Белки были выделены в отдельный класс биологических молекулБелки были выделены
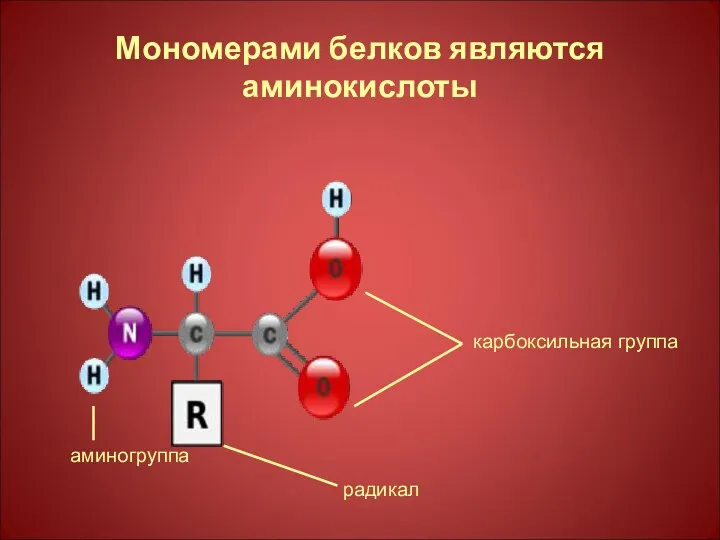
Мономерами белков являются аминокислоты
карбоксильная группа
радикал
аминогруппа
Мономерами белков являются аминокислоты
карбоксильная группа
радикал
аминогруппа

Олигопептиды содержат до 20 амк (среди них различают ди-, три-, тетра-
Олигопептиды содержат до 20 амк (среди них различают ди-, три-, тетра-
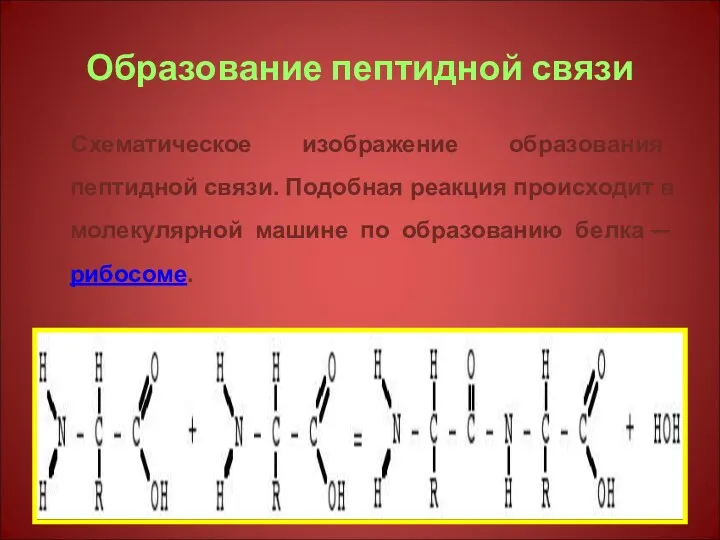
Образование пептидной связи
Схематическое изображение образования пептидной связи. Подобная реакция происходит в
Образование пептидной связи
Схематическое изображение образования пептидной связи. Подобная реакция происходит в
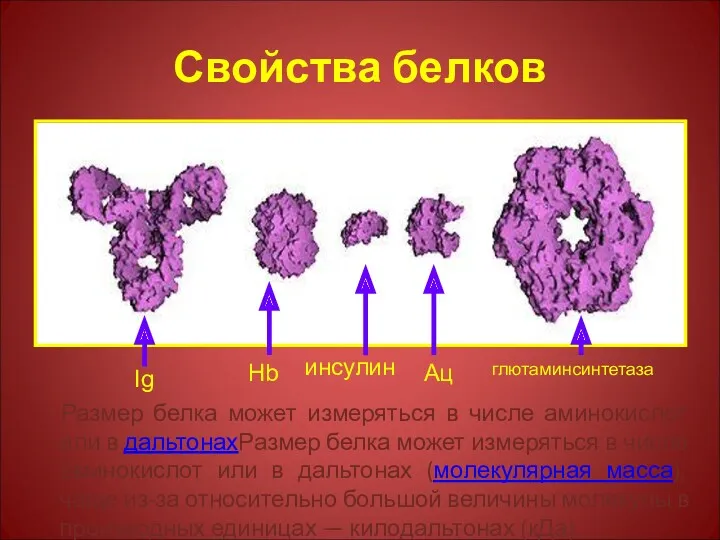
Свойства белков
Размер белка может измеряться в числе аминокислот или в дальтонахРазмер
Свойства белков
Размер белка может измеряться в числе аминокислот или в дальтонахРазмер

- Амфотерность
- Способность к ионизации в растворе
- Характерной константой белков является
- Амфотерность - Способность к ионизации в растворе - Характерной константой белков является

Классификация белков
По химическому строению
простые;
сложные;
По выполняемой функции
белки-ферменты
белки-гормоны
белки-рецепторы
структурные белки
защитные белки
сократительные белки
По форме
глобулярные
фибриллярные
Классификация белков
По химическому строению
простые;
сложные;
По выполняемой функции
белки-ферменты
белки-гормоны
белки-рецепторы
структурные белки
защитные белки
сократительные белки
По форме
глобулярные
фибриллярные

