- Структурные и функциональные особенности поперечно-полосатых и гладких мышц

Содержание
- 2. Основная Литература Лекции, поскольку на них будут представлены основные результаты научных исследований сотрудников лаборатории «Структуры и
- 3. Основной учебник Основной в ряде вопросов
- 4. Второе издание учебника (первое вышло в 1997 году и трижды стереотипно тиражировано в 1998, 2000 и
- 5. Ещё один хороший учебник
- 6. Крупномасштабная структура Вселенной, как она выглядит в инфракрасных лучах с длиной волны 2,2 мкм — 1
- 7. Галактика Млечный Путь Млечный Путь – галактика, в которой находятся Земля, Солнечная система и все отдельные
- 8. Панорама Млечного Пути, сделанная в Долине Смерти, США, 2005 год Панорама южного неба, сделанная около обсерватории
- 9. Солнечная система Возраст 4,5682±0,0006 млрд лет
- 10. История формирования Земли Земля образовалась около 4,567 млрд лет назад путём аккреции из протопланетного диска, дискообразной
- 11. История формирования представлений о нашей планете и Солнечной системе Никола́й Копе́рник (19 февраля 1473, Торунь —
- 12. Джорда́но Бру́но (итал. Giordano Bruno; урождённый Филиппо Бруно, прозвище Бруно Ноланец; 1548, Нола близ Неаполя —
- 13. Памятник Джордано Бруно на площади Кампо деи Фиори в Риме, установленный на месте казни философа (скульптор
- 14. Галиле́о Галиле́й (итал. Galileo Galilei; 15 февраля 1564, Пиза — 8 января 1642, Арчетри) — итальянский
- 15. «Вследствие рассмотрения твоей вины и сознания твоего в ней присуждаем и объявляем тебя, Галилей, за всё
- 16. «И всё-таки она вертится» Общеизвестна легенда, по которой после показного отречения Галилей сказал: «И всё-таки она
- 17. Человек и его интерес к мышцам Впервые о наличии в человеческом теле мышц говорили еще в
- 18. Самым выдающимся ученым в разных областях медицины после Гиппократа стал римский анатом и физиолог Клавдий Гален
- 19. Рисунки Леонардо да Винчи (1452-1519 гг.)
- 20. Физиологию мышц исследовали некоторые ученые XVIII-XX веков: Луиджи Гальвани - открыл явление электрических импульсов в мышцах
- 21. Пионером исследования мышечного сокращения и, в частности, роли электрического поля в живом организме был профессор анатомии
- 22. Вольтов столб Во́льтов сто́лб — устройство, применявшееся на заре электротехники для получения электричества. В 1800 году
- 23. Физиология мышц исследовали и другие ученые XIX-XX веков: Н. Е. Введенский - описал и установил оптимум
- 24. Наука о мышцах – миология! Мышцам присуще свойство сократимости! А присуще ли это свойство немышечным тканям?
- 25. Свойство сократимости обнаруживается не только в животном мире, но и у ряда растений (росянка, захватывающая насекомых),
- 26. Клетка – элементарная структурная единица живой материи Клетка — структурно-функциональная элементарная единица строения и жизнедеятельности всех
- 27. Клеточная теория — одно из общепризнанных биологических обобщений, утверждающих единство принципа строения и развития мира растений,
- 28. Как выглядят клетки Клетки крови человека Одноклеточная водоросль Micrasterias radiata (дифференциальная интерференционно-контрастная микроскопия) Клетки эпителия щеки
- 29. Схематическое изображение животной клетки
- 30. Схема ультрамикроскопического строения клетки (по М. Р. Сапину, Г. Л. Билич, 1989): 1 — цитолемма (плазматическая
- 31. Клеточный центр, или центросома — немембранный органоид в клетках эукариот, состоит из двух центриолей и перицентриолярного
- 32. Я́дрышко — немембранный внутриядерный субкомпартмент, присущий всем без исключения эукариотическим организмам. Представляет собой комплекс белков и
- 33. Отличия животной и растительной клеток
- 34. Каждая клетка имеет мембрану Клеточная мембра́на (или цитолемма, или плазмалемма, или плазматическая мембрана) отделяет содержимое любой
- 35. фосфолипид Фосфолипиды – сложные липиды, сложные эфиры многоатомных спиртов и высших жирных кислот. Содержат остаток фосфорной
- 36. Изображение клеточной мембраны. Маленькие голубые и белые шарики соответствуют гидрофильным «головкам» фосфолипидов, а присоединённые к ним
- 37. MEMBRANE-ASSOCIATED PROTEIN COMPLEXES IN SKELETAL MUSCLE FIBERS & CONNECTIVE TISSUE Extrajunctional muscle membrane: Associated proteins Neuromuscular
- 38. Функции мембраны
- 39. Виды транспорта веществ через мембрану клетки Через мембрану должны проходить молекулы разных веществ. Они могут быть
- 40. Виды транспорта веществ через мембрану клетки Диффузия — пассивный транспорт, она происходит по градиенту электрохимического потенциала
- 41. В мембране по функциональному признаку различают следующие белки: 1. Структурные белки. 2. Рецепторы. 3. Ферменты. 4.
- 42. Стpуктуpные белки составляют остов или основу мембpаны. Остальные белки обеспечивают тpанспоpт веществ чеpез мембpану. Рецептоpы –
- 43. Ионные каналы подразделяются на: 1. Ионоселективные 2. Каналы "утечки" 3. Каналы "насосы" Ионоселективные каналы: 1. Осуществляют
- 44. Каналы "утечки": 1. Осуществляют пассивный тpанспоpт 2. Hе обладают селективностью 3. Hе имеют воpот (т.е. всегда
- 45. Ионные каналы обеспечивают два важных свойства мембраны: селективность и проводимость. ! Каналы "насосы" (Na-K; Ca насосы):
- 46. Мембранный потенциал Мембранный потенциал, также трансмембранный потенциал или напряжение мембраны, иногда потенциал Нернста — разница в
- 47. Электрический потенциал Электри́ческий потенциа́л — временна́я компонента четырёхмерного электромагнитного потенциала, называемый также иногда скалярным потенциалом (скалярным
- 48. В современной физике электромагни́тный потенциа́л обычно означает четырёхмерный потенциал электромагнитного поля, являющийся 4-вектором (1-формой). Именно в
- 49. В 1948 г А. Ходжкин, А. Хаксли и Б. Катц (английские электрофизиологи) получили Нобелевскую премию за
- 50. То, что внешняя мембрана заряжена положительно, а внутренняя отрицательно можно легко убедиться, если ввести внутрь клетки
- 51. Активный перенос ионов осуществляется в результате деятельности ионного калий-натриевого насоса («помпы»). Активный транспорт ионов натрия из
- 52. Пассивное движение ионов осуществляется по градиенту концентрации и не требует затрат энергии. В состоянии покоя клеточная
- 53. Итак, подытожим: Первый этап создания внутренней отрицательности мембраны: обмен Na+ на K+... Второй этап создания отрицательности:
- 54. Когда нервный импульс достигает окончания аксона, на деполяризованной пресинаптической мембране открываются потенциалзависимые Са2+ каналы. Вход Са2+
- 55. Потенциал действия. Потенциалом действия называют быстрое колебание мембранного потенциала, возникшее при возбуждении нервных и мышечных клеток.
- 56. Чувствительность постсинаптической мембраны регулируется активностью фермента – ацетилхолинэстеразы (АЦХ-Э), который гидролизует медиатор АЦХ на составные компоненты
- 57. Дальше на второй лекции рассказывать
- 58. Саркоплазматический ретикулум Мышечные клетки имеют специализированную, подобную гладкому эндоплазматическому ретикулуму, органеллу, называемую саркоплазматическим ретикулумом, которая захватывает
- 59. Саркоплазматический ретикулум мышц гомологичен эндоплазматическому ретикулуму других клеток. Он располагается вокруг каждой миофибриллы наподобие "рваного рукава",
- 60. Chittenden RH, Goodwin R J Physiol. 1891 Apr;12(1):34-41. Myosin-Peptone. . Gorter E1, Van Ormondt H Biochem
- 61. Влади́мир Алекса́ндрович Энгельга́рдт (1894—1984) — выдающийся советский биохимик, специалист в области молекулярной биологии. Академик Академии наук
- 62. А – схема расположения саркомера в миофибриллах и его строение. Толстые нити, состоящие в основном из
- 63. У человека более 600 скелетных мышц (около 40% массы тела). Скелетная мышечная ткань обеспечивает осознанные и
- 64. Структурно-функциональная единица скелетной мышцы - симпласт - скелетное мышечное волокно, имеет форму протяжённого цилиндра с заострёнными
- 65. Рыхлая волокнистая соединительная ткань между отдельными мышечными волокнами (эндомизий) содержит кровеносные и лимфатические сосуды, нервные волокна.
- 66. Фрагмент скелетного мышечного волокна. Цистерны саркоплазматического ретикулума окружают каждую миофибриллу. Т-трубочки подходят к миофибриллам на уровне
- 67. ИННЕРВАЦИЯ СКЕЛЕТНЫХ МЫШЦ В скелетных мышцах различают экстрафузальные и интрафузальные мышечные волокна. • Экстрафузальные мышечные волокна,
- 68. Интрафузальные мышчные волокна специализированы для детектирования натяжения. Они получают спиральные окончания чувствительных нервных волокон групп 1а
- 69. Как подсказывает название "рецептор растяжения", мышечное веретено реагирует на растяжение мышцы. Рис. демонстрирует изменение активности афферентного
- 70. Схема строения мышечного волокна Продольный срез поперечно-полосатой мышцы. Саркомеры. Поперечный срез поперечно-полосатой мышцы. Скелетные мышцы Скелетная
- 71. Медленные фазические волокна окислительного типа. Волокна этого типа характеризуются большим содержанием белка миоглобина (мышечный гемоглобин), который
- 72. Тонические волокна. В отличие от предыдущих мышечных волокон в тонических волокнах двигательный аксон образует множество синаптических
- 73. Мышечные волокна не являются функциональной единицей скелетной мускулатуры. Эту роль выполняет нейромоторная, или двигательная, единица, которая
- 74. КАРДИОМИОЦИТЫ Клетки расположены между элементами рыхлой волокнистой соединительной ткани, содержащей многочисленные кровеносные капилляры бассейна венечных сосудов
- 75. Рабочие кардиомиоциты - морфо-функциональные единицы сердечной мышечной ткани, имеют цилиндрическую ветвящуюся форму диаметром около 15 мкм.
- 76. Предсердные и желудочковые кардиомиоциты. Предсердные и желудочковые кардиомиоциты относятся к разным популяциям рабочих кардиомиоцитов. Предсердные кардиомиоциты
- 77. Водители ритма (пейсмейкерные клетки, пейсмейкеры,) - совокупность специализированных кардиомиоцитов в виде тонких волокон, окружённых рыхлой соединительной
- 78. ДВИГАТЕЛЬНАЯ ИННЕРВАЦИЯ СЕРДЦА На деятельность сердца - сложной авторегуляторной и регулируемой системы - оказывает модулирующее влияние
- 79. Морфо-функциональная единица гладкой мышечной ткани – ГладкоМышечная Клетка (ГМК). Заострёнными концами ГМК вклиниваются между соседними клетками
- 81. Центральное положение в ГМК занимает крупное ядро. У полюсов ядра находятся митохондрии, эндоплазматический ретикулум и комплекс
- 82. Сократительный аппарат гладкомышечной клетки. Плотные тельца содержат α-актинин, это аналоги Z-линий поперечно-полосатой мышцы. В саркоплазме они
- 84. ИННЕРВАЦИЯ ГЛАДКОЙ МЫШЦЫ Симпатические (адренергические) и отчасти парасимпатические (холинергические) нервные волокна иннервируют ГМК. Нейромедиаторы диффундируют из
- 85. Следующая лекция Электромеханическое сопряжение
- 86. Различают следующие типы и виды транспорта веществ через плазматическую мембрану. I. Пассивный транспорт – без затрат
- 87. Функции и свойства скелетных мышц Скелетная мускулатура является составной частью опорно-двигательного аппарата человека. При этом мышцы
- 88. Осмос – основной способ поступления воды в клетки и организм. Основной силой, обусловливающей перемещение воды через
- 89. Путем простой диффузии в клетку проникают половые гормоны и гормон тироксин, аминокислоты с гидрофобными радикалами, моносахариды.
- 90. Путем облегченной диффузии в клетку проникает глюкоза и некоторые аминокислоты. Особенностью процесса облегченной диффузии является то,
- 91. Сердечная мышца В клетках сердечной мышцы млекопитающих имеется одно ядро в центре клетки, около него расположены
- 92. По структуре сократительных элементов сердечная мышца сходна с пореречно полосатыми мышцами, хотя по своим физиологическим свойствам
- 93. Структурным элементом поперечно полосатых мышечных тканей служит уже не клетка, а волокно, которое легко можно изолировать
- 94. Электромеханическое сопряжение скелет мышцы Электромеханическое сопряжение - это последовательность процессов, в результате которых потенциал действия плазматической
- 95. Электромеханическое сопряжение глад мышцы
- 96. Тайтин и миозин in vitro Выпрямленные молекулы тайтина Агрегаты тайтина Нити миозина Шкала 200 нм Нити
- 97. Функции тайтина W. Linke, 2000 1. Функция эластичного саркомерного цитоскелета - Тайтин вносит вклад в пассивное
- 98. Благодарю за внимание!
- 99. Белки мембраны клеток представлены, в основном, гликопротеинами. Различают интегральные белки, проникающие через всю толщу мембраны и
- 101. Скачать презентацию

