- Общие черты организации. Монадный тип организации

Содержание
- 2. Царство Protista, Подцарство Protozoa 1. Общая характеристика К подцарству Protozoa относятся гетеротрофные эукариоты, среди которых есть
- 3. Элементарной единицей организации простейших служит так называемая энергида. Энергида – это часть клетки с ядром и
- 4. Форма тела простейших, их размер и окраска чрезвычайно разнообразны и зависят от условий сред обитания. Об
- 5. Монадный (жгутиковый) тип организации объединяет организмы, у которых во взрослом состоянии имеется жгутик (жгутики) или реснички
- 6. Детальное изучение тонкого строения жгутиконосцев (покровных структур клетки, жгутикового аппарата, митохондрий и хлоропластов и т.д.), их
- 7. Выделяют 4 морфотипа жгутиконосцев: изоконты, анизоконты, гетероконты и стефаноконты. - изоконты с 2-8 жгутиками равной длины,
- 8. Тип. Chlorophyta Колония Volvox sp. Колонии последнего представляют собой слизистую сферу, полую внутри, которая объединяет лежащие
- 9. Надтип Euglenozoa: Tип Euglenophyta Надтип Dinomorpha Панцирные жгутиконосцы
- 10. Надтип Polymastigota, Tип Diplomonadida
- 11. Тип Diplomonadida В состав типа входят свободноживущие и паразитические жгутиконосцы. Мастигонт состоит из 4 кинетосом, одна
- 12. Тип Kinetoplastida
- 13. Tип Kinetoplastida: Класс Bodomonadida, Класс Trypanosomamonadida
- 14. Лейшмании используют в качестве хозяев широкий круг позвоночных животных: рыб, рептилий, млекопита-ющих, в том числе человека,
- 15. Трипаносомы: болезнь Чагаса – через поцелуйного клопа от домашних животных к человеку в Лат.Америке. Возбудитель другой
- 16. Tип Parabasalia:, Класс Тrichomonadеа Это обширная группа, к которой относятся жгутиконосцы, обладающие 4-6 жгутиками. Один из
- 17. Класс Нуреrmastiginea Это относительно крупные жгутиконосцы (до 0,5 мм длиной), обитатели кщиечника насекомых, питающихся древесиной. Для
- 18. Среди одноклеточных протистов Ciliophora характеризуются наиболее сложным строением. Их внешняя морфология очень разнообразна. Для них в
- 20. 2. Покровные структуры инфузорий включают альвеолярную пелликулу и кортикальный ц и т о с к е
- 21. С кинетосомами связаны корешки, залегающие в субальвеолярной зоне параллельно поверхности тела. Обычно от одной кинетосомы отходят
- 22. Кроме того, реснички могут группироваться в относительно крупные «надресничные» агрегаты — м е м б р
- 23. Класс Oligohymenophorea объединяет в своем составе весьма разнообразных в морфологическом и биологическом отношениях инфузорий. Общим для
- 24. Класс Heterotrichea, или разноресничные инфузории,
- 25. К покровам приурочены специальные органеллы нападения и защиты - экструсомы. У инфузорий известны 12 типов экструсом
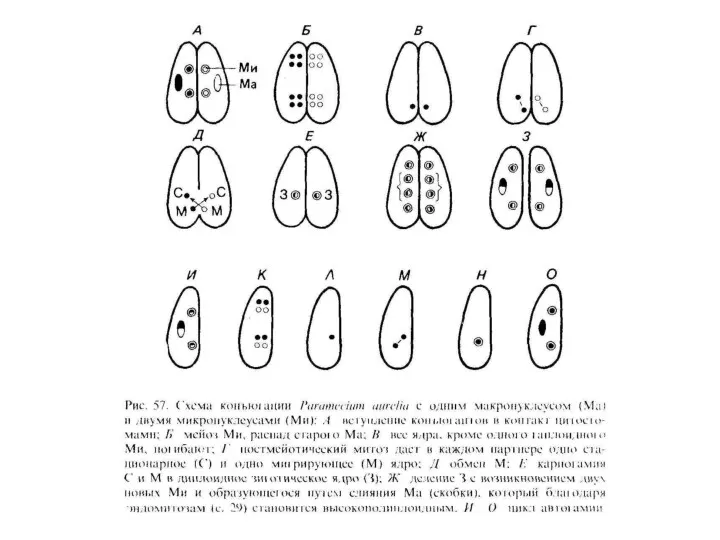
- 27. Для всех инфузорий характерен я д е р н ы й д у а л и
- 28. Размножаются инфузории только агамно. Чаще всего это бинарное поперечное деление. После расхождения дочерние клетки претерпевают сложный
- 30. Саркодовый тип организации Надтип Rhizopoda - Саркодовые - корненожки Тип Lobosea Класс Gymnamoebea – голые амебы