Классификация белков
По молекулярной массе
низкомолекулярные
высокомолекулярные
По локализации в клетке
цитоплазматические
лизосомальные
ядерные и т.д.
По локализации в
Классификация белков
По молекулярной массе
низкомолекулярные
высокомолекулярные
По локализации в клетке
цитоплазматические
лизосомальные
ядерные и т.д.
По локализации в

Классификация белков
По молекулярной массе
низкомолекулярные
Высокомолекулярные
По локализации в клетке
цитоплазматические
лизосомальные
ядерные и т.д.
По локализации в
Классификация белков
По молекулярной массе
низкомолекулярные
Высокомолекулярные
По локализации в клетке
цитоплазматические
лизосомальные
ядерные и т.д.
По локализации в

Классификация белков
По возможности адаптативно регулировать количество данных белков
белки, синтезирующиеся с
Классификация белков
По возможности адаптативно регулировать количество данных белков
белки, синтезирующиеся с
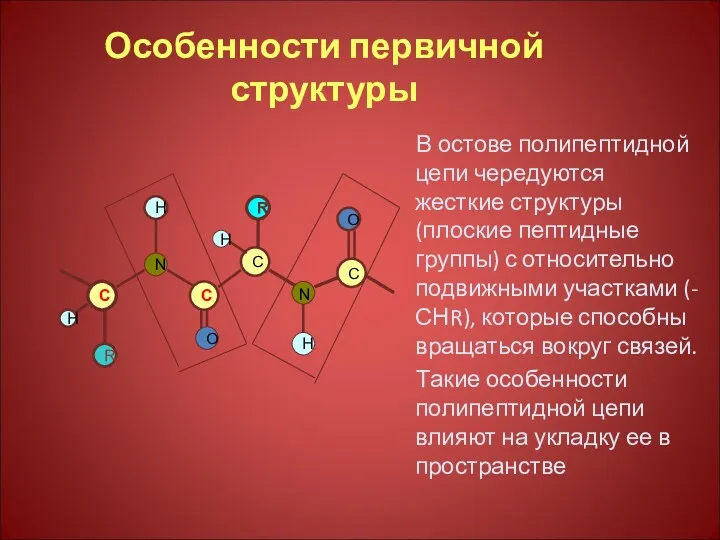
Особенности первичной структуры
В остове полипептидной цепи чередуются жесткие структуры (плоские пептидные
Особенности первичной структуры
В остове полипептидной цепи чередуются жесткие структуры (плоские пептидные

Пептидная связь – образуется при взаимодействии аминогруппы одной аминокислоты с карбоксильной
Пептидная связь – образуется при взаимодействии аминогруппы одной аминокислоты с карбоксильной
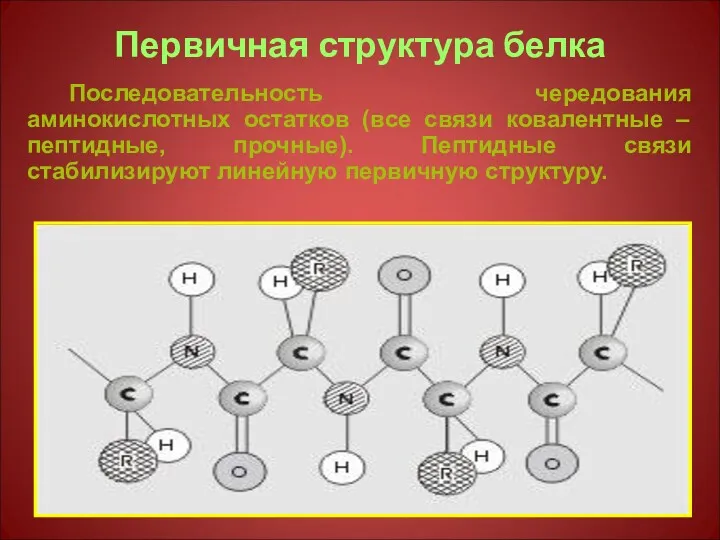
Первичная структура белка
Последовательность чередования аминокислотных остатков (все связи ковалентные – пептидные,
Первичная структура белка
Последовательность чередования аминокислотных остатков (все связи ковалентные – пептидные,
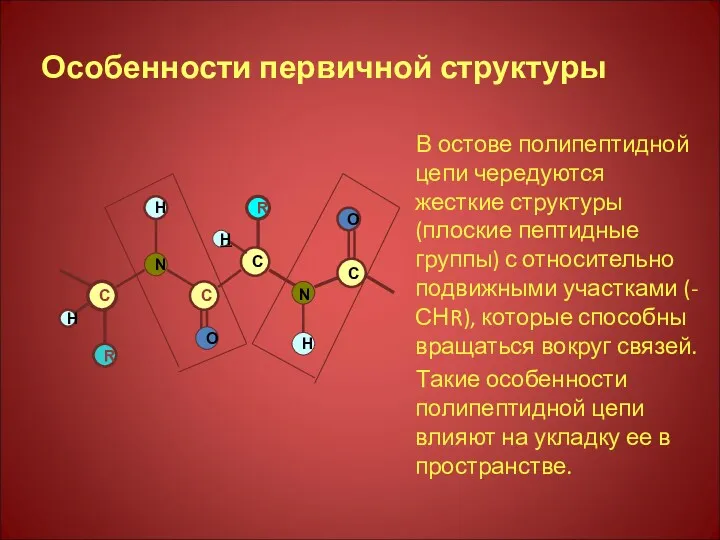
Особенности первичной структуры
В остове полипептидной цепи чередуются жесткие структуры (плоские пептидные
Особенности первичной структуры
В остове полипептидной цепи чередуются жесткие структуры (плоские пептидные