Основная Литература
Лекции, поскольку на них будут представлены основные результаты научных исследований
Основная Литература
Лекции, поскольку на них будут представлены основные результаты научных исследований

Основной учебник
Основной в ряде вопросов
Основной учебник
Основной в ряде вопросов

Второе издание учебника (первое вышло в 1997 году и трижды стереотипно
Второе издание учебника (первое вышло в 1997 году и трижды стереотипно

Ещё один хороший учебник
Ещё один хороший учебник
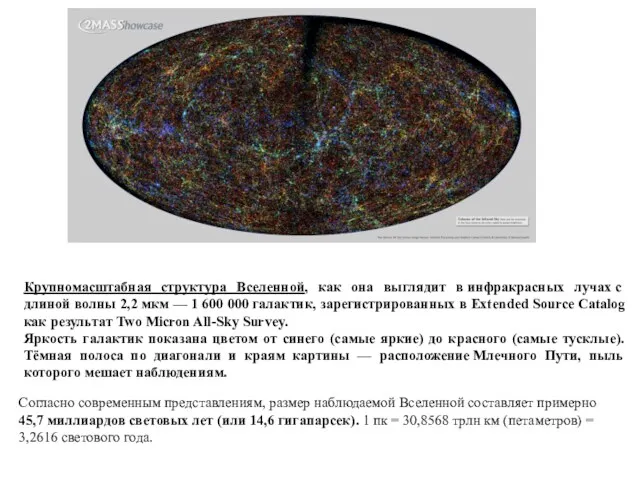
Крупномасштабная структура Вселенной, как она выглядит в инфракрасных лучах с длиной волны 2,2
Крупномасштабная структура Вселенной, как она выглядит в инфракрасных лучах с длиной волны 2,2
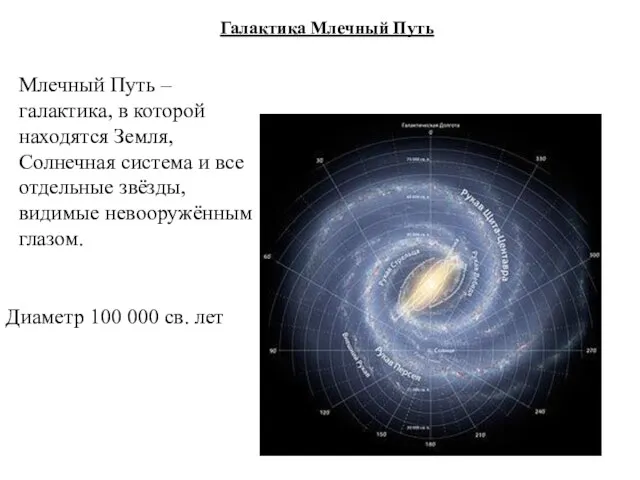
Галактика Млечный Путь
Млечный Путь – галактика, в которой находятся Земля, Солнечная
Галактика Млечный Путь
Млечный Путь – галактика, в которой находятся Земля, Солнечная