- 31. Саркодовый тип организации КОРНЕНОЖКИ (Надтип Rhizopoda) К Rhizopoda, или корненожкам, относятся фаготрофные, одно- и много-ядерные протисты,
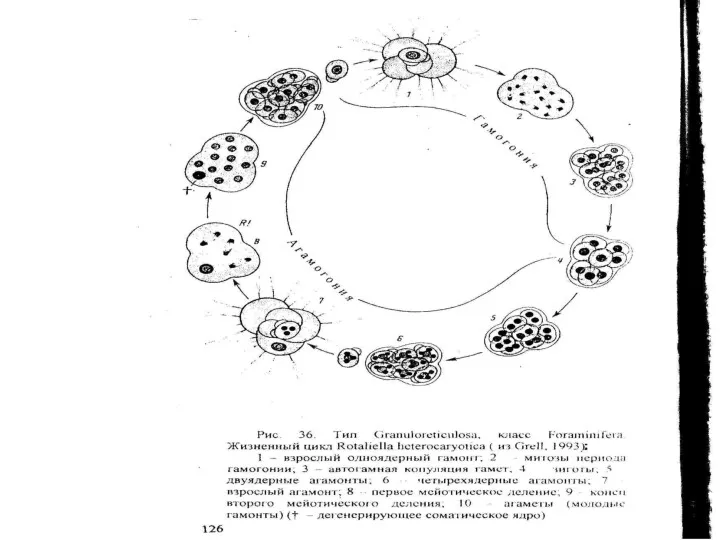
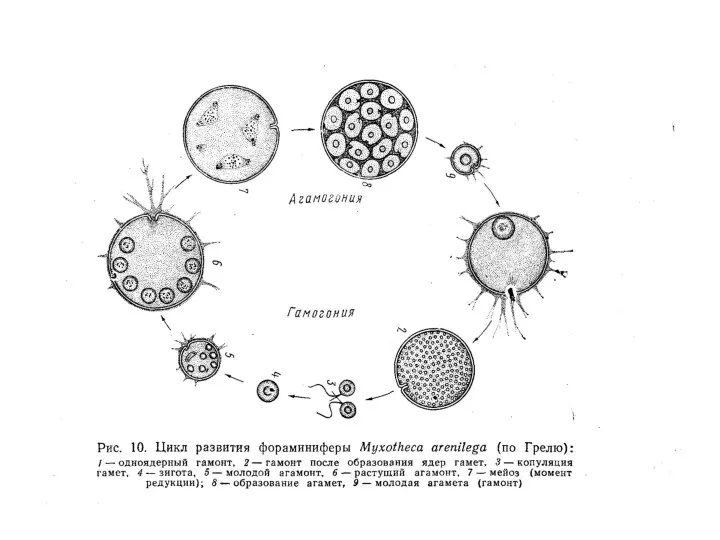
- 35. Тип Granuloreticulosea – раковинные амебы Класс Foraminifera – Фораминиферы Фораминиферы — гетеротрофные, преимущественно свободноживущие простейшие, характеризующиеся
- 37. 5. Фораминиферы обладают особыми псевдоподиями — г р а н у л о р е т
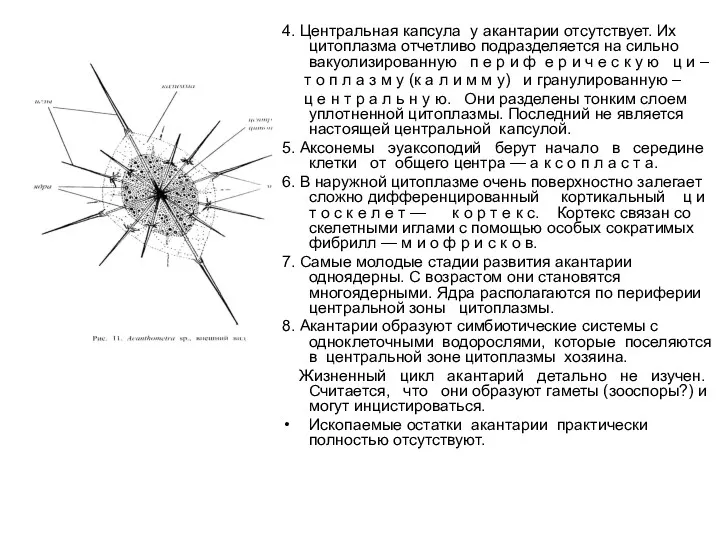
- 38. 3. У всех акантарий спикулы скелета построены из сульфата стронция (SrSO4 ), или ц е л
- 39. 4. Центральная капсула у акантарии отсутствует. Их цитоплазма отчетливо подразделяется на сильно вакуолизированную п е р
- 41. Подтип Euradiolaria Класс Polycystinea (полицистинеи) Класс Polycystinea объединяет в своем составе одиночные и колониальные планктонные формы
- 42. 3. Полицистинеи обладают настоящей центральной капсулой. Последняя представляет собой замкнутое мешковидное образование, стенки которого построены из
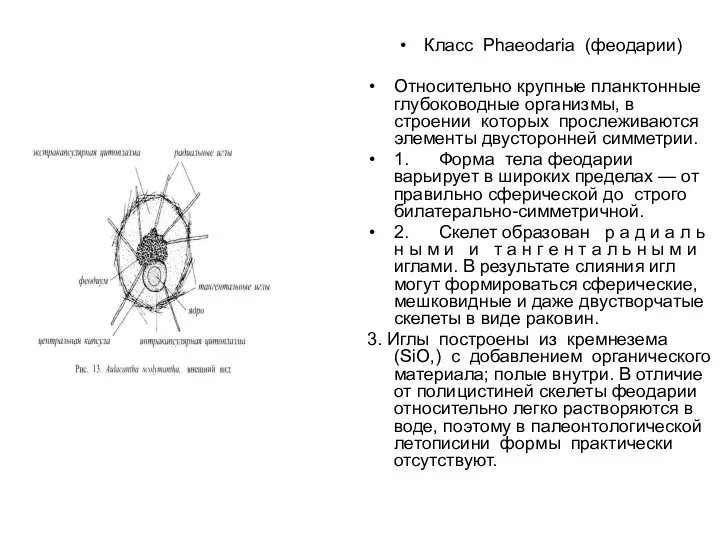
- 43. Класс Phaeodaria (феодарии) Относительно крупные планктонные глубоководные организмы, в строении которых прослеживаются элементы двусторонней симметрии. 1.
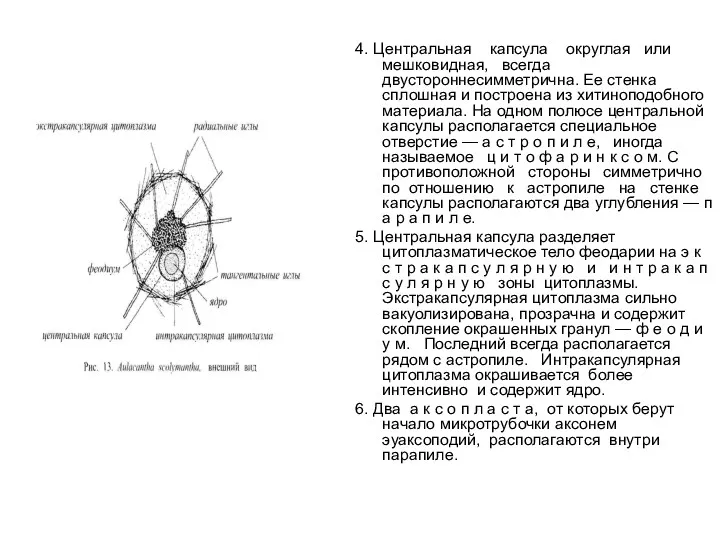
- 44. 4. Центральная капсула округлая или мешковидная, всегда двустороннесимметрична. Ее стенка сплошная и построена из хитиноподобного материала.
- 45. Тип Heliozoa Название «с о л н е ч н и к и» впервые было использовано
- 48. Надтип Apicomplexa: Тип Sporozoaе (Споровики): Класс Gregarinomorpha, Класс Coccidiomorphina, Отр. Сoccidiida, Отр. Haemosporidia, Тип Slopalinata: Класс
- 49. Тип Sporozoaе Для споровиков характерен сложный жизненный цикл, который обычно включает три последовательные фазы. Первая представлена
- 50. Главной отличительной особенностью Арiсоmplеха является то, что в их жизненном цикле обязательно присутствует стадия (вегетативная клетка—организм,
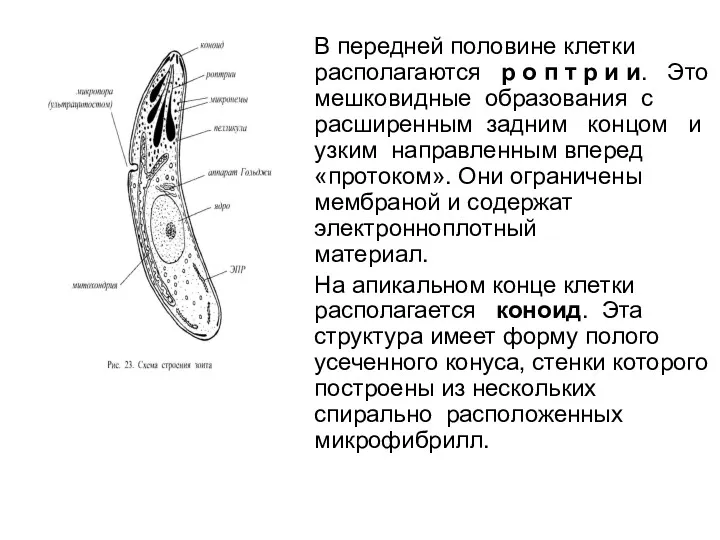
- 51. В передней половине клетки располагаются р о п т р и и. Это мешковидные образования с
- 52. Класс Gregarinomorphа В состав класса входят полостные и кишечные паразиты беспозвоночных и низших хордовых. Мужские и
- 53. Класс Gregarinomorpha
- 55. Класс Coccidiomorphina В состав класса входят преимущественно внутриклеточные паразиты беспозвоночных и позвоночных животных. Главной отличительной чертой
- 58. Отряд HAEMOSPORIDIA К кровяным споровикам относятся представители типа Sporozoae, паразитирующие в кровеносном русле позвоночных животных. Жизненный
- 60. Надтип Chromophyta Тип Slopalinata Класс Opalinatea Все представители класса - полиэнергидные простейшие. Число ядер у них
- 62. Скачать презентацию
Царство Protista,
Подцарство Protozoa
1. Общая характеристика
К подцарству Protozoa относятся гетеротрофные эукариоты, среди которых есть
Царство Protista,
Подцарство Protozoa
1. Общая характеристика
К подцарству Protozoa относятся гетеротрофные эукариоты, среди которых есть

