- Структурные компоненты нуклеиновых кислот. Уровни организации ДНК и РНК

Содержание
- 2. В каждом живом организме присутствуют 2 типа нуклеиновых кислот: рибонуклеиновая кислота (РНК) и дезоксирибонуклеиновая кислота (ДНК).
- 3. История открытия 60 – е года 19 в. – швейцарский ученый Фридрих Мишер выделил из ядер
- 4. Локализация в клетке Основная часть ДНК находится в ядре клетки – в составе хроматина. 0, 25
- 5. Структурная организация НК – полимеры, мономерами которых являются нуклеотиды и выполняющие в клетке функции хранения, передачи
- 6. Строение нуклеотида
- 7. Нуклеотиды - фосфорные эфиры нуклеозидов. Остаток фосфорной кислоты присоединён к 5'-углеродному атому пентозы (5'-фосфоэфирная связь).
- 8. Азотистые основания Ароматические гетероциклические соединения, производные пиримидина и пурина.
- 9. Пентоза
- 10. Строение – структура - функция
- 11. Репликация – синтез дочерней молекулы двухцепочечной ДНК, идентичной родительской двухцепочечной ДНК (матрица – нити родительской ДНК).
- 12. Схема реализации генетической информации в фенотипические признаки. Реализацию потока информации в клетке можно представить схемой ДНК-"РНК-"белок.
- 13. Структура ДНК Первичная структура ДНК – порядок чередования дезоксирибонуклеозидмонофосфатов (дНМФ) в полинуклеотидной цепи. Связь между нуклеотидами
- 14. Фрагмент цепи ДНК
- 15. Макромолекулярная структура ДНК В 1953 г. Дж. Уотсон и Ф. Крик предложили модель структуры ДНК. При
- 16. Модель пространственной структуры ДНК. Согласно этой модели, молекула ДНК имеет форму спирали, образованную двумя полинуклеотидными цепями,
- 17. Связи, участвующие в образовании вторичной структуры ДНК Водородные связи между комплементарными азотистыми основаниями; Комплементарые основания уложены
- 18. Правило Чаргаффа: число пуриновых оснований (А + G) равно числу пиримидиновых оснований (Т + С).
- 19. Формы ДНК и их характеристики Для человека характерна B - форма спирали. Именно она и была
- 20. Третичная структура ДНК (суперспирализация ДНК). Компактизация и суперспирализация ДНК осуществляются с помощью разнообразных белков, взаимодействующих с
- 21. Все связывающиеся с ДНК эукариотов белки можно разделить на 2 группы: гисгоновые и негистоновые белки. Гистоны
- 22. Структура нуклеосом. Восемь молекул гистонов (Н2А, Н2В, НЗ, Н4)2 составляют ядро нуклеосомы, вокруг которого ДНК образует
- 23. РНК Первичная структура РНК - порядок чередования рибонуклеозидмонофосфатов (НМФ) в полинуклеотидной. Вторичная структура РНК Молекула рибонуклеиновой
- 24. Основные типы РНК Матричная (информационная) РНК (мРНК) Рибосомальная РНК (рРНК) Транспортная РНК Малые ядерные РНК (мяРНК)
- 25. Процессинг мРНК Процессинг РНК – совокупность процессов в клетке, которые способствуют превращению первичных транскриптов (пре –
- 26. Ковалентная модификация концевых нуклеотидных остатков первичного транскрипта мРНК.
- 27. Сплайсинг В процессе сплайсинга принимают участие различные малые ядерные рибонуклеопротеины (мяРНП), которые формируют сплайсосому. мяРНП, взаимодействуя
- 28. Предшественники рРНК и тРНК – таже как и мРНК подвергаются в ядре химической модификации (процессингу).
- 29. Процессинг первичного транскрипта тРНК Процессинг тРНК включает формирование последовательности -ССА на 3'-конце (акцепторный конец), к которому
- 30. Процессинг первичного транскрипта рРНК – формирование рибосом. Рибосомы – место синтеза белка. Представляют собой комплекс белков
- 31. Занятие № 2 Методы молекулярной биологии
- 32. Выделение НК Основными требованиями, предъявляемыми к методу выделения, являются эффективное отделение нуклеиновых кислот от белков, а
- 33. Выделение нуклеиновых кислот основано на методе разделения веществ при помощи магнитных шариков
- 34. Определение концентрации и качества препаратов нуклеиновых кислот методом спектрофотомерии Спектрофотометрия (абсорбционная) – физико-химический метод исследования растворов
- 35. Секвенирование Секвенирование (sequencing) – это общее название методов, которые позволяют установить последовательность нуклеотидов в молекуле ДНК.
- 36. В настоящее время для секвенирования генов обычно применяют метод Сенгера с дидезоксинуклеозидтрифосфатами (ddNTP). Обычно до начала
- 37. Секвенирование ДНК по Сенгеру Пусть есть образец ДНК, надо узнать его последовательность. Возьмем раствор, содержащий наш
- 38. Секвенирование ДНК по Сенгеру А теперь разобьем всю смесь на 4 пробирки, в которых содержатся в
- 39. Секвенирование ДНК по Сенгеру Позволим полимеразе работать в каждой из смесей. Для каждой молекулы образца она
- 40. Применение в медицине: Микробиология: 1) Выявление патогенов: холера – на Гавайях, токсичная E.coli в Германии. 2)
- 41. Секвенирование ДНК по Максаму и Гилберту: метод химической деградации В 1976 г. А. Максамом и У.
- 42. Пиросеквенирование Это метод секвенирования ДНК (определение последовательности нуклеотидов в молекуле ДНК), основанный на принципе «секвенирование путём
- 43. MS - анализ Метод масс-спектрометрии основан на пространственной или временной сепарации различающихся по массе и предварительно
- 45. Рентгеноструктурный анализ
- 48. Скачать презентацию