Панорама Млечного Пути, сделанная в Долине Смерти, США, 2005 год
Панорама южного
Панорама Млечного Пути, сделанная в Долине Смерти, США, 2005 год
Панорама южного
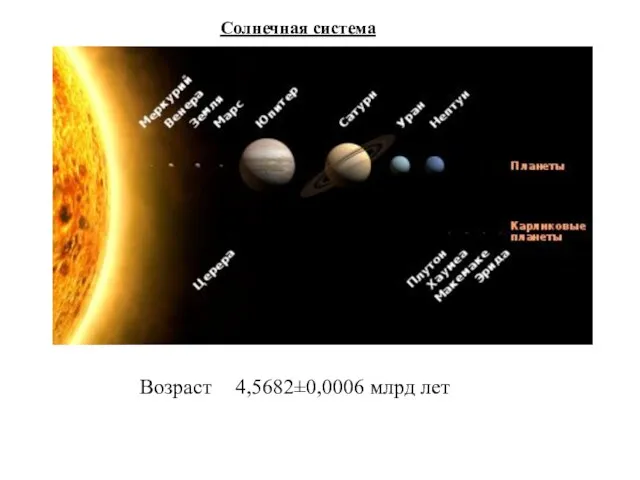
Солнечная система
Возраст 4,5682±0,0006 млрд лет
Солнечная система
Возраст 4,5682±0,0006 млрд лет
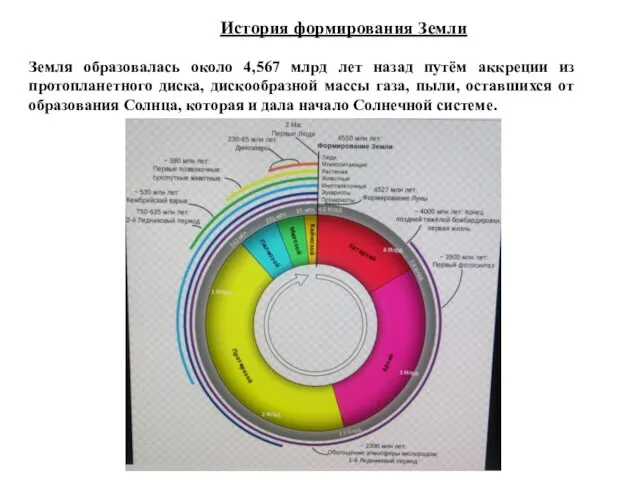
История формирования Земли
Земля образовалась около 4,567 млрд лет назад путём
История формирования Земли
Земля образовалась около 4,567 млрд лет назад путём

История формирования представлений о нашей планете и Солнечной системе
Никола́й Копе́рник
История формирования представлений о нашей планете и Солнечной системе
Никола́й Копе́рник

Джорда́но Бру́но (итал. Giordano Bruno; урождённый Филиппо Бруно, прозвище Бруно Ноланец;
Джорда́но Бру́но (итал. Giordano Bruno; урождённый Филиппо Бруно, прозвище Бруно Ноланец;

Памятник Джордано Бруно на площади Кампо деи Фиори в Риме, установленный
Памятник Джордано Бруно на площади Кампо деи Фиори в Риме, установленный

Галиле́о Галиле́й (итал. Galileo Galilei; 15 февраля 1564, Пиза — 8
Галиле́о Галиле́й (итал. Galileo Galilei; 15 февраля 1564, Пиза — 8

«Вследствие рассмотрения твоей вины и сознания твоего в ней присуждаем и
«Вследствие рассмотрения твоей вины и сознания твоего в ней присуждаем и

«И всё-таки она вертится»
Общеизвестна легенда, по которой после показного отречения Галилей
«И всё-таки она вертится»
Общеизвестна легенда, по которой после показного отречения Галилей

Человек и его интерес к мышцам
Впервые о наличии в человеческом теле
Человек и его интерес к мышцам
Впервые о наличии в человеческом теле

Самым выдающимся ученым в разных областях медицины после Гиппократа стал римский
Самым выдающимся ученым в разных областях медицины после Гиппократа стал римский
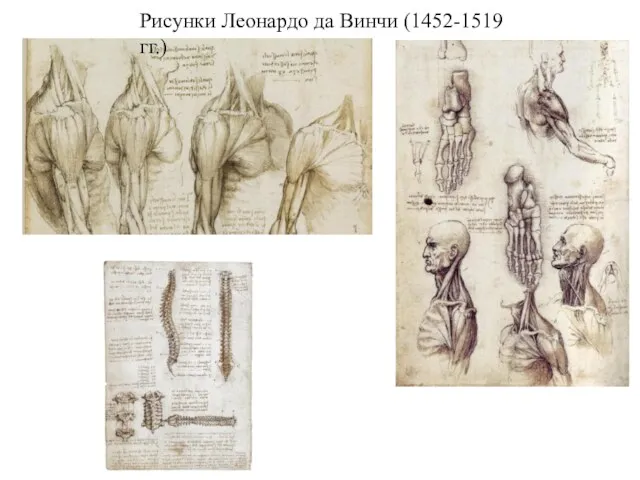
Рисунки Леонардо да Винчи (1452-1519 гг.)
Рисунки Леонардо да Винчи (1452-1519 гг.)

Физиологию мышц исследовали некоторые ученые XVIII-XX веков:
Луиджи Гальвани - открыл
Физиологию мышц исследовали некоторые ученые XVIII-XX веков:
Луиджи Гальвани - открыл

Пионером исследования мышечного сокращения и, в частности, роли электрического поля в
Пионером исследования мышечного сокращения и, в частности, роли электрического поля в

Вольтов столб
Во́льтов сто́лб — устройство, применявшееся на заре электротехники для получения
Вольтов столб
Во́льтов сто́лб — устройство, применявшееся на заре электротехники для получения

Физиология мышц исследовали и другие ученые XIX-XX веков:
Н. Е. Введенский
Физиология мышц исследовали и другие ученые XIX-XX веков:
Н. Е. Введенский

Наука о мышцах – миология!
Мышцам присуще свойство сократимости!
А присуще
Наука о мышцах – миология!
Мышцам присуще свойство сократимости!
А присуще

Свойство сократимости обнаруживается не только в животном мире, но и у
Свойство сократимости обнаруживается не только в животном мире, но и у

Клетка – элементарная структурная единица живой материи
Клетка — структурно-функциональная элементарная единица
Клетка – элементарная структурная единица живой материи
Клетка — структурно-функциональная элементарная единица