Особенности пептидной связи
Компланарность – все атомы, входящие в пептидную группу находятся
Особенности пептидной связи
Компланарность – все атомы, входящие в пептидную группу находятся

Последовательность изучения первичной структуры белков.
Расщепление полипептидной цепи белка на более короткие
Последовательность изучения первичной структуры белков.
Расщепление полипептидной цепи белка на более короткие

К слабым взаимодействиям относят:
Водородные связи – внутри-и межмолекулярные (эти связи формируют
К слабым взаимодействиям относят:
Водородные связи – внутри-и межмолекулярные (эти связи формируют

Вторичная структура
Вторичная структура это пространственная конфигурация полипептидных цепей, которые стремятся
Вторичная структура
Вторичная структура это пространственная конфигурация полипептидных цепей, которые стремятся
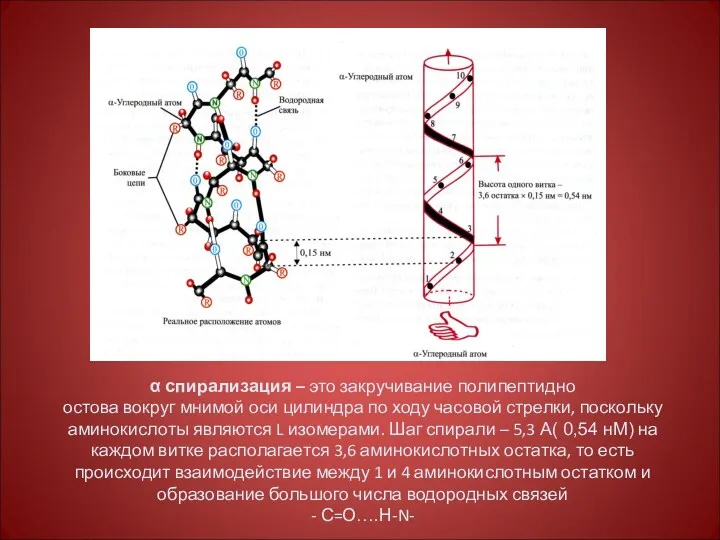
α спирализация – это закручивание полипептидно
остова вокруг мнимой оси цилиндра по
α спирализация – это закручивание полипептидно
остова вокруг мнимой оси цилиндра по

ά-спираль
Водородные связи формируют трехмерную вторичную структуру белка и образуются между карбонильной
ά-спираль
Водородные связи формируют трехмерную вторичную структуру белка и образуются между карбонильной

Спираль-клубочек
Содержание ά-спирали в белках неодинаково и является индивидуальной особенностью каждой белковой
Спираль-клубочек
Содержание ά-спирали в белках неодинаково и является индивидуальной особенностью каждой белковой

Основные особенности ά-спирали
Спиральная конфигурация полипептидной цепи имеет винтовую симметрию (напоминает растянутую
Основные особенности ά-спирали
Спиральная конфигурация полипептидной цепи имеет винтовую симметрию (напоминает растянутую

Некоторые фибриллярные белки образуют конформацию β – структуры- структуры складчатого листа,
Некоторые фибриллярные белки образуют конформацию β – структуры- структуры складчатого листа,