В каждом живом организме присутствуют 2 типа нуклеиновых кислот: рибонуклеиновая
В каждом живом организме присутствуют 2 типа нуклеиновых кислот: рибонуклеиновая

История открытия
60 – е года 19 в. – швейцарский ученый Фридрих
История открытия
60 – е года 19 в. – швейцарский ученый Фридрих

Локализация в клетке
Основная часть ДНК находится в ядре клетки – в
Локализация в клетке
Основная часть ДНК находится в ядре клетки – в

Структурная организация
НК – полимеры, мономерами которых являются нуклеотиды и выполняющие в
Структурная организация
НК – полимеры, мономерами которых являются нуклеотиды и выполняющие в
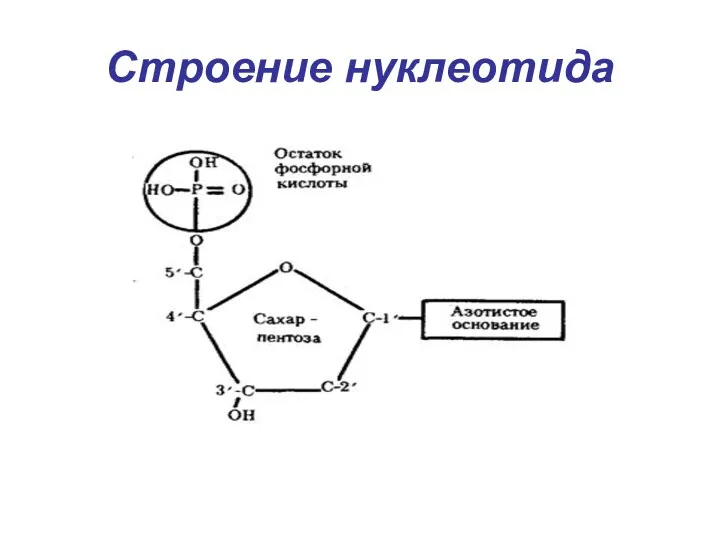
Строение нуклеотида
Строение нуклеотида
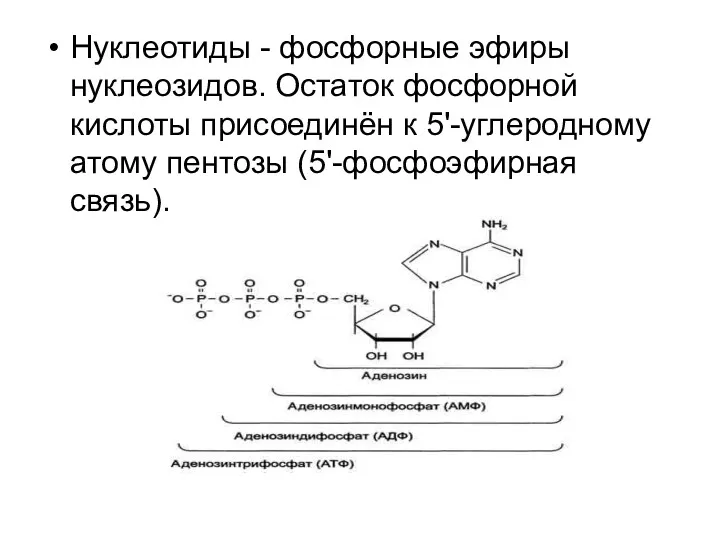
Нуклеотиды - фосфорные эфиры нуклеозидов. Остаток фосфорной кислоты присоединён к 5'-углеродному
Нуклеотиды - фосфорные эфиры нуклеозидов. Остаток фосфорной кислоты присоединён к 5'-углеродному
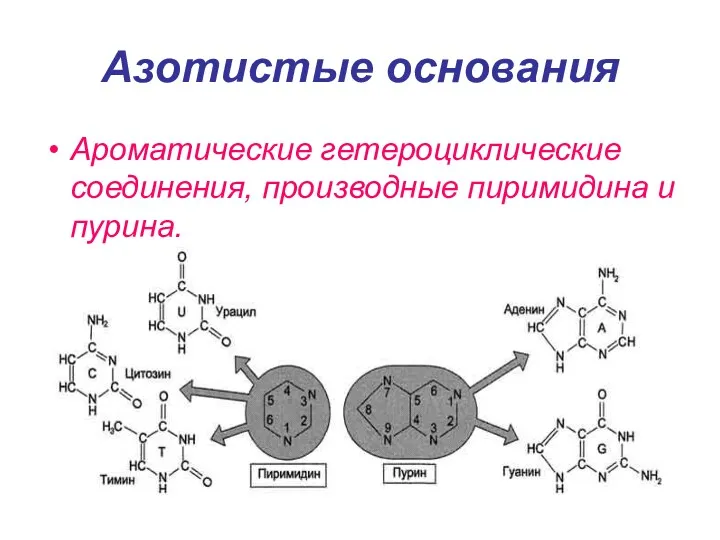
Азотистые основания
Ароматические гетероциклические соединения, производные пиримидина и пурина.
Азотистые основания
Ароматические гетероциклические соединения, производные пиримидина и пурина.
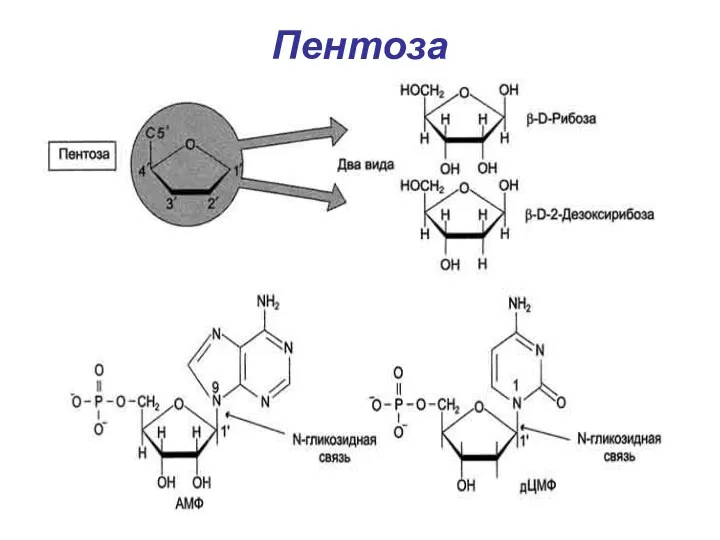
Пентоза
Пентоза
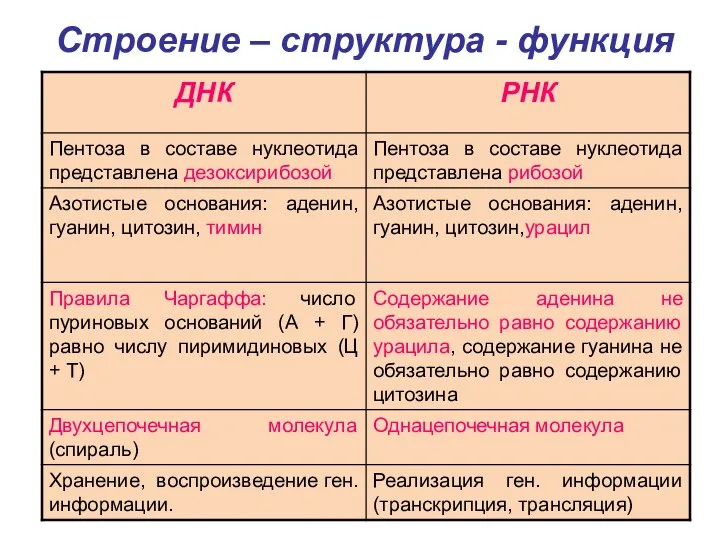
Строение – структура - функция
Строение – структура - функция