Элементарной единицей организации простейших служит так называемая энергида. Энергида – это часть
Элементарной единицей организации простейших служит так называемая энергида. Энергида – это часть

Форма тела простейших, их размер и окраска чрезвычайно разнообразны и зависят от условий

Монадный (жгутиковый) тип организации
объединяет организмы, у которых во взрослом состоянии имеется жгутик
Монадный (жгутиковый) тип организации
объединяет организмы, у которых во взрослом состоянии имеется жгутик

Детальное изучение тонкого строения жгутиконосцев (покровных структур клетки, жгутикового аппарата, митохондрий и хлоропластов

Выделяют 4 морфотипа жгутиконосцев: изоконты, анизоконты, гетероконты и стефаноконты.
- изоконты с 2-8 жгутиками
Выделяют 4 морфотипа жгутиконосцев: изоконты, анизоконты, гетероконты и стефаноконты.
- изоконты с 2-8 жгутиками

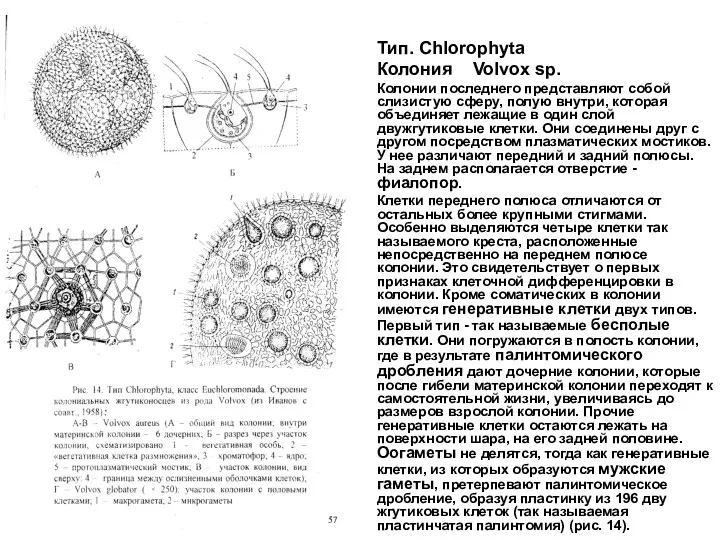
Тип. Chlorophyta
Колония Volvox sp.
Колонии последнего представляют собой слизистую сферу, полую внутри, которая
Тип. Chlorophyta
Колония Volvox sp.
Колонии последнего представляют собой слизистую сферу, полую внутри, которая
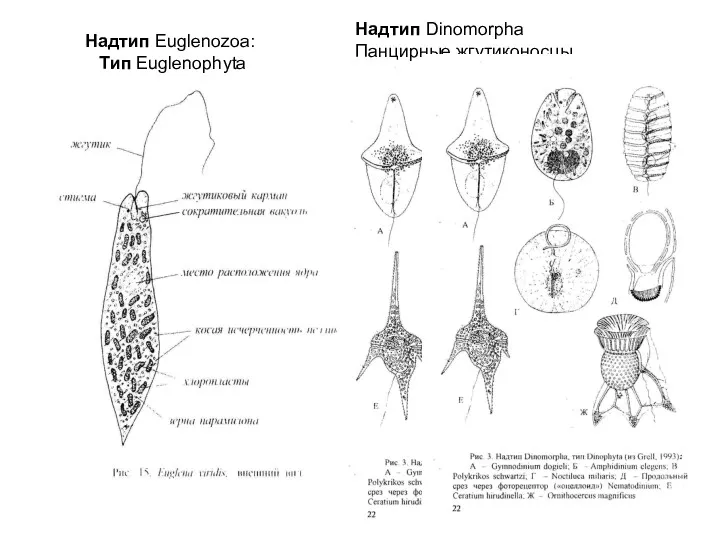
Надтип Euglenozoa:
Tип Euglenophyta
Надтип Dinomorpha
Панцирные жгутиконосцы
Надтип Euglenozoa:
Tип Euglenophyta
Надтип Dinomorpha
Панцирные жгутиконосцы
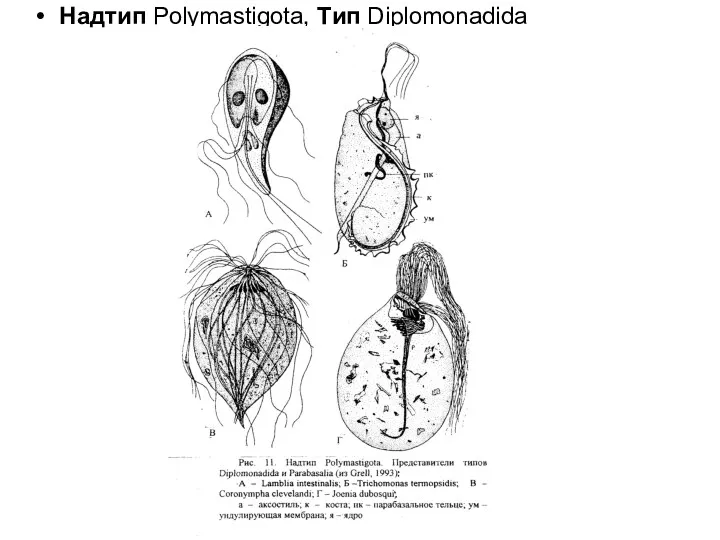
Надтип Polymastigota, Tип Diplomonadida
Надтип Polymastigota, Tип Diplomonadida
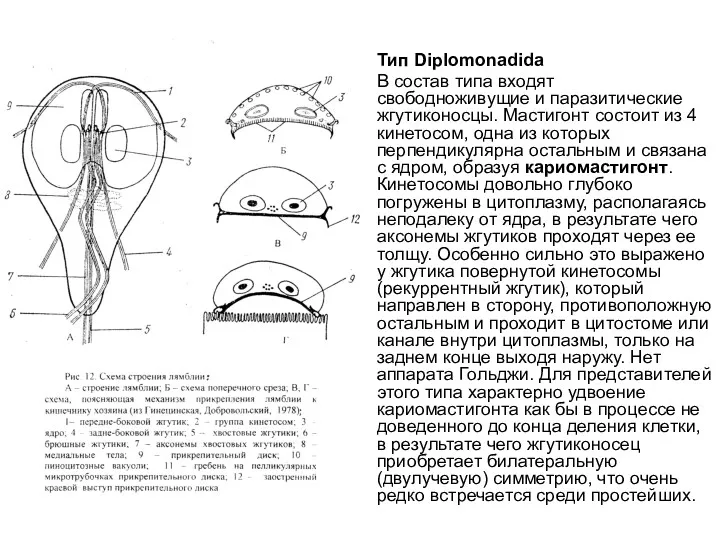
Тип Diplomonadida
В состав типа входят свободноживущие и паразитические жгутиконосцы. Мастигонт состоит из 4
Тип Diplomonadida
В состав типа входят свободноживущие и паразитические жгутиконосцы. Мастигонт состоит из 4
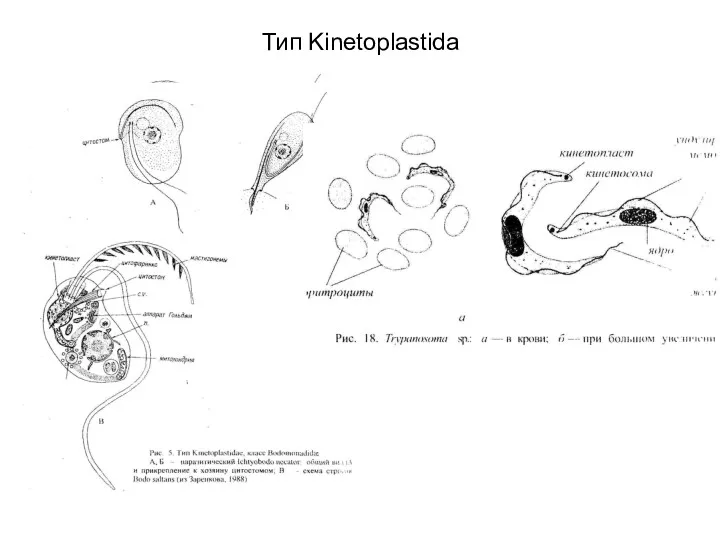
Тип Kinetoplastida
Тип Kinetoplastida
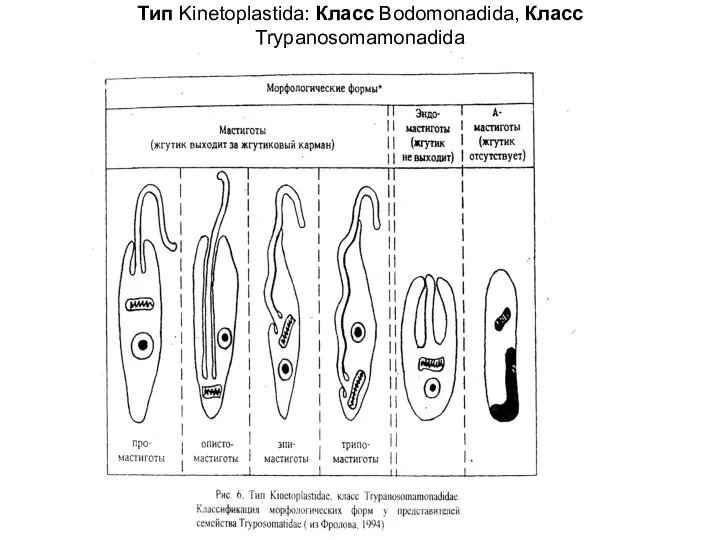
Tип Kinetoplastida: Класс Bodomonadida, Класс Trypanosomamonadida
Tип Kinetoplastida: Класс Bodomonadida, Класс Trypanosomamonadida
Лейшмании используют в качестве хозяев широкий круг позвоночных животных: рыб, рептилий, млекопита-ющих, в
Лейшмании используют в качестве хозяев широкий круг позвоночных животных: рыб, рептилий, млекопита-ющих, в