β-структура –
Складчатого типа, водородные связи образуются менее системно, формируя гофрированную
β-структура –
Складчатого типа, водородные связи образуются менее системно, формируя гофрированную
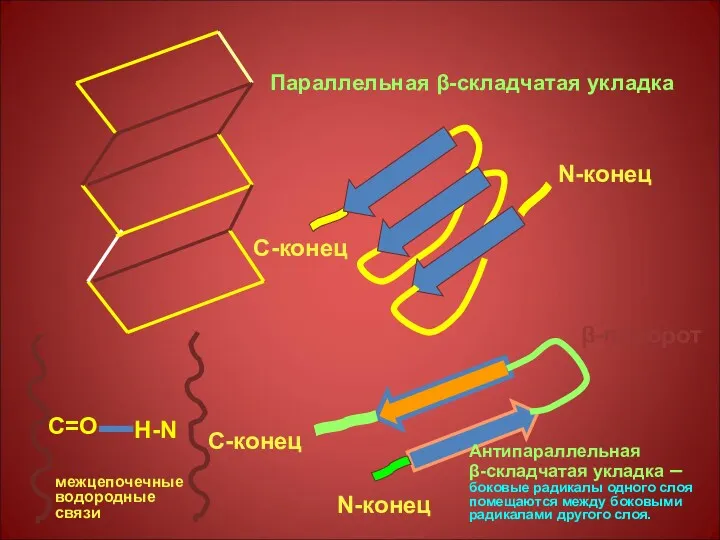
N-конец
С-конец
Параллельная β-складчатая укладка
β-поворот
N-конец
С-конец
Антипараллельная
β-складчатая укладка – боковые радикалы одного слоя помещаются
N-конец
С-конец
Параллельная β-складчатая укладка
β-поворот
N-конец
С-конец
Антипараллельная
β-складчатая укладка – боковые радикалы одного слоя помещаются
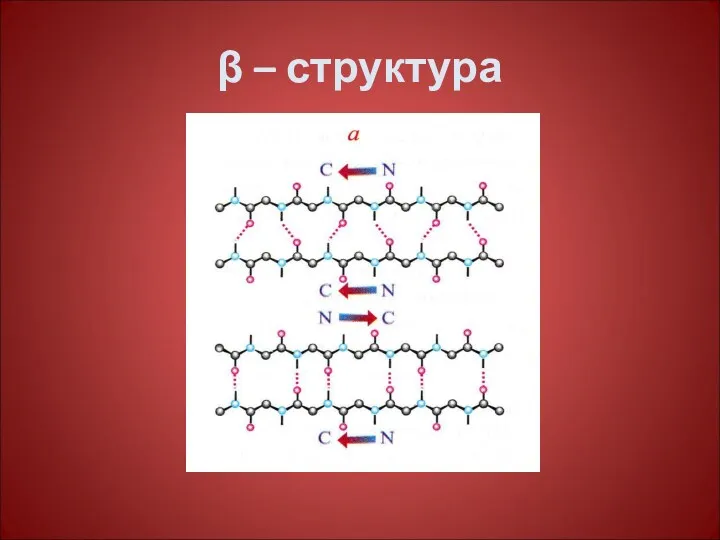
β – структура
β – структура
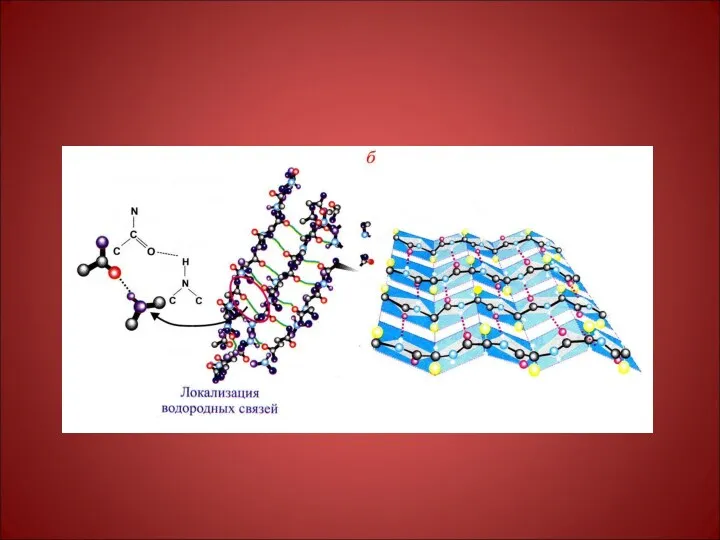

Связи, поддерживающие
вторичную структуру:
водородные
электростатические
гидрофобные
ван-дер-ваальсовые
Связи, поддерживающие
вторичную структуру:
водородные
электростатические
гидрофобные
ван-дер-ваальсовые

Водородные:
1) N - H H - C - R
R – C
Водородные:
1) N - H H - C - R
R – C

Домены
Это более сложные уровни организации вторичной структуры. Они представляют собой
Домены
Это более сложные уровни организации вторичной структуры. Они представляют собой

Доменная организация характерна для многих белков. В этих белках находится, как
Доменная организация характерна для многих белков. В этих белках находится, как

Третичная структура-
Третичная структура- это способ укладки полипептидной цепи в определенном объеме
Третичная структура-
Третичная структура- это способ укладки полипептидной цепи в определенном объеме
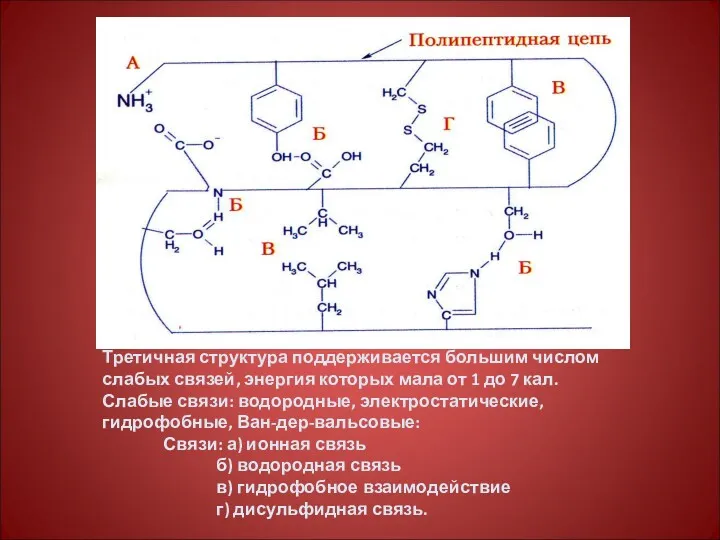
Третичная структура поддерживается большим числом слабых связей, энергия которых мала от
Третичная структура поддерживается большим числом слабых связей, энергия которых мала от

Третичная структура белка
Это реальная трехмерная конфигурация, которую принимает в пространстве закрученная
Третичная структура белка
Это реальная трехмерная конфигурация, которую принимает в пространстве закрученная