Репликация – синтез дочерней молекулы двухцепочечной ДНК, идентичной родительской двухцепочечной ДНК
Репликация – синтез дочерней молекулы двухцепочечной ДНК, идентичной родительской двухцепочечной ДНК
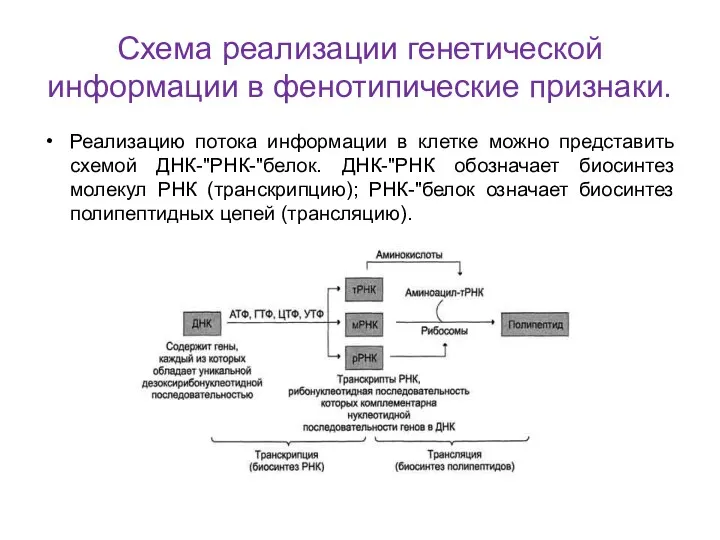
Схема реализации генетической информации в фенотипические признаки.
Реализацию потока информации в
Схема реализации генетической информации в фенотипические признаки.
Реализацию потока информации в