Клеточная теория — одно из общепризнанных биологических обобщений, утверждающих единство принципа
Клеточная теория — одно из общепризнанных биологических обобщений, утверждающих единство принципа
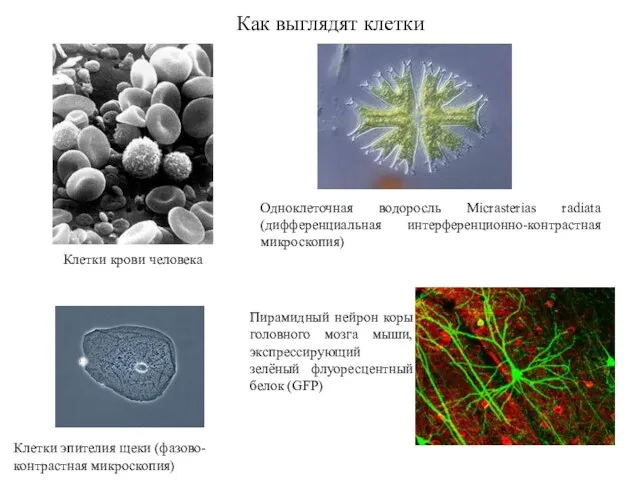
Как выглядят клетки
Клетки крови человека
Одноклеточная водоросль Micrasterias radiata (дифференциальная интерференционно-контрастная
Как выглядят клетки
Клетки крови человека
Одноклеточная водоросль Micrasterias radiata (дифференциальная интерференционно-контрастная
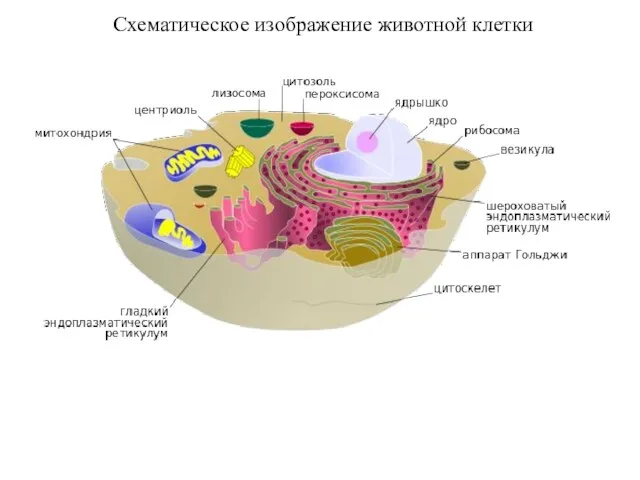
Схематическое изображение животной клетки
Схематическое изображение животной клетки
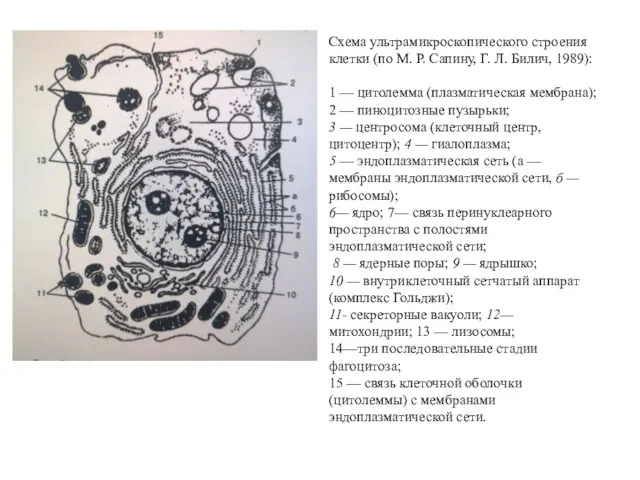
Схема ультрамикроскопического строения клетки (по М. Р. Сапину, Г. Л. Билич,
Схема ультрамикроскопического строения клетки (по М. Р. Сапину, Г. Л. Билич,

Клеточный центр, или центросома — немембранный органоид в клетках эукариот, состоит
Клеточный центр, или центросома — немембранный органоид в клетках эукариот, состоит

Я́дрышко — немембранный внутриядерный субкомпартмент, присущий всем без исключения эукариотическим организмам.
Я́дрышко — немембранный внутриядерный субкомпартмент, присущий всем без исключения эукариотическим организмам.
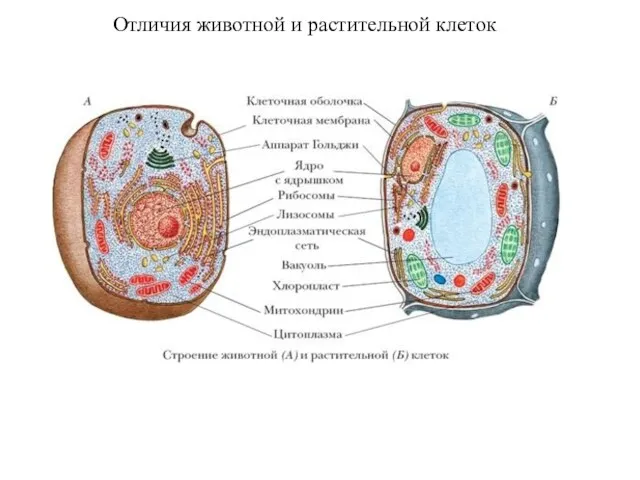
Отличия животной и растительной клеток
Отличия животной и растительной клеток
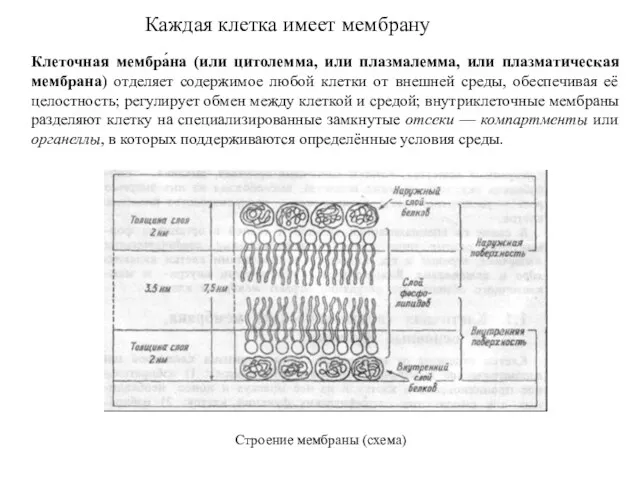
Каждая клетка имеет мембрану
Клеточная мембра́на (или цитолемма, или плазмалемма, или плазматическая
Каждая клетка имеет мембрану
Клеточная мембра́на (или цитолемма, или плазмалемма, или плазматическая
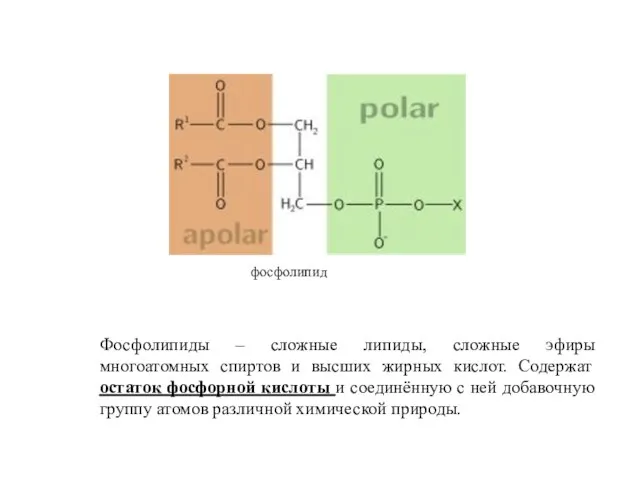
фосфолипид
Фосфолипиды – сложные липиды, сложные эфиры многоатомных спиртов и высших жирных
фосфолипид
Фосфолипиды – сложные липиды, сложные эфиры многоатомных спиртов и высших жирных
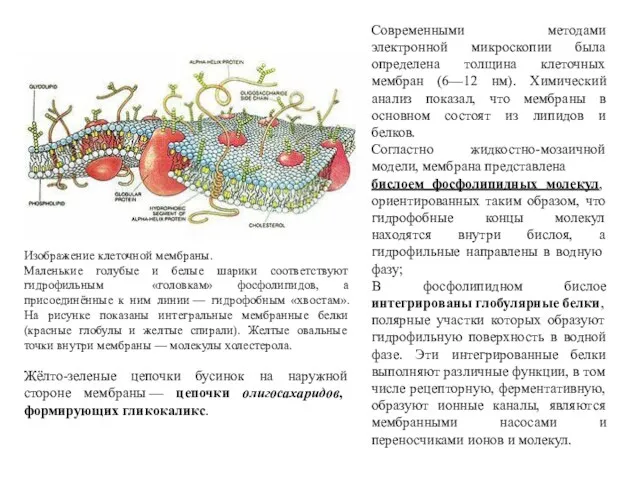
Изображение клеточной мембраны.
Маленькие голубые и белые шарики соответствуют гидрофильным «головкам»
Изображение клеточной мембраны.
Маленькие голубые и белые шарики соответствуют гидрофильным «головкам»
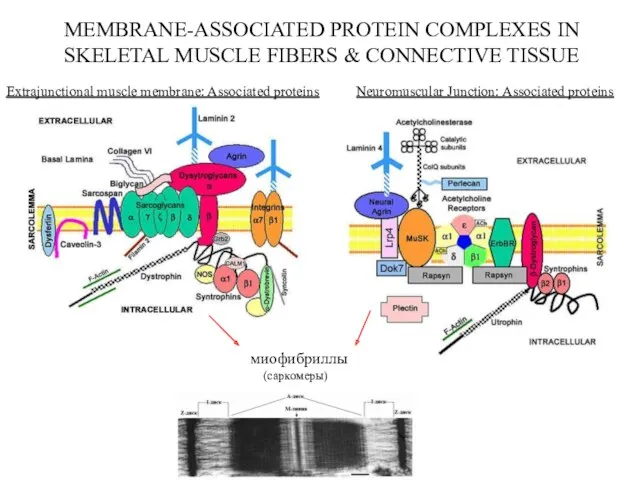
MEMBRANE-ASSOCIATED PROTEIN COMPLEXES IN
SKELETAL MUSCLE FIBERS & CONNECTIVE TISSUE
Extrajunctional muscle
MEMBRANE-ASSOCIATED PROTEIN COMPLEXES IN
SKELETAL MUSCLE FIBERS & CONNECTIVE TISSUE
Extrajunctional muscle