Связи, стабилизирующие третичную структуру белка
Ковалентные - сильные:
дисульфидные (-S-S-боковые радикалы цис);
изопептидные или
Связи, стабилизирующие третичную структуру белка
Ковалентные - сильные:
дисульфидные (-S-S-боковые радикалы цис);
изопептидные или

2. Полярные – слабые:
водородные – между группой –NH2, -OH,-SH бокового радикала
2. Полярные – слабые: водородные – между группой –NH2, -OH,-SH бокового радикала

Особенности организации третичной структуры.
Конформация третичной структуры полипептидной цепи определяется свойствами боковых
Особенности организации третичной структуры.
Конформация третичной структуры полипептидной цепи определяется свойствами боковых
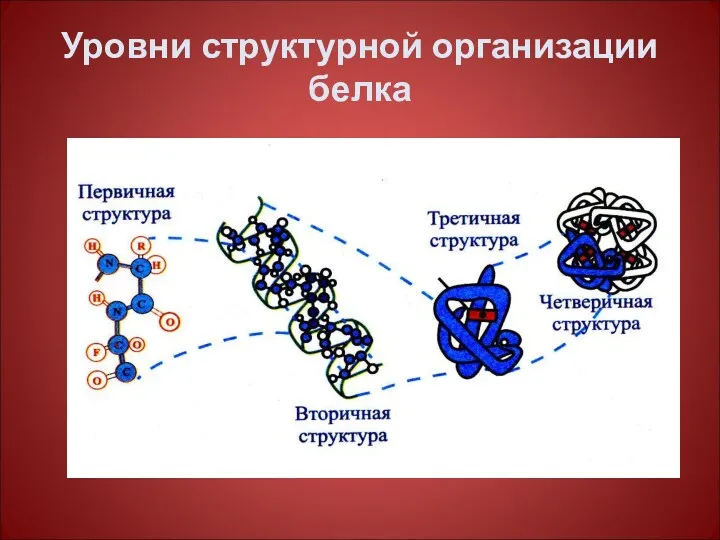
Уровни структурной организации белка
Уровни структурной организации белка

Четвертичная структура
Четвертичная структура – это способ укладки в пространстве отдельных полипептидных
Четвертичная структура
Четвертичная структура – это способ укладки в пространстве отдельных полипептидных

Стабилизация четвертичной структуры белков.
В стабилизации четвертичной структуры принимают участие те же
Стабилизация четвертичной структуры белков.
В стабилизации четвертичной структуры принимают участие те же
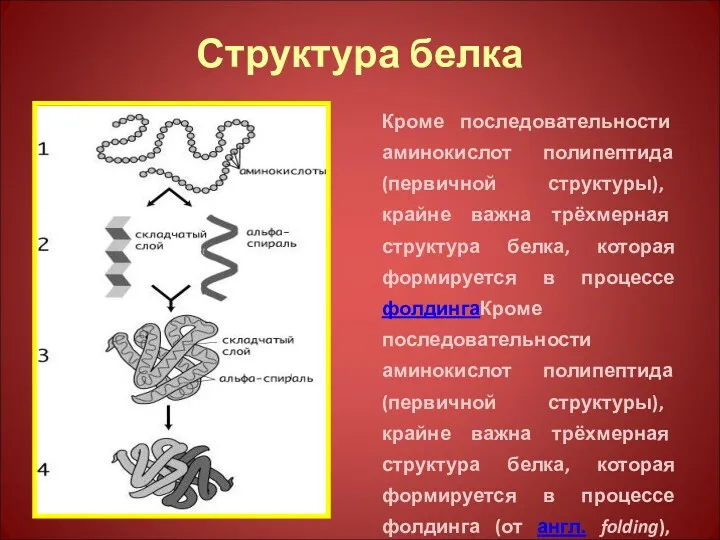
Структура белка
Кроме последовательности аминокислот полипептида (первичной структуры), крайне важна трёхмерная структура
Структура белка
Кроме последовательности аминокислот полипептида (первичной структуры), крайне важна трёхмерная структура