Структура ДНК
Первичная структура ДНК – порядок чередования дезоксирибонуклеозидмонофосфатов (дНМФ) в полинуклеотидной
Структура ДНК
Первичная структура ДНК – порядок чередования дезоксирибонуклеозидмонофосфатов (дНМФ) в полинуклеотидной
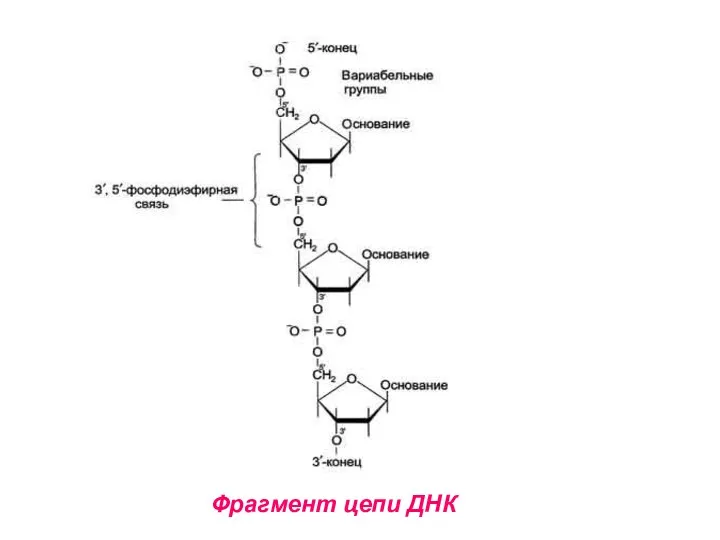
Фрагмент цепи ДНК
Фрагмент цепи ДНК

Макромолекулярная структура ДНК
В 1953 г. Дж. Уотсон и Ф. Крик
Макромолекулярная структура ДНК
В 1953 г. Дж. Уотсон и Ф. Крик
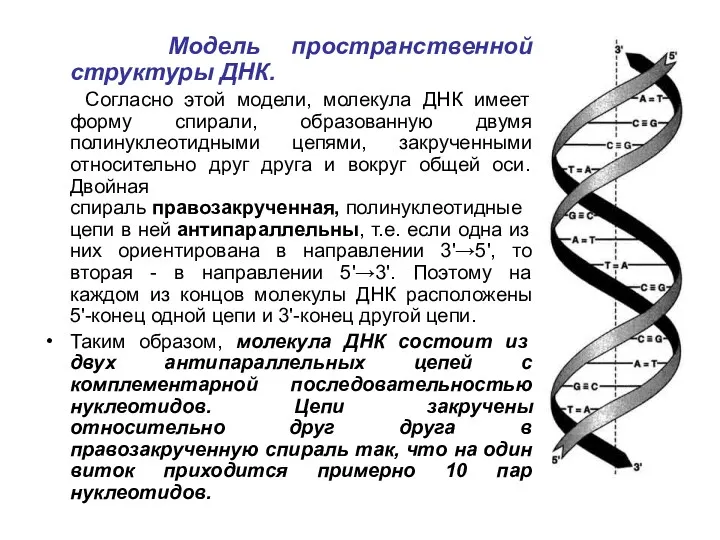
Модель пространственной структуры ДНК.
Согласно этой модели, молекула ДНК имеет
Модель пространственной структуры ДНК.
Согласно этой модели, молекула ДНК имеет

Связи, участвующие в образовании вторичной структуры ДНК
Водородные связи между комплементарными азотистыми
Связи, участвующие в образовании вторичной структуры ДНК
Водородные связи между комплементарными азотистыми
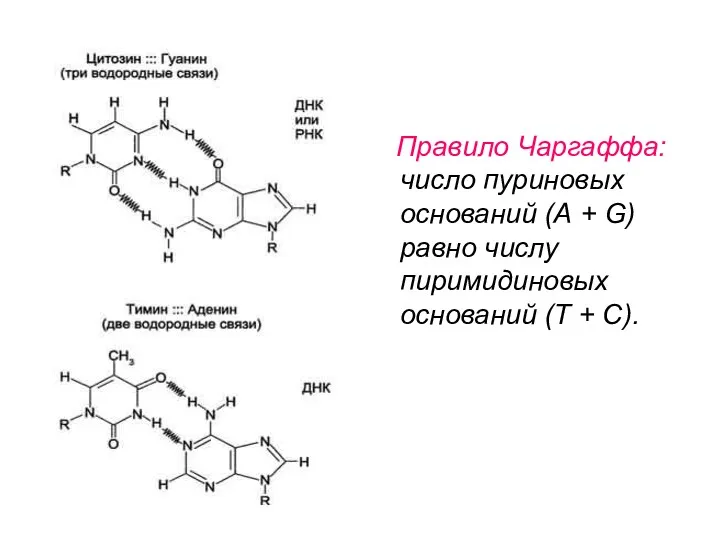
Правило Чаргаффа: число пуриновых оснований (А + G) равно числу
Правило Чаргаффа: число пуриновых оснований (А + G) равно числу
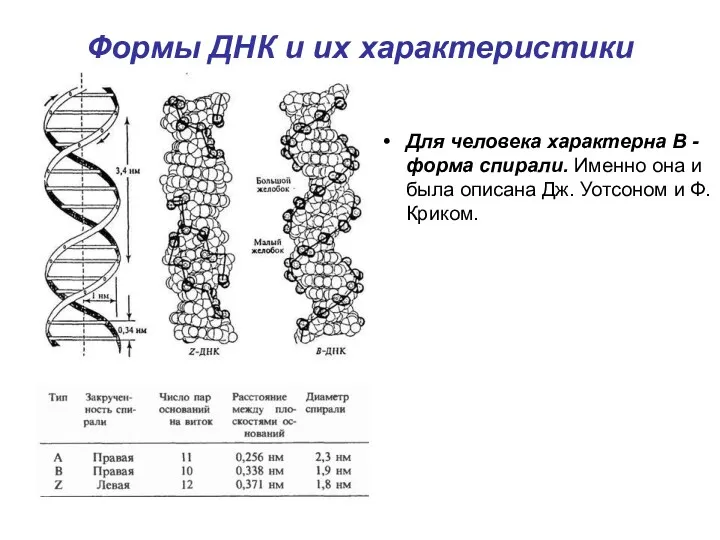
Формы ДНК и их характеристики
Для человека характерна B - форма
Формы ДНК и их характеристики
Для человека характерна B - форма