Функции мембраны
Функции мембраны

Виды транспорта веществ через мембрану клетки
Через мембрану должны проходить молекулы разных
Виды транспорта веществ через мембрану клетки
Через мембрану должны проходить молекулы разных

Виды транспорта веществ через мембрану клетки
Диффузия — пассивный транспорт, она происходит
Виды транспорта веществ через мембрану клетки
Диффузия — пассивный транспорт, она происходит

В мембране по функциональному признаку различают следующие белки:
1. Структурные белки.
2. Рецепторы.
3.
В мембране по функциональному признаку различают следующие белки:
1. Структурные белки.
2. Рецепторы.
3.

Стpуктуpные белки составляют остов или основу мембpаны.
Остальные белки обеспечивают тpанспоpт веществ
Стpуктуpные белки составляют остов или основу мембpаны.
Остальные белки обеспечивают тpанспоpт веществ
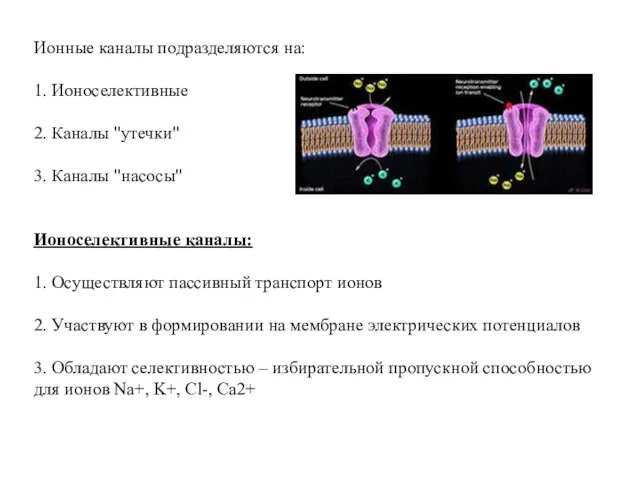
Ионные каналы подразделяются на:
1. Ионоселективные
2. Каналы "утечки"
3. Каналы "насосы"
Ионоселективные каналы:
1. Осуществляют
Ионные каналы подразделяются на:
1. Ионоселективные
2. Каналы "утечки"
3. Каналы "насосы"
Ионоселективные каналы:
1. Осуществляют

Каналы "утечки":
1. Осуществляют пассивный тpанспоpт
2. Hе обладают селективностью
3. Hе имеют воpот
Каналы "утечки":
1. Осуществляют пассивный тpанспоpт
2. Hе обладают селективностью
3. Hе имеют воpот

Ионные каналы обеспечивают два важных свойства мембраны: селективность и проводимость.
!
Каналы "насосы"
Ионные каналы обеспечивают два важных свойства мембраны: селективность и проводимость.
!
Каналы "насосы"

Мембранный потенциал
Мембранный потенциал, также трансмембранный потенциал или напряжение мембраны, иногда
Мембранный потенциал
Мембранный потенциал, также трансмембранный потенциал или напряжение мембраны, иногда

Электрический потенциал
Электри́ческий потенциа́л — временна́я компонента четырёхмерного электромагнитного потенциала, называемый
Электрический потенциал
Электри́ческий потенциа́л — временна́я компонента четырёхмерного электромагнитного потенциала, называемый

В современной физике электромагни́тный потенциа́л обычно означает четырёхмерный потенциал электромагнитного поля,
В современной физике электромагни́тный потенциа́л обычно означает четырёхмерный потенциал электромагнитного поля,

В 1948 г А. Ходжкин, А. Хаксли и Б. Катц (английские
В 1948 г А. Ходжкин, А. Хаксли и Б. Катц (английские

То, что внешняя мембрана заряжена положительно, а внутренняя отрицательно можно легко
То, что внешняя мембрана заряжена положительно, а внутренняя отрицательно можно легко

Активный перенос ионов осуществляется в результате деятельности ионного калий-натриевого насоса («помпы»).
Активный перенос ионов осуществляется в результате деятельности ионного калий-натриевого насоса («помпы»).

Пассивное движение ионов осуществляется по градиенту концентрации и не требует затрат

Итак, подытожим:
Первый этап создания внутренней отрицательности мембраны: обмен Na+ на
Итак, подытожим:
Первый этап создания внутренней отрицательности мембраны: обмен Na+ на

Когда нервный импульс достигает окончания аксона, на деполяризованной пресинаптической мембране открываются
Когда нервный импульс достигает окончания аксона, на деполяризованной пресинаптической мембране открываются

Потенциал действия. Потенциалом действия называют быстрое колебание мембранного потенциала, возникшее при
Потенциал действия. Потенциалом действия называют быстрое колебание мембранного потенциала, возникшее при
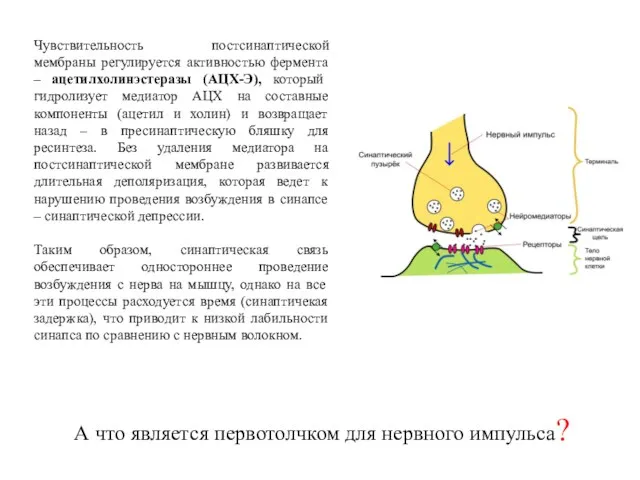
Чувствительность постсинаптической мембраны регулируется активностью фермента – ацетилхолинэстеразы (АЦХ-Э), который гидролизует
Чувствительность постсинаптической мембраны регулируется активностью фермента – ацетилхолинэстеразы (АЦХ-Э), который гидролизует

Дальше на второй лекции рассказывать
Дальше на второй лекции рассказывать
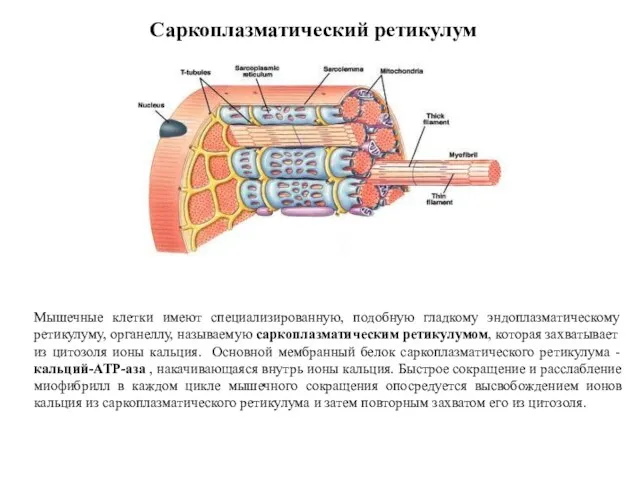
Саркоплазматический ретикулум
Мышечные клетки имеют специализированную, подобную гладкому эндоплазматическому ретикулуму, органеллу, называемую
Саркоплазматический ретикулум
Мышечные клетки имеют специализированную, подобную гладкому эндоплазматическому ретикулуму, органеллу, называемую