Гемоглобин
Гемоглобин
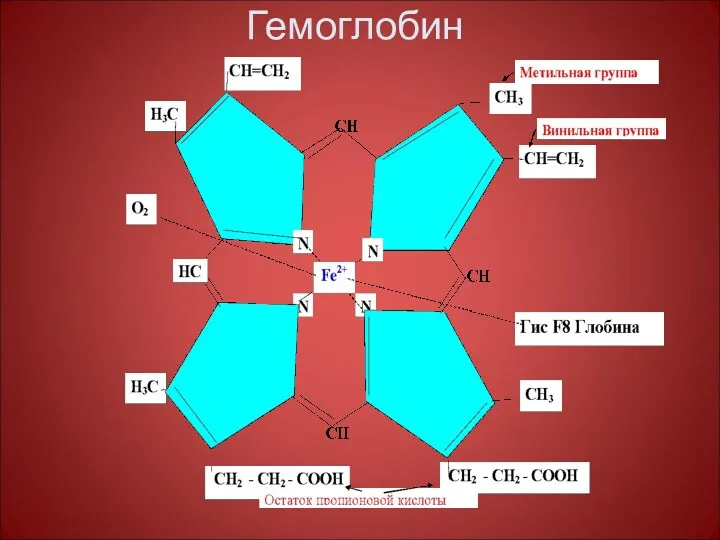
Гемоглобин
Гемоглобин
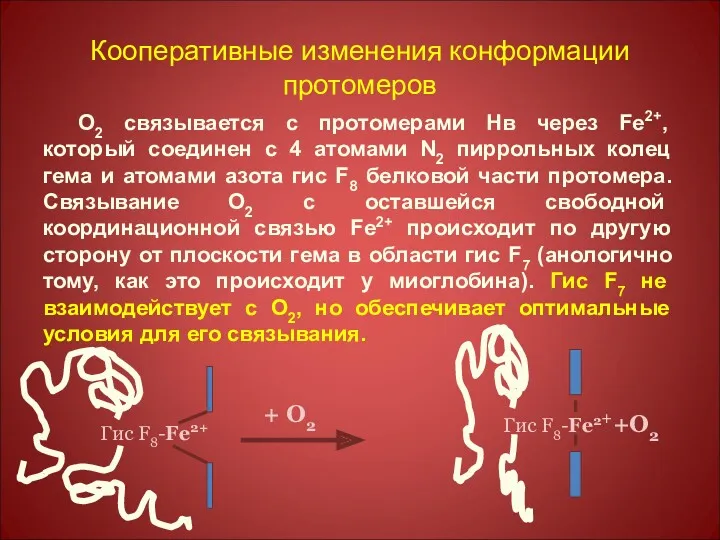
Кооперативные изменения конформации протомеров
О2 связывается с протомерами Нв через Fe2+, который
Кооперативные изменения конформации протомеров
О2 связывается с протомерами Нв через Fe2+, который
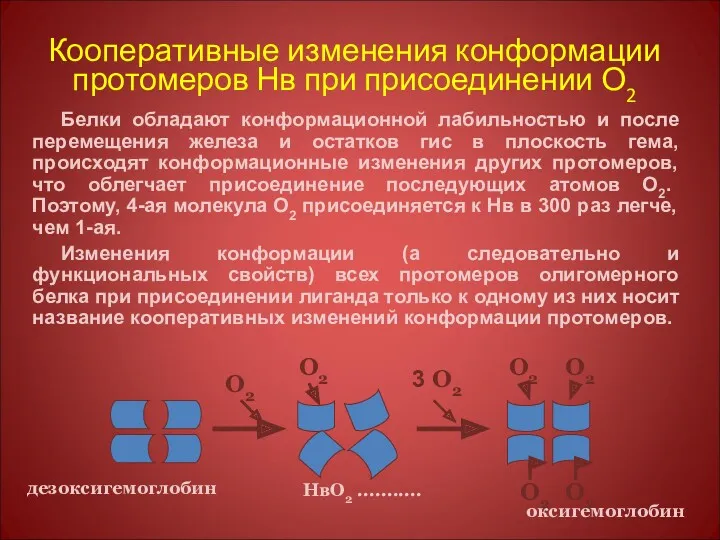
Кооперативные изменения конформации протомеров Нв при присоединении О2
Белки обладают конформационной лабильностью
Кооперативные изменения конформации протомеров Нв при присоединении О2
Белки обладают конформационной лабильностью

Гем присоединяется к неполярным радикалам активного центра своими пиррольными циклами, а
Гем присоединяется к неполярным радикалам активного центра своими пиррольными циклами, а

Эмбриональный и фетальный гемоглобин
Эмбриональный гемоглобин синтезируется в эмбриональном желточном мешке через
Эмбриональный и фетальный гемоглобин
Эмбриональный гемоглобин синтезируется в эмбриональном желточном мешке через

Формы гемоглобина
Оксигемоглобин (НвО2) – полностью оксигенированный Нв;
Дезоксигемоглобин (Нв) – незамещенная 6
Формы гемоглобина
Оксигемоглобин (НвО2) – полностью оксигенированный Нв;
Дезоксигемоглобин (Нв) – незамещенная 6
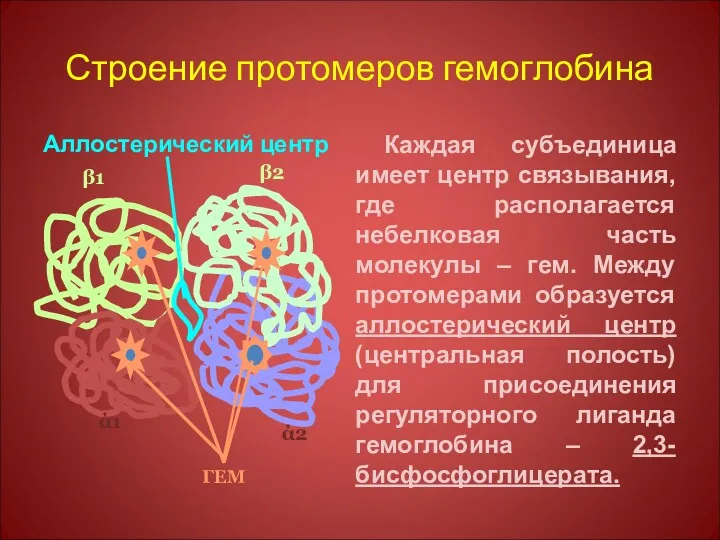
Строение протомеров гемоглобина
Аллостерический центр
Каждая субъединица имеет центр связывания, где располагается небелковая
Строение протомеров гемоглобина
Аллостерический центр
Каждая субъединица имеет центр связывания, где располагается небелковая