Третичная структура ДНК (суперспирализация ДНК). Компактизация и суперспирализация ДНК осуществляются с
Третичная структура ДНК (суперспирализация ДНК). Компактизация и суперспирализация ДНК осуществляются с

Все связывающиеся с ДНК эукариотов белки можно разделить на 2 группы: гисгоновые
Все связывающиеся с ДНК эукариотов белки можно разделить на 2 группы: гисгоновые
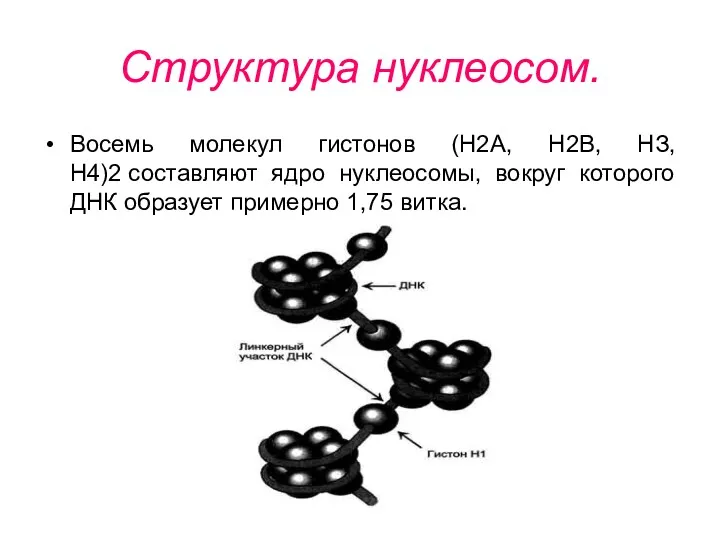
Структура нуклеосом.
Восемь молекул гистонов (Н2А, Н2В, НЗ, Н4)2 составляют ядро нуклеосомы,
Структура нуклеосом.
Восемь молекул гистонов (Н2А, Н2В, НЗ, Н4)2 составляют ядро нуклеосомы,

РНК
Первичная структура РНК - порядок чередования рибонуклеозидмонофосфатов (НМФ) в полинуклеотидной.
Вторичная структура РНК
РНК
Первичная структура РНК - порядок чередования рибонуклеозидмонофосфатов (НМФ) в полинуклеотидной.
Вторичная структура РНК

Основные типы РНК
Матричная (информационная) РНК (мРНК)
Рибосомальная РНК (рРНК)
Транспортная РНК
Малые ядерные РНК
Основные типы РНК
Матричная (информационная) РНК (мРНК)
Рибосомальная РНК (рРНК)
Транспортная РНК
Малые ядерные РНК