Трипаносомы: болезнь Чагаса – через поцелуйного клопа от домашних животных к человеку в
Трипаносомы: болезнь Чагаса – через поцелуйного клопа от домашних животных к человеку в

Tип Parabasalia:,
Класс Тrichomonadеа
Это обширная группа, к которой относятся жгутиконосцы, обладающие 4-6 жгутиками. Один
Tип Parabasalia:,
Класс Тrichomonadеа
Это обширная группа, к которой относятся жгутиконосцы, обладающие 4-6 жгутиками. Один

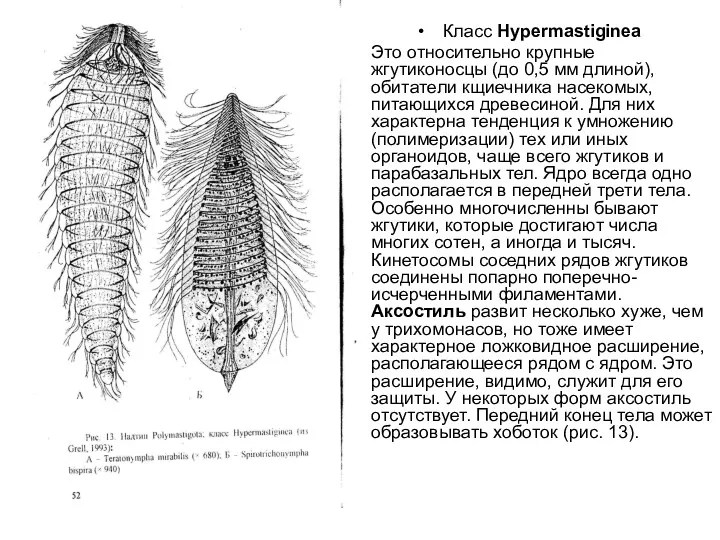
Класс Нуреrmastiginea
Это относительно крупные жгутиконосцы (до 0,5 мм длиной), обитатели кщиечника насекомых, питающихся
Класс Нуреrmastiginea
Это относительно крупные жгутиконосцы (до 0,5 мм длиной), обитатели кщиечника насекомых, питающихся
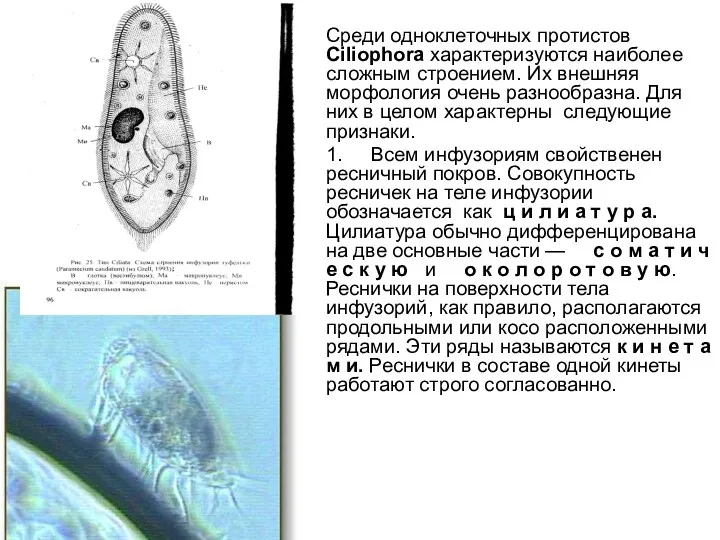
Среди одноклеточных протистов Ciliophora характеризуются наиболее сложным строением. Их внешняя морфология очень разнообразна.
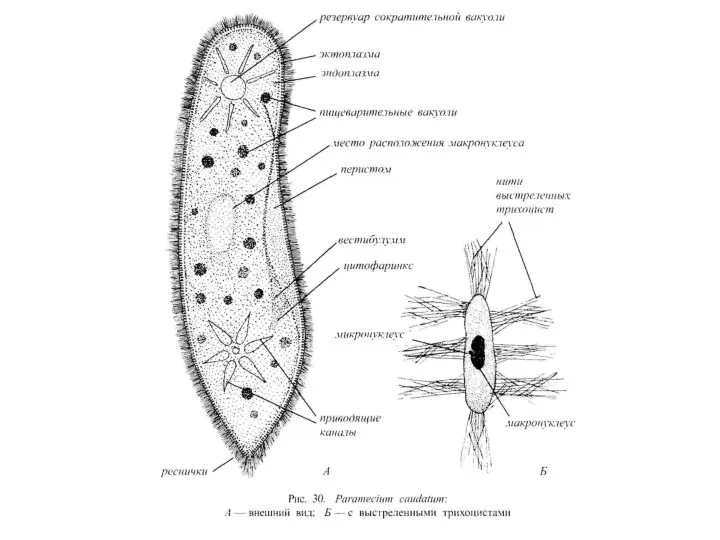
2. Покровные структуры инфузорий включают альвеолярную пелликулу и кортикальный ц и т о с
2. Покровные структуры инфузорий включают альвеолярную пелликулу и кортикальный ц и т о с