Саркоплазматический ретикулум мышц гомологичен эндоплазматическому ретикулуму других клеток. Он располагается вокруг

Chittenden RH, Goodwin R J Physiol. 1891 Apr;12(1):34-41.
Myosin-Peptone.
.
Gorter E1, Van Ormondt
Chittenden RH, Goodwin R J Physiol. 1891 Apr;12(1):34-41.
Myosin-Peptone.
.
Gorter E1, Van Ormondt

Влади́мир Алекса́ндрович Энгельга́рдт (1894—1984) — выдающийся советский биохимик, специалист в области
Влади́мир Алекса́ндрович Энгельга́рдт (1894—1984) — выдающийся советский биохимик, специалист в области
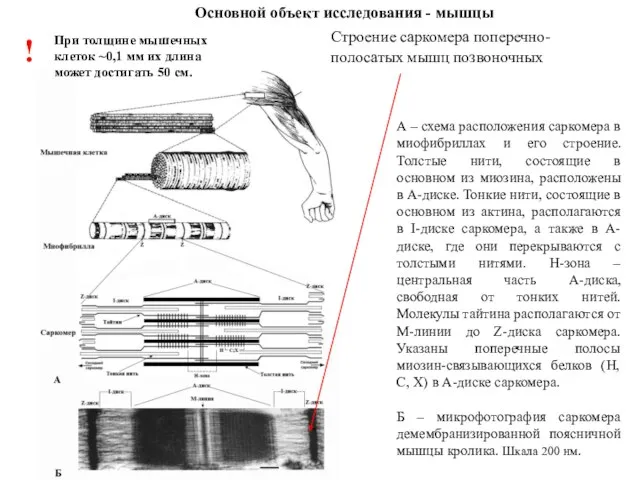
А – схема расположения саркомера в миофибриллах и его строение. Толстые

У человека более 600 скелетных мышц (около 40% массы тела). Скелетная
У человека более 600 скелетных мышц (около 40% массы тела). Скелетная
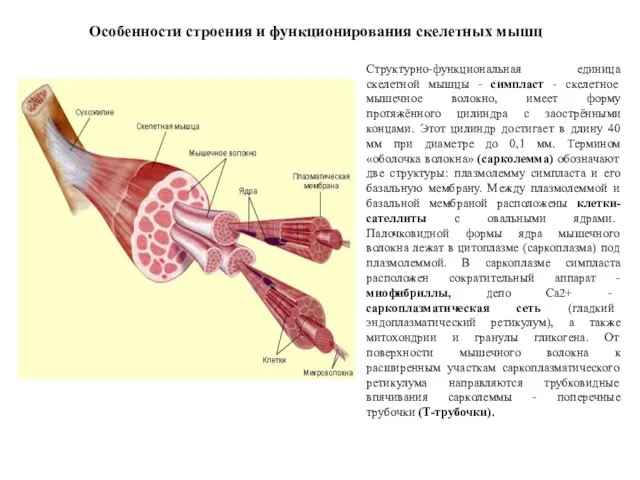
Структурно-функциональная единица скелетной мышцы - симпласт - скелетное мышечное волокно, имеет
Структурно-функциональная единица скелетной мышцы - симпласт - скелетное мышечное волокно, имеет
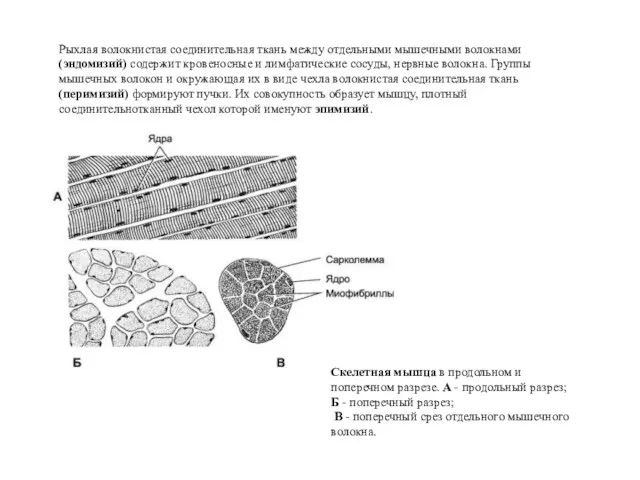
Рыхлая волокнистая соединительная ткань между отдельными мышечными волокнами (эндомизий) содержит кровеносные
Рыхлая волокнистая соединительная ткань между отдельными мышечными волокнами (эндомизий) содержит кровеносные
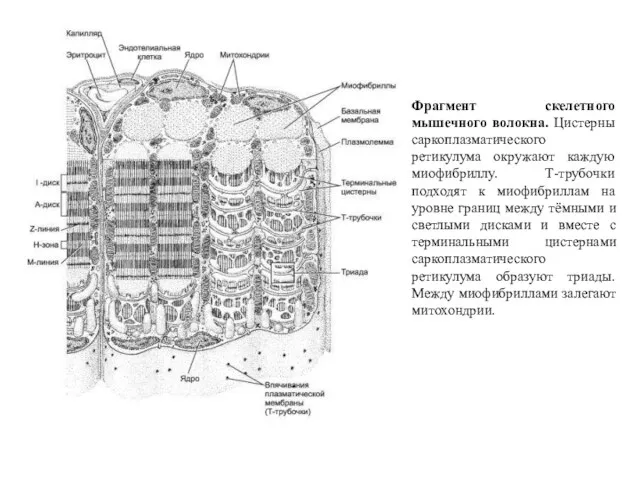
Фрагмент скелетного мышечного волокна. Цистерны саркоплазматического ретикулума окружают каждую миофибриллу. Т-трубочки
Фрагмент скелетного мышечного волокна. Цистерны саркоплазматического ретикулума окружают каждую миофибриллу. Т-трубочки

ИННЕРВАЦИЯ СКЕЛЕТНЫХ МЫШЦ
В скелетных мышцах различают экстрафузальные и интрафузальные мышечные волокна.
•
ИННЕРВАЦИЯ СКЕЛЕТНЫХ МЫШЦ
В скелетных мышцах различают экстрафузальные и интрафузальные мышечные волокна.
•

Интрафузальные мышчные волокна специализированы для детектирования натяжения. Они получают спиральные окончания
Интрафузальные мышчные волокна специализированы для детектирования натяжения. Они получают спиральные окончания

Как подсказывает название "рецептор растяжения", мышечное веретено реагирует на растяжение мышцы.
Как подсказывает название "рецептор растяжения", мышечное веретено реагирует на растяжение мышцы.