Процессинг мРНК
Процессинг РНК – совокупность процессов в клетке, которые способствуют превращению
Процессинг мРНК
Процессинг РНК – совокупность процессов в клетке, которые способствуют превращению
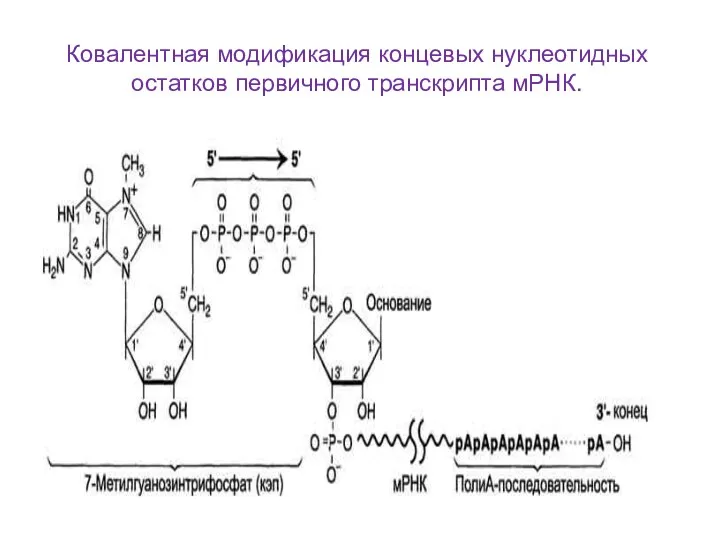
Ковалентная модификация концевых нуклеотидных остатков первичного транскрипта мРНК.
Ковалентная модификация концевых нуклеотидных остатков первичного транскрипта мРНК.
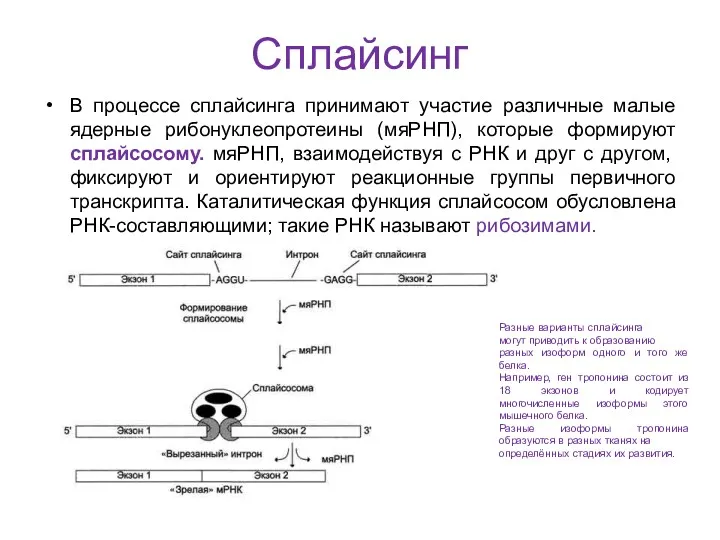
Сплайсинг
В процессе сплайсинга принимают участие различные малые ядерные рибонуклеопротеины (мяРНП), которые
Сплайсинг
В процессе сплайсинга принимают участие различные малые ядерные рибонуклеопротеины (мяРНП), которые

Предшественники рРНК и тРНК – таже как и мРНК подвергаются в
Предшественники рРНК и тРНК – таже как и мРНК подвергаются в
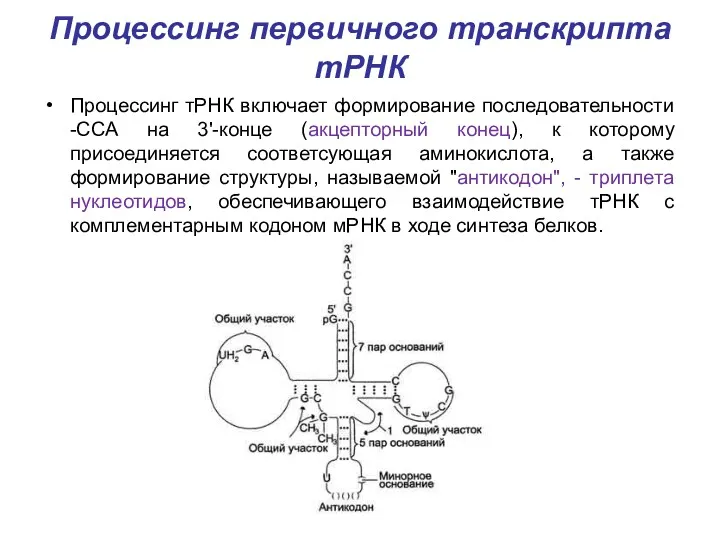
Процессинг первичного транскрипта тРНК
Процессинг тРНК включает формирование последовательности -ССА на 3'-конце
Процессинг первичного транскрипта тРНК
Процессинг тРНК включает формирование последовательности -ССА на 3'-конце
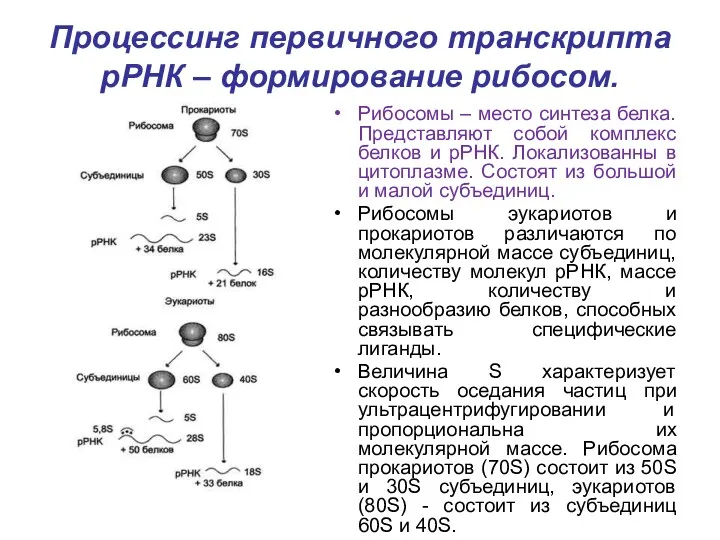
Процессинг первичного транскрипта рРНК – формирование рибосом.
Рибосомы – место синтеза белка.
Процессинг первичного транскрипта рРНК – формирование рибосом.
Рибосомы – место синтеза белка.