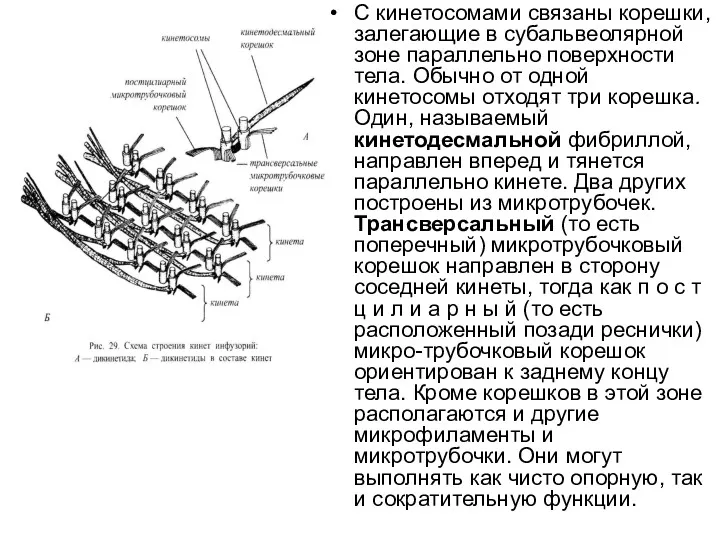
С кинетосомами связаны корешки, залегающие в субальвеолярной зоне параллельно поверхности тела. Обычно от
С кинетосомами связаны корешки, залегающие в субальвеолярной зоне параллельно поверхности тела. Обычно от
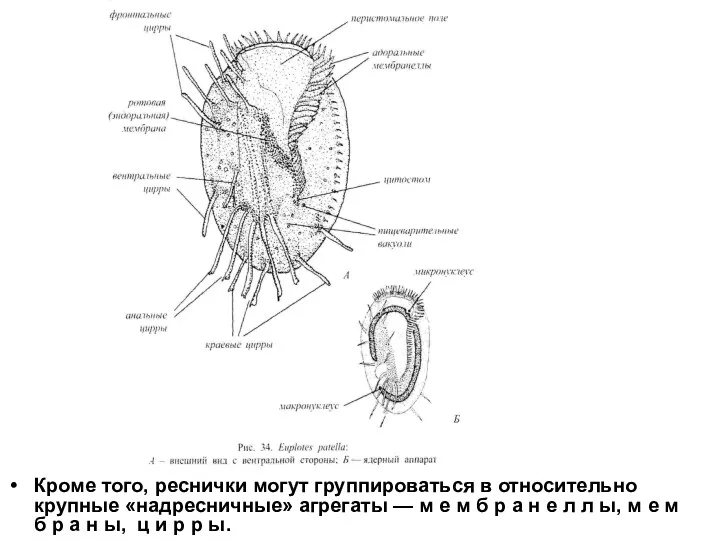
Кроме того, реснички могут группироваться в относительно крупные «надресничные» агрегаты — м е
Кроме того, реснички могут группироваться в относительно крупные «надресничные» агрегаты — м е
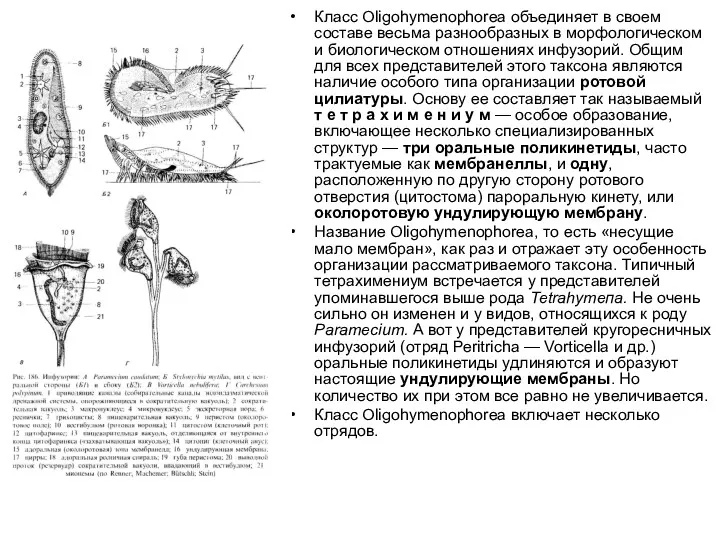
Класс Oligohymenophorea объединяет в своем составе весьма разнообразных в морфологическом и биологическом отношениях
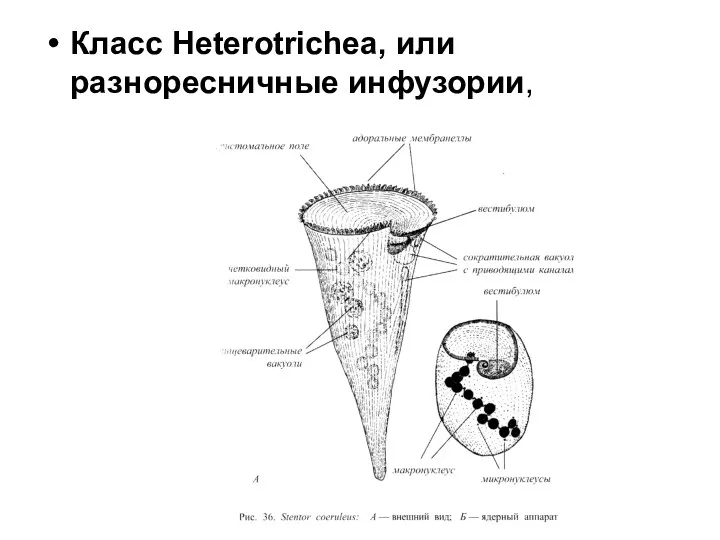
Класс Heterotrichea, или разноресничные инфузории,
Класс Heterotrichea, или разноресничные инфузории,
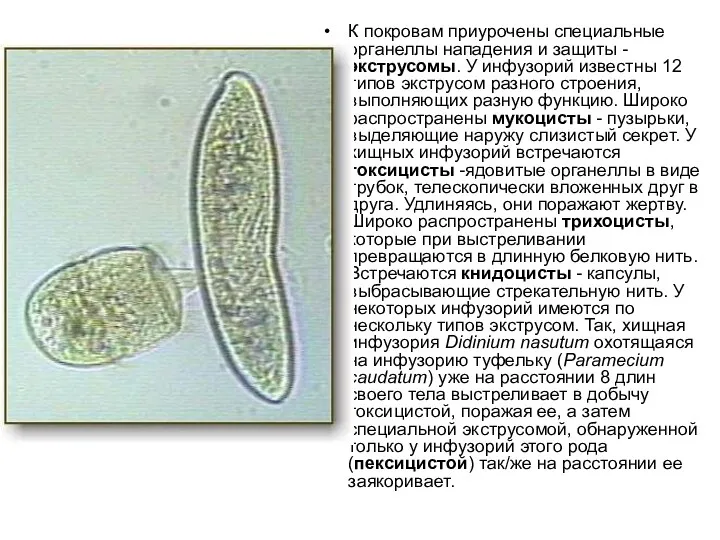
К покровам приурочены специальные органеллы нападения и защиты - экструсомы. У инфузорий известны
К покровам приурочены специальные органеллы нападения и защиты - экструсомы. У инфузорий известны
Для всех инфузорий характерен я д е р н ы й д у