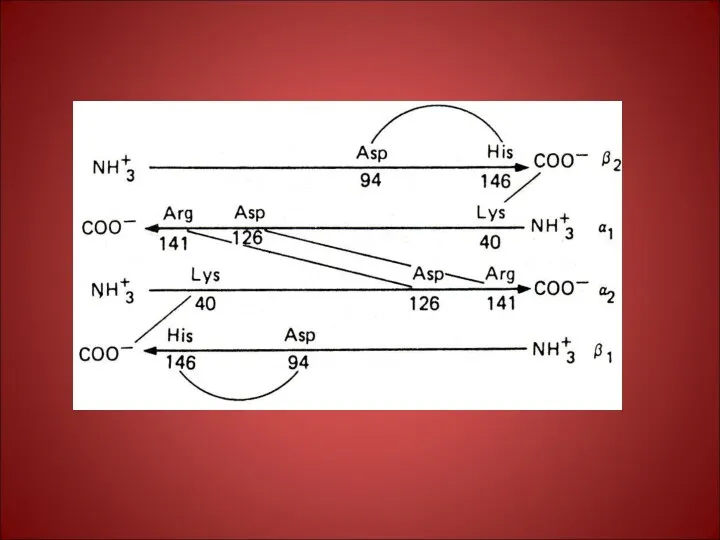
Таким образом:
1. в центре тетрамерной молекулы Нв находится полость, которую образуют
Таким образом: 1. в центре тетрамерной молекулы Нв находится полость, которую образуют

2,3-Бисфосоглицерат (БФГ) – вещество, синтезируемое в Эр из промежуточного продукта окисления
2,3-Бисфосоглицерат (БФГ) – вещество, синтезируемое в Эр из промежуточного продукта окисления

Изменение концентрации БФГ – как механизм адаптации организма к гипоксии.
Концентрация БФГ
Изменение концентрации БФГ – как механизм адаптации организма к гипоксии.
Концентрация БФГ

Аллостерическими регуляторами активности Нв, которые присоединяются к аллостерическим центрам (пространственно удаленным
Аллостерическими регуляторами активности Нв, которые присоединяются к аллостерическим центрам (пространственно удаленным
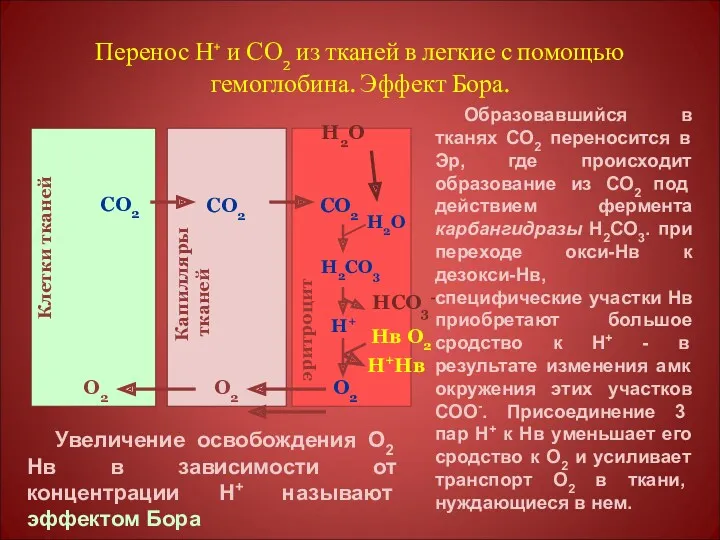
Перенос Н+ и СО2 из тканей в легкие с помощью гемоглобина.
Перенос Н+ и СО2 из тканей в легкие с помощью гемоглобина.

Оксигенирование дезоксигемоглобина в легких, образование и выделение СО2.
Увеличение концентрации протонов в
Оксигенирование дезоксигемоглобина в легких, образование и выделение СО2.
Увеличение концентрации протонов в
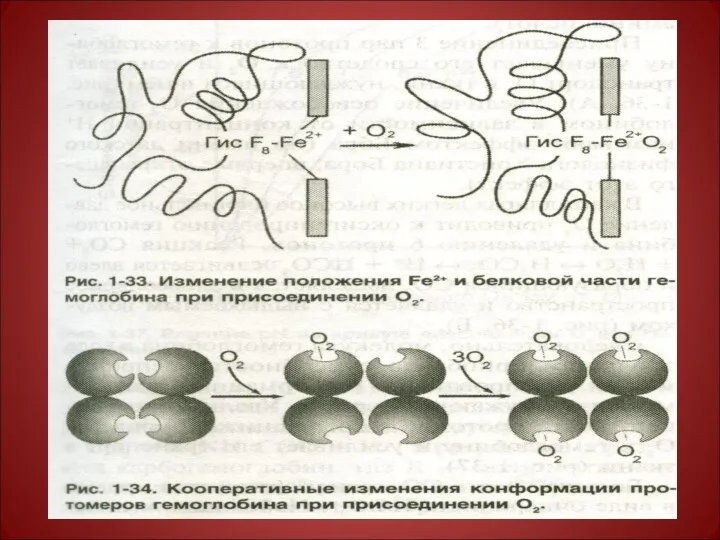
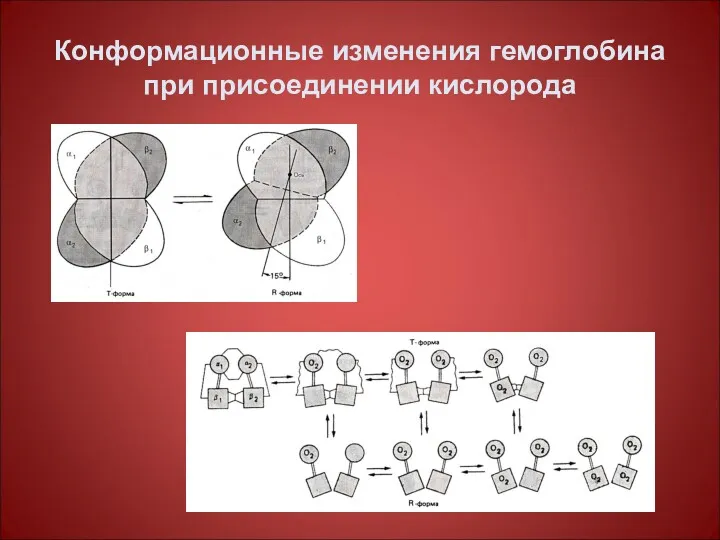
Конформационные изменения гемоглобина при присоединении кислорода
Конформационные изменения гемоглобина при присоединении кислорода