Занятие № 2
Методы молекулярной биологии
Занятие № 2
Методы молекулярной биологии

Выделение НК
Основными требованиями, предъявляемыми к методу выделения, являются эффективное отделение нуклеиновых
Выделение НК
Основными требованиями, предъявляемыми к методу выделения, являются эффективное отделение нуклеиновых
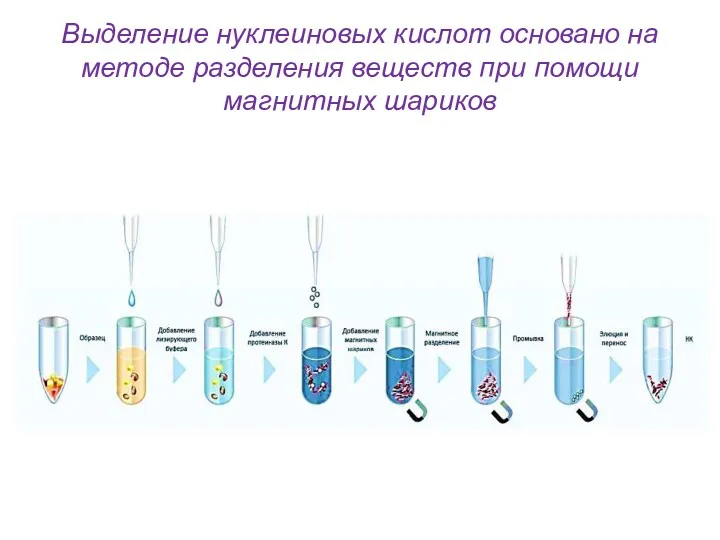
Выделение нуклеиновых кислот основано на методе разделения веществ при помощи магнитных
Выделение нуклеиновых кислот основано на методе разделения веществ при помощи магнитных

Определение концентрации и качества препаратов нуклеиновых кислот методом спектрофотомерии
Спектрофотометрия (абсорбционная) –
Определение концентрации и качества препаратов нуклеиновых кислот методом спектрофотомерии
Спектрофотометрия (абсорбционная) –

Секвенирование
Секвенирование (sequencing) – это общее название методов, которые позволяют установить последовательность
Секвенирование
Секвенирование (sequencing) – это общее название методов, которые позволяют установить последовательность

В настоящее время для секвенирования генов обычно применяют метод Сенгера с дидезоксинуклеозидтрифосфатами
В настоящее время для секвенирования генов обычно применяют метод Сенгера с дидезоксинуклеозидтрифосфатами

Секвенирование ДНК по Сенгеру
Пусть есть образец ДНК, надо узнать его последовательность.
Возьмем
Секвенирование ДНК по Сенгеру
Пусть есть образец ДНК, надо узнать его последовательность.
Возьмем
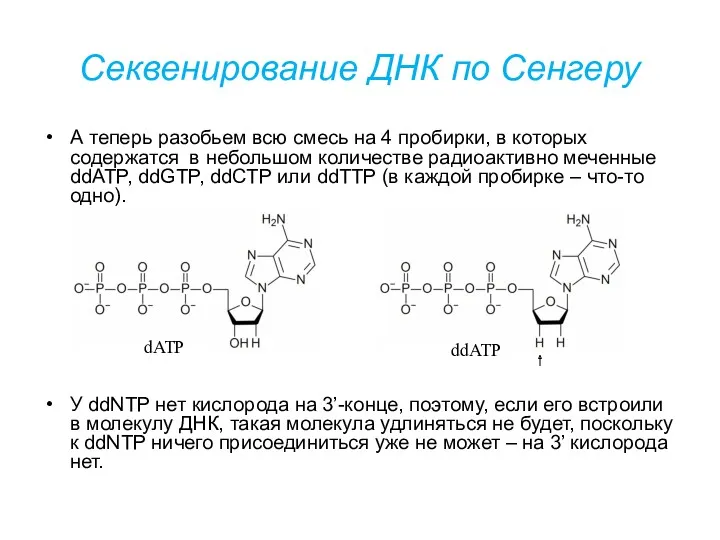
Секвенирование ДНК по Сенгеру
А теперь разобьем всю смесь на 4 пробирки,
Секвенирование ДНК по Сенгеру
А теперь разобьем всю смесь на 4 пробирки,

Секвенирование ДНК по Сенгеру
Позволим полимеразе работать в каждой из смесей. Для
Секвенирование ДНК по Сенгеру
Позволим полимеразе работать в каждой из смесей. Для

Применение в медицине:
Микробиология:
1) Выявление патогенов: холера – на Гавайях, токсичная E.coli
Применение в медицине:
Микробиология:
1) Выявление патогенов: холера – на Гавайях, токсичная E.coli

Секвенирование ДНК по Максаму и Гилберту: метод химической деградации
В 1976 г.
Секвенирование ДНК по Максаму и Гилберту: метод химической деградации
В 1976 г.