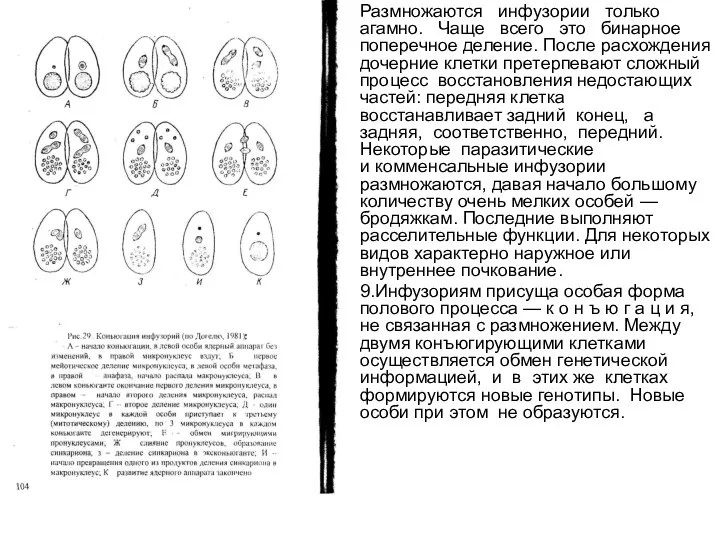
Размножаются инфузории только агамно. Чаще всего это бинарное поперечное деление. После расхождения дочерние
Размножаются инфузории только агамно. Чаще всего это бинарное поперечное деление. После расхождения дочерние
Саркодовый тип организации
Надтип Rhizopoda -
Саркодовые - корненожки
Тип Lobosea
Класс Gymnamoebea –
Саркодовый тип организации Надтип Rhizopoda - Саркодовые - корненожки Тип Lobosea Класс Gymnamoebea –

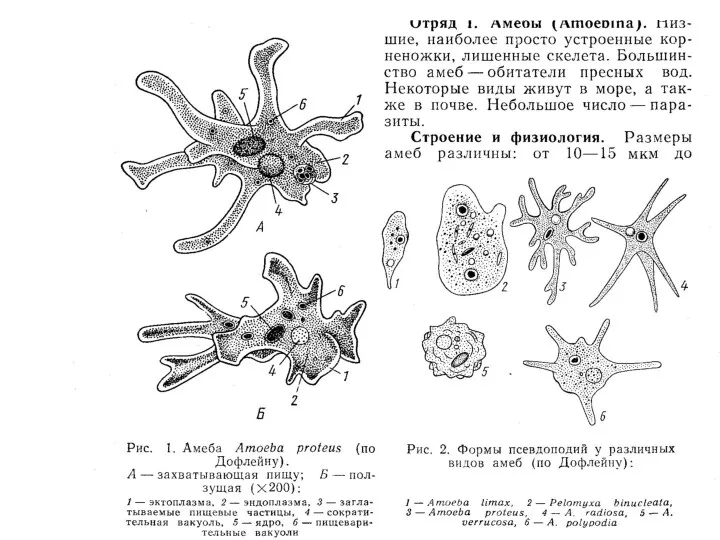
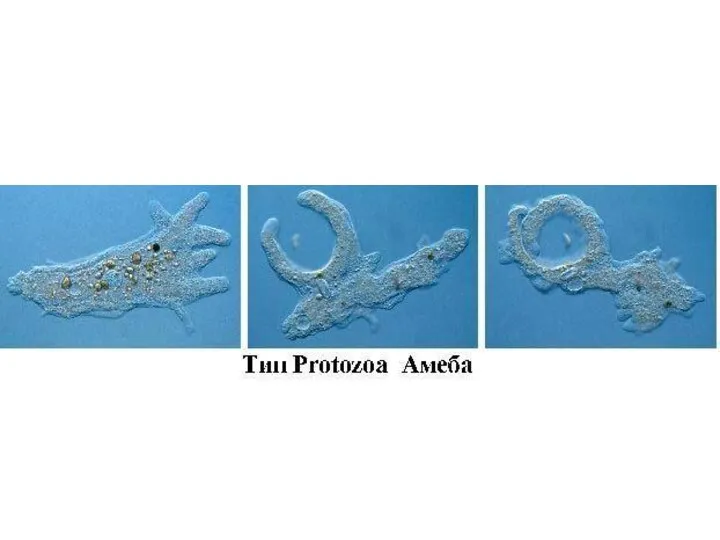
Саркодовый тип организации
КОРНЕНОЖКИ (Надтип Rhizopoda)
К Rhizopoda, или корненожкам, относятся фаготрофные, одно- и много-ядерные
Саркодовый тип организации
КОРНЕНОЖКИ (Надтип Rhizopoda)
К Rhizopoda, или корненожкам, относятся фаготрофные, одно- и много-ядерные

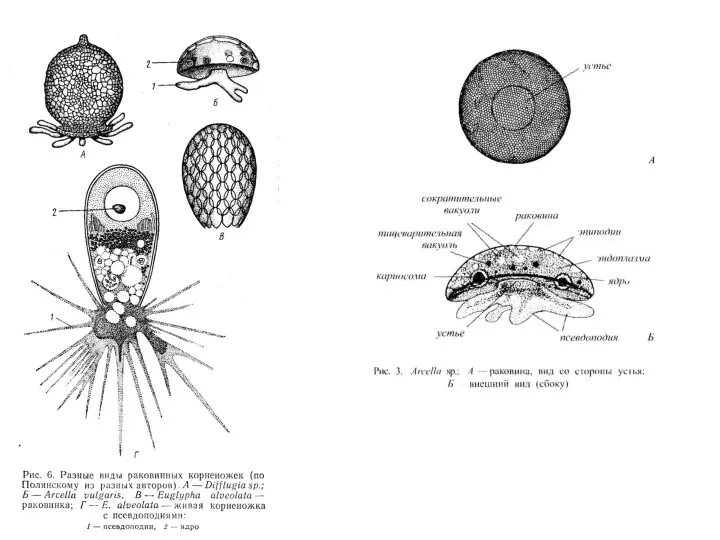
Тип Granuloreticulosea – раковинные амебы
Класс Foraminifera – Фораминиферы
Фораминиферы — гетеротрофные, преимущественно свободноживущие простейшие,
Класс Foraminifera – Фораминиферы
Фораминиферы — гетеротрофные, преимущественно свободноживущие простейшие,
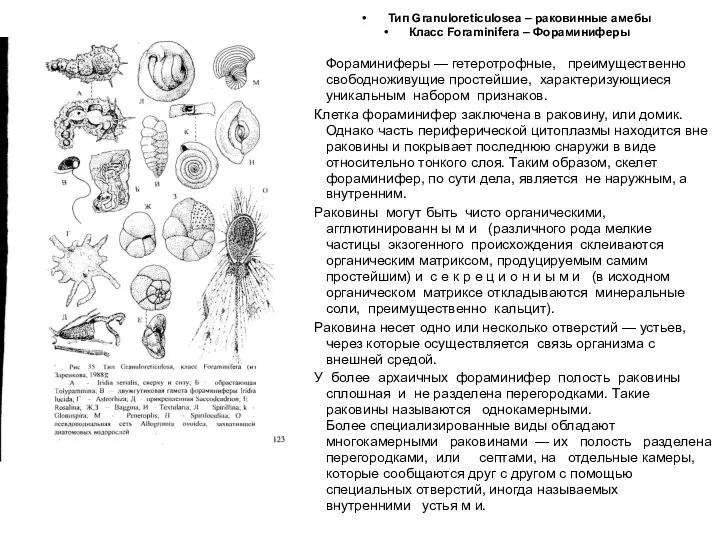

5. Фораминиферы обладают особыми псевдоподиями —
г р а н у л
г р а н у л

3. У всех акантарий спикулы скелета построены из сульфата стронция (SrSO4 ), или
3. У всех акантарий спикулы скелета построены из сульфата стронция (SrSO4 ), или