Схема строения мышечного
волокна
Продольный срез поперечно-полосатой
мышцы. Саркомеры.
Поперечный срез поперечно-полосатой мышцы.
Скелетные
Схема строения мышечного
волокна
Продольный срез поперечно-полосатой
мышцы. Саркомеры.
Поперечный срез поперечно-полосатой мышцы.
Скелетные

Медленные фазические волокна окислительного типа. Волокна этого типа характеризуются большим содержанием
Медленные фазические волокна окислительного типа. Волокна этого типа характеризуются большим содержанием

Тонические волокна. В отличие от предыдущих мышечных волокон в тонических волокнах
Тонические волокна. В отличие от предыдущих мышечных волокон в тонических волокнах
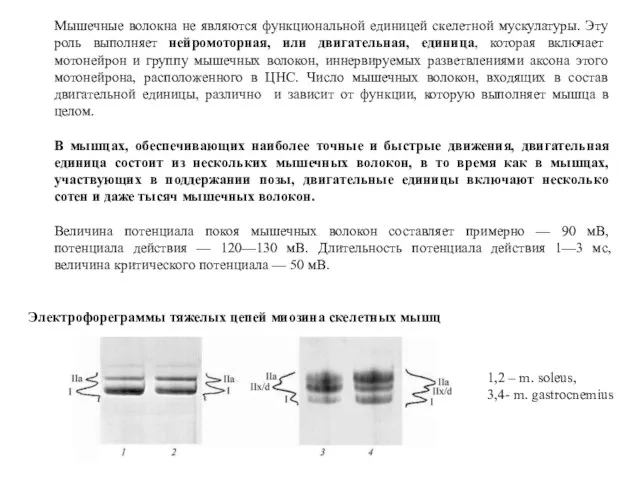
Мышечные волокна не являются функциональной единицей скелетной мускулатуры. Эту роль выполняет
Мышечные волокна не являются функциональной единицей скелетной мускулатуры. Эту роль выполняет
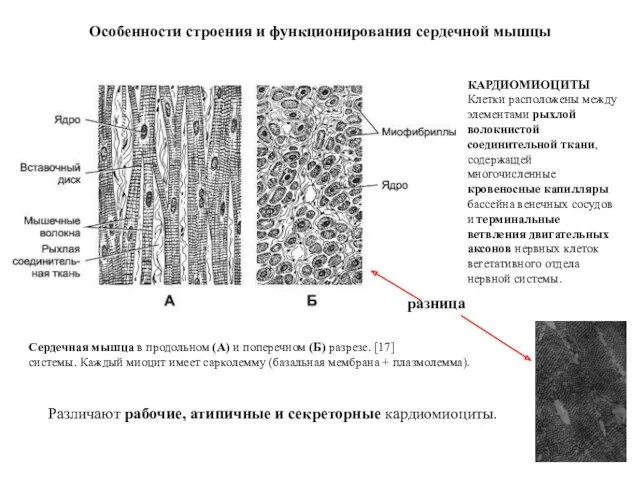
КАРДИОМИОЦИТЫ
Клетки расположены между элементами рыхлой волокнистой соединительной ткани, содержащей многочисленные кровеносные
КАРДИОМИОЦИТЫ
Клетки расположены между элементами рыхлой волокнистой соединительной ткани, содержащей многочисленные кровеносные
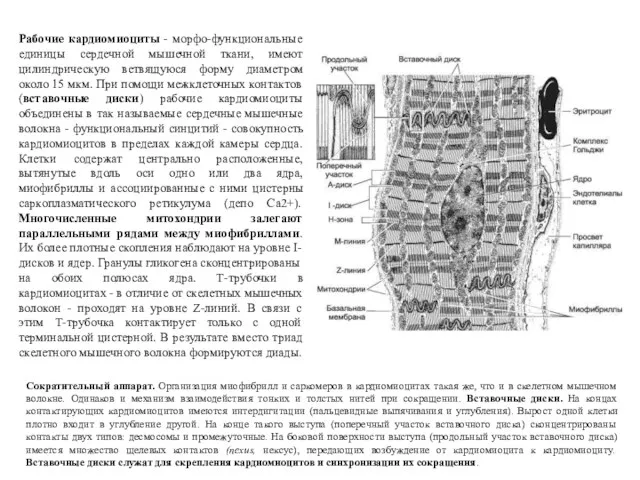
Рабочие кардиомиоциты - морфо-функциональные единицы сердечной мышечной ткани, имеют цилиндрическую ветвящуюся
Рабочие кардиомиоциты - морфо-функциональные единицы сердечной мышечной ткани, имеют цилиндрическую ветвящуюся

Предсердные и желудочковые кардиомиоциты. Предсердные и желудочковые кардиомиоциты относятся к разным
Предсердные и желудочковые кардиомиоциты. Предсердные и желудочковые кардиомиоциты относятся к разным
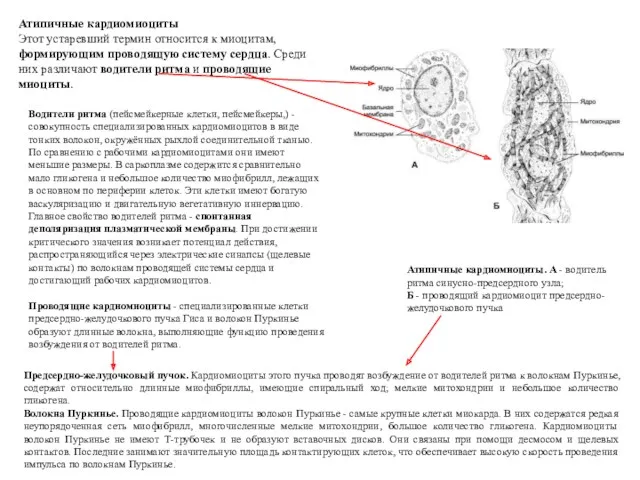
Водители ритма (пейсмейкерные клетки, пейсмейкеры,) - совокупность специализированных кардиомиоцитов в виде
Водители ритма (пейсмейкерные клетки, пейсмейкеры,) - совокупность специализированных кардиомиоцитов в виде

ДВИГАТЕЛЬНАЯ ИННЕРВАЦИЯ СЕРДЦА
На деятельность сердца - сложной авторегуляторной и регулируемой системы
ДВИГАТЕЛЬНАЯ ИННЕРВАЦИЯ СЕРДЦА
На деятельность сердца - сложной авторегуляторной и регулируемой системы

Морфо-функциональная единица гладкой мышечной ткани – ГладкоМышечная Клетка (ГМК). Заострёнными концами
Морфо-функциональная единица гладкой мышечной ткани – ГладкоМышечная Клетка (ГМК). Заострёнными концами

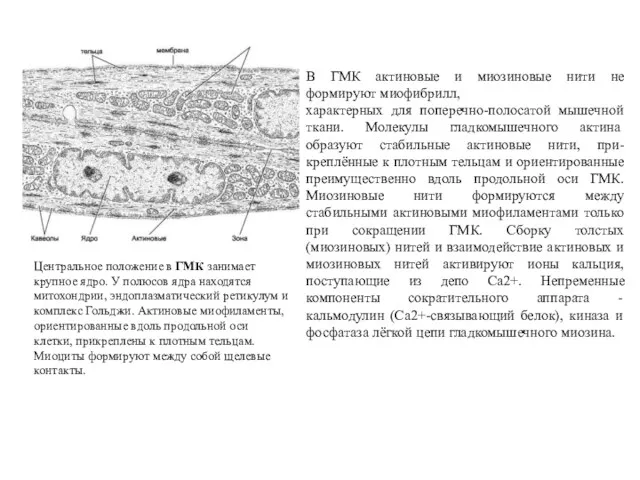
Центральное положение в ГМК занимает крупное ядро. У полюсов ядра находятся
Центральное положение в ГМК занимает крупное ядро. У полюсов ядра находятся
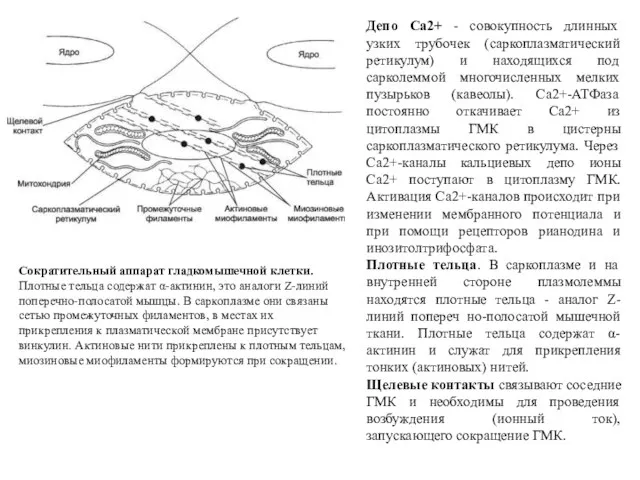
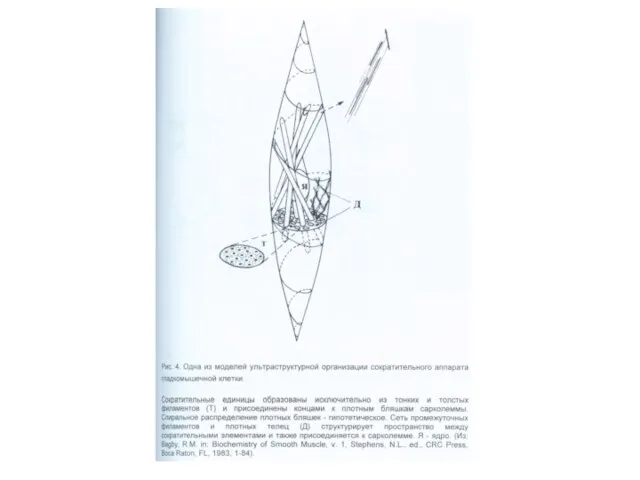
Сократительный аппарат гладкомышечной клетки. Плотные тельца содержат α-актинин, это аналоги Z-линий
Сократительный аппарат гладкомышечной клетки. Плотные тельца содержат α-актинин, это аналоги Z-линий