Изменение конформации гемоглобина в результате связывания с кислородом
Изменение конформации гемоглобина в результате связывания с кислородом
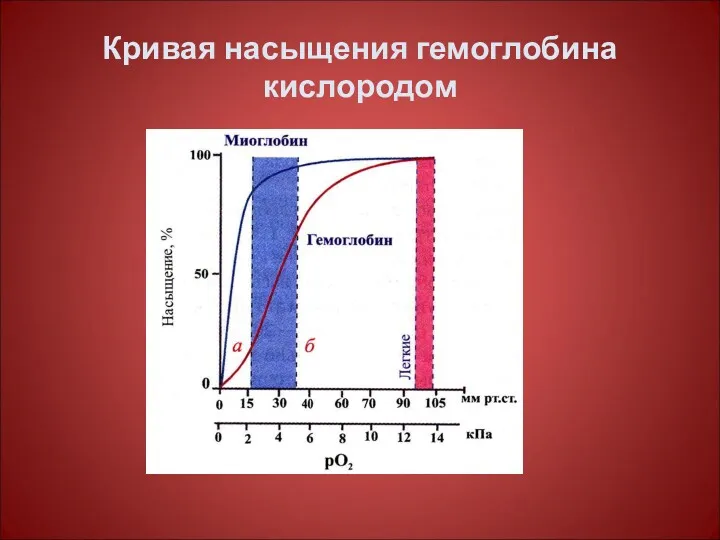
Кривая насыщения гемоглобина кислородом
Кривая насыщения гемоглобина кислородом

Виды гемоглобина человека
Гемоглобин А (НвА) - α 2β2 - основной тип
Виды гемоглобина человека
Гемоглобин А (НвА) - α 2β2 - основной тип
 Отдел Покрытосеменные Класс Двудольные
Отдел Покрытосеменные Класс Двудольные Один из методов решения генетических задач
Один из методов решения генетических задач Игра Кто это (презентация)
Игра Кто это (презентация) Тип Моллюски или мягкотелые
Тип Моллюски или мягкотелые Общие признаки животных
Общие признаки животных Морфологическое, систематическое и экологическое разнообразие насекомых
Морфологическое, систематическое и экологическое разнообразие насекомых Человек как результат биологической и социокультурной эволюции. Обществознание. 10 класс
Человек как результат биологической и социокультурной эволюции. Обществознание. 10 класс Тип моллюски. Курс зоологии
Тип моллюски. Курс зоологии Оксифотобактерии, или Цианобактерии
Оксифотобактерии, или Цианобактерии Сообщество, биоценоз, биогеоценоз , экосистема, биотоп
Сообщество, биоценоз, биогеоценоз , экосистема, биотоп Как устроен организм человека
Как устроен организм человека Группа ракообразные
Группа ракообразные Лабораторный практикум. Рост и развитие растений. (6 класс)
Лабораторный практикум. Рост и развитие растений. (6 класс) Психика и центральная нервная система. (Тема 6)
Психика и центральная нервная система. (Тема 6) Роль соли для организма человека
Роль соли для организма человека Что изучает генетика? Что такое наследственность? Каким способом передаются наследственные признаки?
Что изучает генетика? Что такое наследственность? Каким способом передаются наследственные признаки? Происхождение геоботаники
Происхождение геоботаники Всероссийская проверочная работа по биологии
Всероссийская проверочная работа по биологии Подготовка к ЕГЭ по биологии
Подготовка к ЕГЭ по биологии Система органов дыхания
Система органов дыхания Амфибии
Амфибии Открытый урок по биологии на тему Гуморальная регуляция
Открытый урок по биологии на тему Гуморальная регуляция Подцарство Одноклеточные, или Простейшие
Подцарство Одноклеточные, или Простейшие Презентация Класс Земноводные
Презентация Класс Земноводные Докази еволюції
Докази еволюції Разнообразие растений на Земле
Разнообразие растений на Земле Деревья леса
Деревья леса Класс Пресмыкающиеся (Рептилии)
Класс Пресмыкающиеся (Рептилии)