4. Центральная капсула у акантарии отсутствует. Их цитоплазма отчетливо подразделяется на сильно вакуолизированную
4. Центральная капсула у акантарии отсутствует. Их цитоплазма отчетливо подразделяется на сильно вакуолизированную
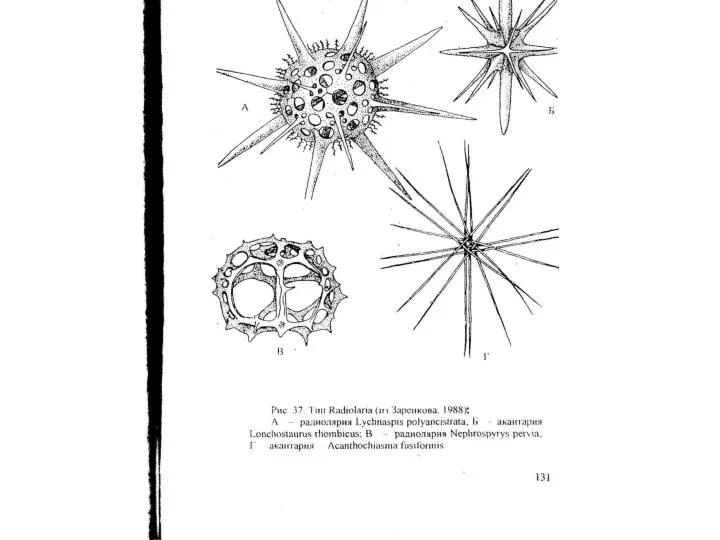
Подтип Euradiolaria
Класс Polycystinea (полицистинеи)
Класс Polycystinea объединяет в своем составе одиночные и колониальные планктонные
Класс Polycystinea (полицистинеи)
Класс Polycystinea объединяет в своем составе одиночные и колониальные планктонные
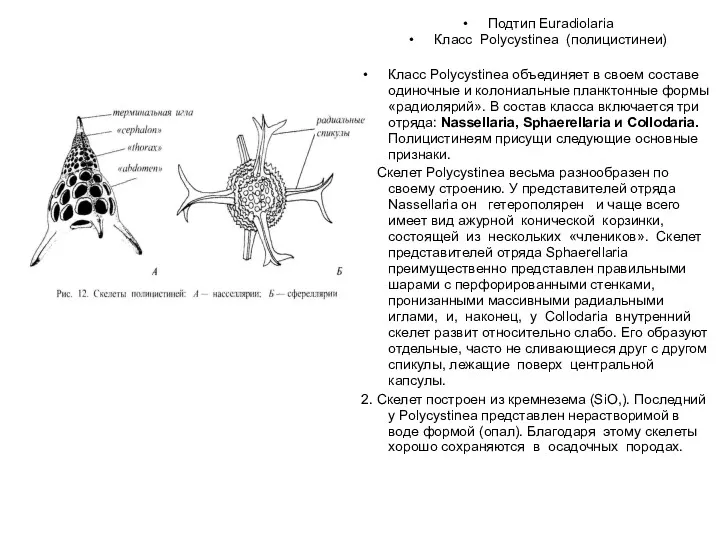
3. Полицистинеи обладают настоящей центральной капсулой. Последняя представляет собой замкнутое мешковидное образование, стенки

Класс Phaeodaria (феодарии)
Относительно крупные планктонные глубоководные организмы, в строении которых прослеживаются элементы двусторонней
Относительно крупные планктонные глубоководные организмы, в строении которых прослеживаются элементы двусторонней
4. Центральная капсула округлая или мешковидная, всегда
двустороннесимметрична. Ее стенка сплошная и построена из
Тип Heliozoa
Название «с о л н е ч н и к и» впервые
Тип Heliozoa
Название «с о л н е ч н и к и» впервые

Надтип Apicomplexa:
Тип Sporozoaе (Споровики): Класс Gregarinomorpha, Класс Coccidiomorphina, Отр. Сoccidiida, Отр. Haemosporidia, Тип
Надтип Apicomplexa: Тип Sporozoaе (Споровики): Класс Gregarinomorpha, Класс Coccidiomorphina, Отр. Сoccidiida, Отр. Haemosporidia, Тип

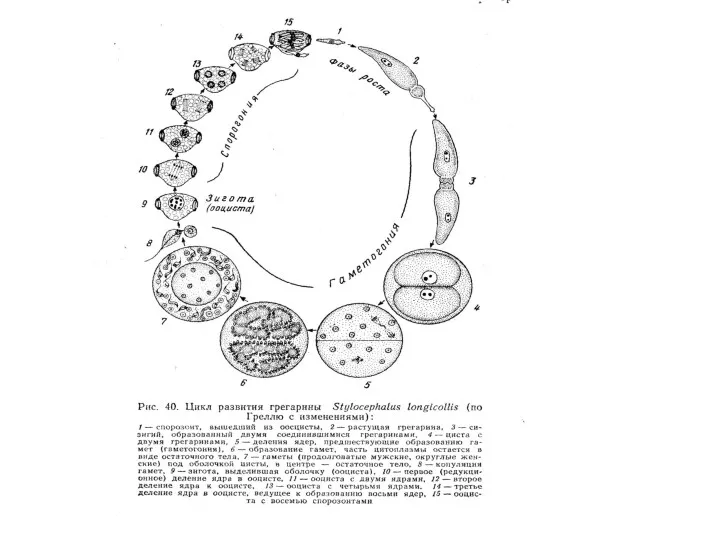
Тип Sporozoaе
Для споровиков характерен сложный жизненный цикл, который обычно включает три последовательные фазы.
Тип Sporozoaе
Для споровиков характерен сложный жизненный цикл, который обычно включает три последовательные фазы.

Главной отличительной особенностью Арiсоmplеха является то, что в их жизненном цикле обязательно присутствует
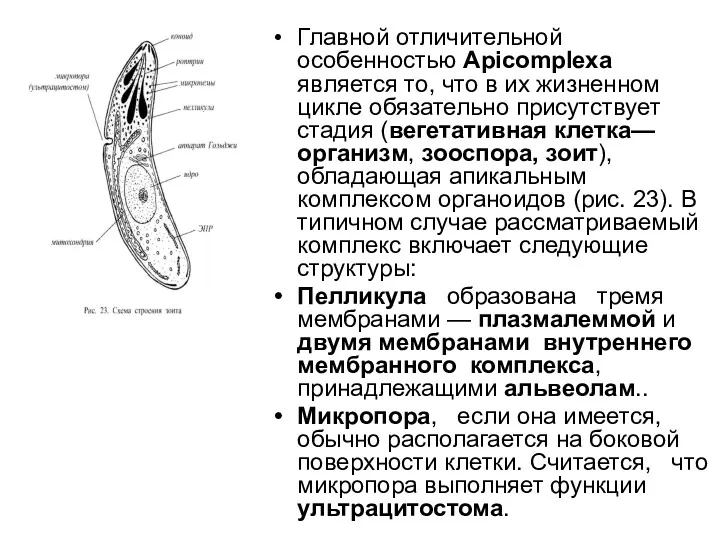
В передней половине клетки располагаются р о п т р и и. Это
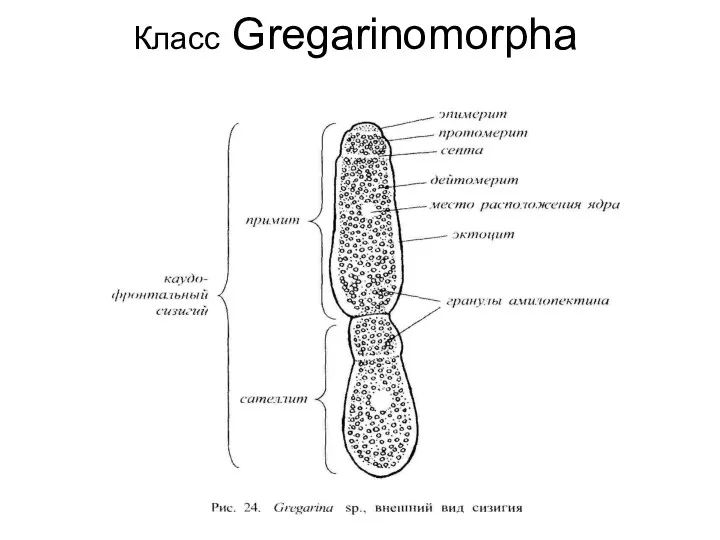
Класс Gregarinomorphа
В состав класса входят полостные и кишечные паразиты беспозвоночных и низших хордовых.
Класс Gregarinomorphа
В состав класса входят полостные и кишечные паразиты беспозвоночных и низших хордовых.