ИННЕРВАЦИЯ ГЛАДКОЙ МЫШЦЫ
Симпатические (адренергические) и отчасти парасимпатические (холинергические) нервные волокна иннервируют
ИННЕРВАЦИЯ ГЛАДКОЙ МЫШЦЫ
Симпатические (адренергические) и отчасти парасимпатические (холинергические) нервные волокна иннервируют

Следующая лекция
Электромеханическое сопряжение
Следующая лекция
Электромеханическое сопряжение

Различают следующие типы и виды транспорта веществ через плазматическую мембрану.
I. Пассивный
Различают следующие типы и виды транспорта веществ через плазматическую мембрану. I. Пассивный

Функции и свойства скелетных мышц
Скелетная мускулатура является составной частью опорно-двигательного аппарата
Функции и свойства скелетных мышц
Скелетная мускулатура является составной частью опорно-двигательного аппарата

Осмос – основной способ поступления воды в клетки и организм. Основной
Осмос – основной способ поступления воды в клетки и организм. Основной

Путем простой диффузии в клетку проникают половые гормоны и гормон тироксин,
Путем простой диффузии в клетку проникают половые гормоны и гормон тироксин,

Путем облегченной диффузии в клетку проникает глюкоза и некоторые аминокислоты. Особенностью
Путем облегченной диффузии в клетку проникает глюкоза и некоторые аминокислоты. Особенностью
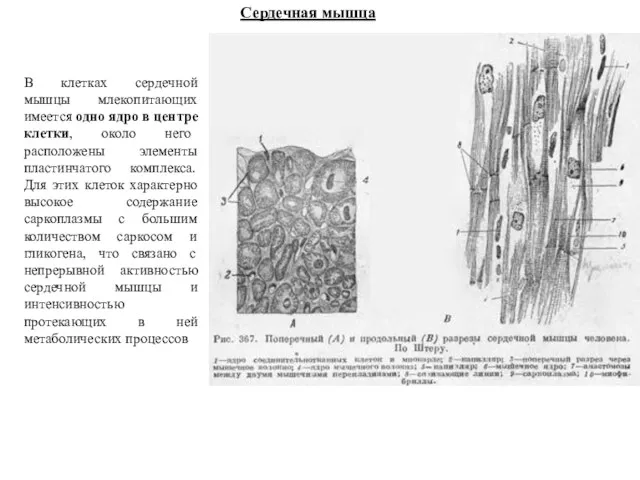
Сердечная мышца
В клетках сердечной мышцы млекопитающих имеется одно ядро в центре
Сердечная мышца
В клетках сердечной мышцы млекопитающих имеется одно ядро в центре

По структуре сократительных элементов сердечная мышца сходна с пореречно полосатыми мышцами,
По структуре сократительных элементов сердечная мышца сходна с пореречно полосатыми мышцами,

Структурным элементом поперечно полосатых мышечных тканей служит уже не клетка, а
Структурным элементом поперечно полосатых мышечных тканей служит уже не клетка, а

Электромеханическое сопряжение скелет мышцы
Электромеханическое сопряжение - это последовательность процессов, в
Электромеханическое сопряжение скелет мышцы
Электромеханическое сопряжение - это последовательность процессов, в

Электромеханическое сопряжение глад мышцы
Электромеханическое сопряжение глад мышцы

Тайтин и миозин in vitro
Выпрямленные молекулы тайтина
Агрегаты тайтина
Нити миозина
Шкала 200 нм
Тайтин и миозин in vitro
Выпрямленные молекулы тайтина
Агрегаты тайтина
Нити миозина
Шкала 200 нм
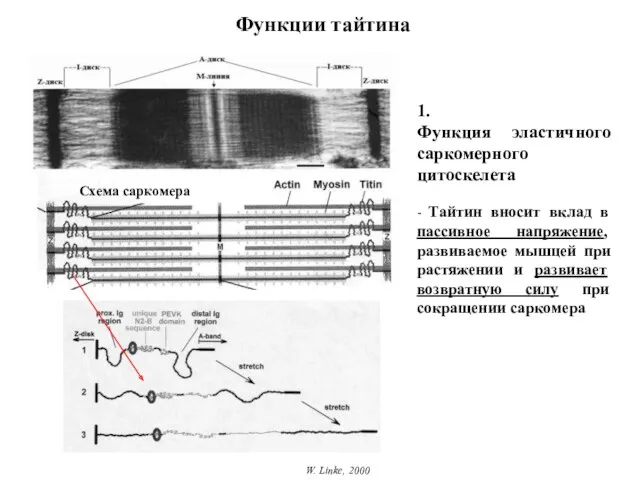
Функции тайтина
W. Linke, 2000
1.
Функция эластичного саркомерного цитоскелета
- Тайтин вносит вклад
Функции тайтина
W. Linke, 2000
1.
Функция эластичного саркомерного цитоскелета
- Тайтин вносит вклад

Благодарю
за
внимание!
Благодарю
за
внимание!

Белки мембраны клеток представлены, в основном, гликопротеинами.
Различают интегральные белки, проникающие через
Белки мембраны клеток представлены, в основном, гликопротеинами.
Различают интегральные белки, проникающие через
 Помощь бездомным животным. Проект
Помощь бездомным животным. Проект Біологія - наука про живу природу
Біологія - наука про живу природу Ротовая полость
Ротовая полость Life cycle of mosses and ferns
Life cycle of mosses and ferns Отцы - основатели биоинформатики
Отцы - основатели биоинформатики Генетика, законы Менделя
Генетика, законы Менделя Комнатные растения в интерьере квартиры
Комнатные растения в интерьере квартиры Индивидуальное развитие организма. Онтогенез
Индивидуальное развитие организма. Онтогенез Систематика покрытосеменных растений
Систематика покрытосеменных растений Мир вокруг нас. Почему звенит звонок
Мир вокруг нас. Почему звенит звонок Цікаві факти про метеликів: швидше, далі, яскравіше
Цікаві факти про метеликів: швидше, далі, яскравіше История развития анатомии, физиологии и медицины
История развития анатомии, физиологии и медицины Протерозойська ера
Протерозойська ера Коронавирусная инфекция vs Инфекционный перитонит кошек - FIP
Коронавирусная инфекция vs Инфекционный перитонит кошек - FIP Человек как фактор развития природы
Человек как фактор развития природы Функциональная анатомия вегетативной нервной системы
Функциональная анатомия вегетативной нервной системы Lancelet - its importance in evolution
Lancelet - its importance in evolution Үсемлек белән тукланучы һәм ерткыч хайваннар
Үсемлек белән тукланучы һәм ерткыч хайваннар Красная книга по Красноярскому краю
Красная книга по Красноярскому краю Топография сердца
Топография сердца Лишайники. Строение и жизнедеятельность лишайников
Лишайники. Строение и жизнедеятельность лишайников Гігієна транспотування тварин
Гігієна транспотування тварин Углеводы, липиды. Тема 7
Углеводы, липиды. Тема 7 Органы чувств. Зрительный анализатор
Органы чувств. Зрительный анализатор блок_4_Зоо_Простейшие
блок_4_Зоо_Простейшие Немного про акул
Немного про акул Mutation
Mutation Последствия близкородственных браков
Последствия близкородственных браков