Пиросеквенирование
Это метод секвенирования ДНК (определение последовательности нуклеотидов в молекуле ДНК), основанный на принципе «секвенирование путём синтеза».
Пиросеквенирование
Это метод секвенирования ДНК (определение последовательности нуклеотидов в молекуле ДНК), основанный на принципе «секвенирование путём синтеза».
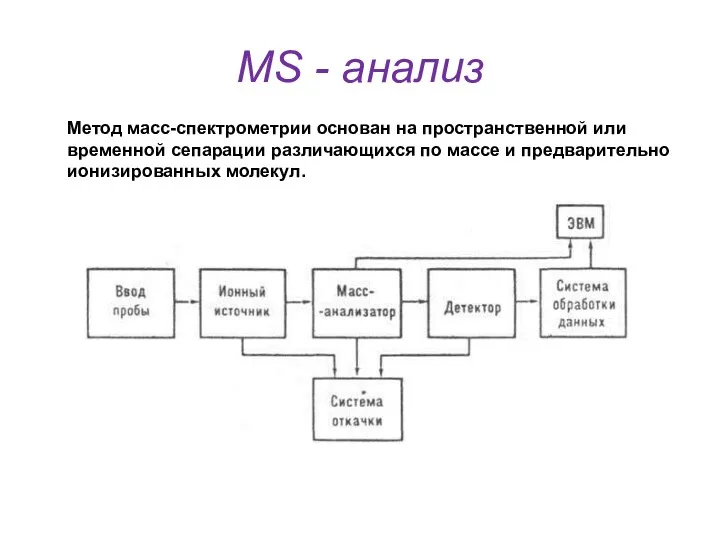
MS - анализ
Метод масс-спектрометрии основан на пространственной или
временной сепарации различающихся
MS - анализ
Метод масс-спектрометрии основан на пространственной или
временной сепарации различающихся

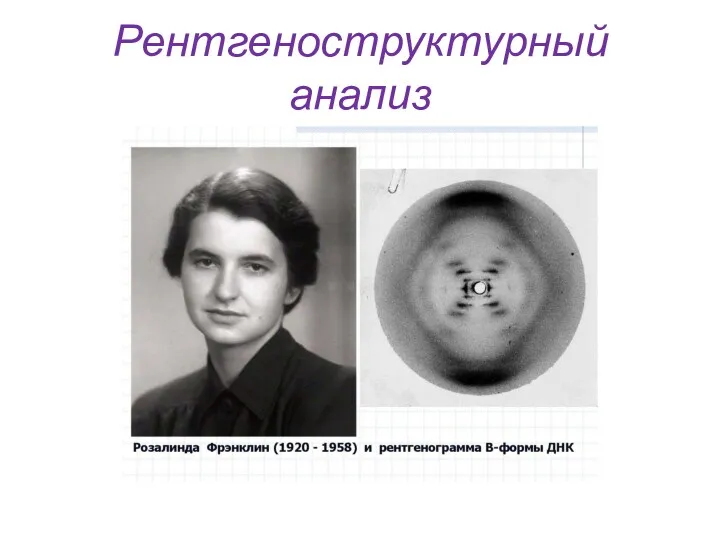
Рентгеноструктурный анализ
Рентгеноструктурный анализ

 Оценка иммунного статуса человека
Оценка иммунного статуса человека Биохимия гормонов
Биохимия гормонов Краснокнижные виды растений и животных РТ
Краснокнижные виды растений и животных РТ Царство Растения
Царство Растения Клеточная теория. Цитология
Клеточная теория. Цитология Строение и работа сердца. 8 класс
Строение и работа сердца. 8 класс Рептилії
Рептилії Обмен веществ и энергии. Питательные вещества. Пищевые продукты
Обмен веществ и энергии. Питательные вещества. Пищевые продукты Хвойные растения
Хвойные растения Биологические науки
Биологические науки Возврат органики в почву. Грядка Розума
Возврат органики в почву. Грядка Розума Выращивание гороха в условиях искусственного лета
Выращивание гороха в условиях искусственного лета Общие черты организации. Монадный тип организации
Общие черты организации. Монадный тип организации Строение кожи и терморегуляция.
Строение кожи и терморегуляция. Выявление ароморфозов и идиоадаптаций у растений
Выявление ароморфозов и идиоадаптаций у растений Презентация Тип Кишечнополостные
Презентация Тип Кишечнополостные Генеративные органы растения
Генеративные органы растения Жизнь в океане
Жизнь в океане Основы ресурсосберегающей технологии возделывания
Основы ресурсосберегающей технологии возделывания Зоология - наука о животных. Подготовка к ОГЭ и ЕГЭ по биологии
Зоология - наука о животных. Подготовка к ОГЭ и ЕГЭ по биологии Психогенетические исследования моторной деятельности и ее вегетативной регуляции
Психогенетические исследования моторной деятельности и ее вегетативной регуляции Строение и жизнедеятельность бактерий
Строение и жизнедеятельность бактерий Общая характеристика мхов. Отдел Bryophyta - Мохообразные
Общая характеристика мхов. Отдел Bryophyta - Мохообразные Строение клетки клеточная мембрана. Ядро
Строение клетки клеточная мембрана. Ядро Чибис - птица 2010 года
Чибис - птица 2010 года Парнокопытные и непарнокопытные животные
Парнокопытные и непарнокопытные животные Самые опасные насекомые в мире
Самые опасные насекомые в мире Разработка урока по теме:Экологические факторы среды и их влияние на живые организмы.
Разработка урока по теме:Экологические факторы среды и их влияние на живые организмы.