Класс Gregarinomorpha
Класс Gregarinomorpha
Класс Coccidiomorphina
В состав класса входят преимущественно внутриклеточные паразиты беспозвоночных и позвоночных животных. Главной
Класс Coccidiomorphina
В состав класса входят преимущественно внутриклеточные паразиты беспозвоночных и позвоночных животных. Главной

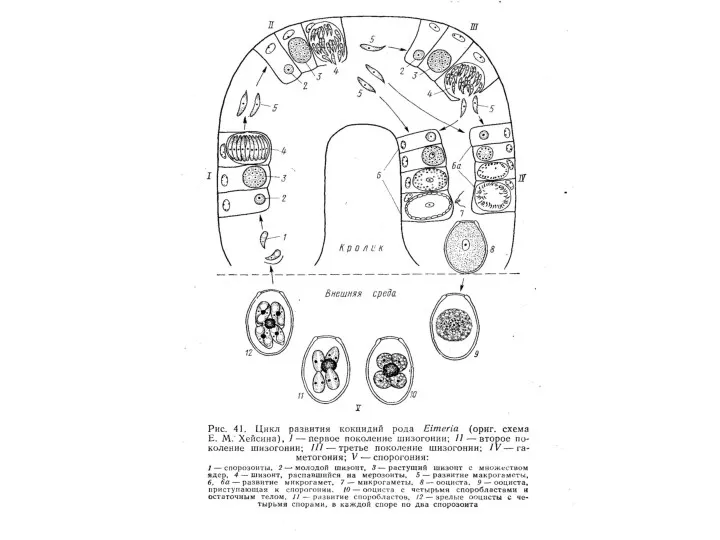
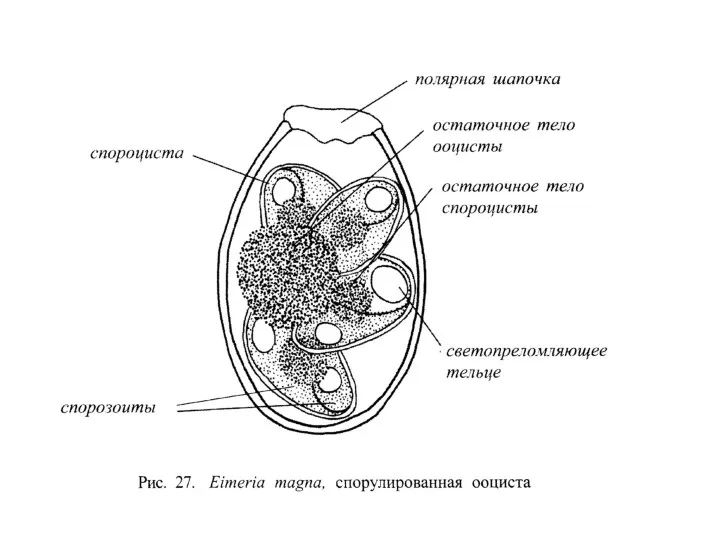
Отряд HAEMOSPORIDIA
К кровяным споровикам относятся представители типа Sporozoae, паразитирующие в кровеносном русле позвоночных
Отряд HAEMOSPORIDIA
К кровяным споровикам относятся представители типа Sporozoae, паразитирующие в кровеносном русле позвоночных

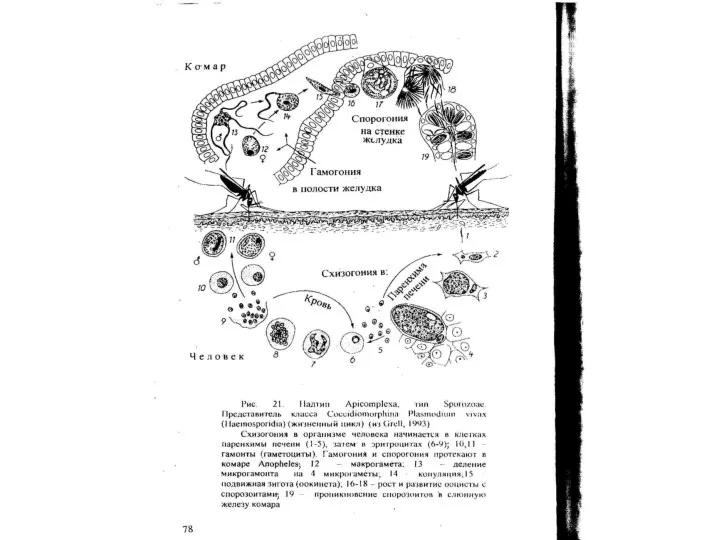
Надтип Chromophyta
Тип Slopalinata
Класс Opalinatea
Все представители класса - полиэнергидные простейшие. Число ядер у них
Тип Slopalinata
Класс Opalinatea
Все представители класса - полиэнергидные простейшие. Число ядер у них

 Высшая нервная деятельность (ВНД) человека
Высшая нервная деятельность (ВНД) человека Tissues. The histophysiology of the epithelial tissue
Tissues. The histophysiology of the epithelial tissue Культивирование вирусов
Культивирование вирусов Закономерности влияния окружающей среды на развитие фенотипа животных. (Лекция 1)
Закономерности влияния окружающей среды на развитие фенотипа животных. (Лекция 1) Обмен веществ и превращение энергии в клетке
Обмен веществ и превращение энергии в клетке Структура центра терморегуляции. Роль центра в регуляции теплообмена
Структура центра терморегуляции. Роль центра в регуляции теплообмена Основы селекции растений, животных, микроорганизмов
Основы селекции растений, животных, микроорганизмов Соцветия. Биологическое значение, классификация, происхождение соцветий
Соцветия. Биологическое значение, классификация, происхождение соцветий Сцепленное наследование признаков. Закон Томаса Моргана. (9 класс)
Сцепленное наследование признаков. Закон Томаса Моргана. (9 класс) Цитология. Методы фиксации и окрашивания клеток и тканей
Цитология. Методы фиксации и окрашивания клеток и тканей Bases of plants ecology
Bases of plants ecology Многолетние цветковые растения
Многолетние цветковые растения Пресмыкающиеся Нижегородской области
Пресмыкающиеся Нижегородской области Генная и клеточная инженерия
Генная и клеточная инженерия Понятие о гене. Биосинтез белка. Транскрипция
Понятие о гене. Биосинтез белка. Транскрипция Учение Ч. Дарвина
Учение Ч. Дарвина Современные методы секвенирования ДНК
Современные методы секвенирования ДНК Тип Хордовые. Подтип Бесчерепные (класс Ланцетник) и Черепные (класс Круглоротые)
Тип Хордовые. Подтип Бесчерепные (класс Ланцетник) и Черепные (класс Круглоротые) Скелет и мышцы
Скелет и мышцы Омыртқасыз және омыртқалы жануарлардың қозғалуы
Омыртқасыз және омыртқалы жануарлардың қозғалуы Человек, как результат биологической и социокультурной эволюции
Человек, как результат биологической и социокультурной эволюции Биотехнологии в сельском хозяйстве
Биотехнологии в сельском хозяйстве Царство грибов
Царство грибов Экологический праздник Синичкин день
Экологический праздник Синичкин день Мы - друзья пернатых
Мы - друзья пернатых Physiology of Bacteria
Physiology of Bacteria Занимательное путешествие в мир насекомых
Занимательное путешествие в мир насекомых Корень, побег, стебель и лист покрытосеменных
Корень, побег, стебель и лист покрытосеменных