- Субстратное фосфорилирование. Виды брожения

Содержание
- 2. Субстратное фосфорилирование — характерная для всех живых организмов реакция синтеза АТФ или ГТФ путём прямого переноса
- 3. Фосфорилирование – присоединение фосфатной группы к молекуле. Субстратное фосфорилирование — это синтез АТФ в процессе гликолиза,
- 5. Общая схема субстратного фосфорилирования.
- 6. Механизм После фосфорилирования промежуточного продукта, его фосфатная группа (неорганический фосфат) переносится на АДФ. Поскольку потенциал передачи
- 7. Реакция 1,3-дифосфоглицерата, содержащего макроэргическую связь в 1 положении, ферментом фосфоглицераткиназой на молекулу АДФ переносится остаток фосфорной
- 8. Броже́ние — биохимический процесс, основанный на окислительно-восстановительных превращениях органических соединений в анаэробных условиях. В ходе брожения
- 9. Брожение осуществляют многие микроорганизмы, так называемые бродильщики, как прокариотические, так и эукариотические, например, дрожжи рода Saccharomyces[en]
- 10. В качестве субстрата в процессах брожения могут выступать различные органические соединения, в которых углерод окислен не
- 11. Виды брожения Спиртовое брожение в 90 % случаев осуществляют дрожжи родов Saccharomyces и Schizosaccharomyces. Они сбраживают
- 13. Молочнокислое брожение осуществляют филогенетически неродственные организмы: представители порядков Lactobacillales, Bacillales а также семейства Bifidobacteriaceae. Эти бактерии
- 14. Пропионовокислое брожение осуществляют преимущественно бактерии подпорядка Propionibacterineae класса Actinobacteria, обитающие в рубце и кишечнике жвачных животных.
- 15. Муравьинокислое (также известно как смешанное) брожение осуществляют бактерии порядка Enterobacteriales, большинство из которых относится к кишечной
- 16. К маслянокислому брожению способны некоторые представители родов Clostridium, Butyrivibrio, Fusobacterium, Eubacterium. Клостридии в качестве субстрата для
- 18. Скачать презентацию

Субстратное фосфорилирование — характерная для всех живых организмов реакция синтеза АТФ или ГТФ путём прямого
Субстратное фосфорилирование — характерная для всех живых организмов реакция синтеза АТФ или ГТФ путём прямого

Фосфорилирование – присоединение фосфатной группы к молекуле.
Субстратное фосфорилирование — это синтез
Фосфорилирование – присоединение фосфатной группы к молекуле.
Субстратное фосфорилирование — это синтез

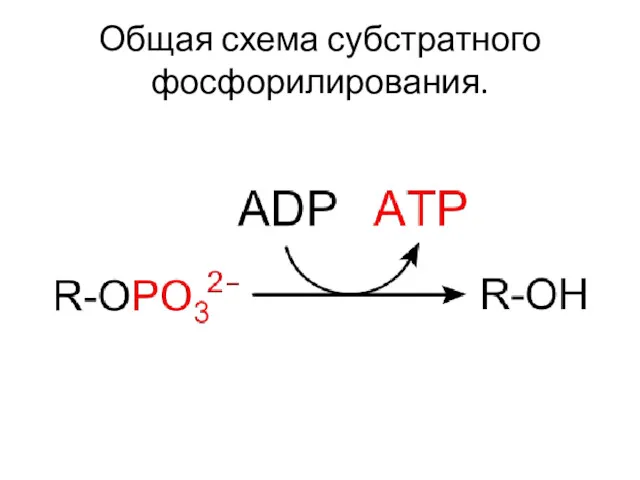
Общая схема субстратного фосфорилирования.
Общая схема субстратного фосфорилирования.

Механизм
После фосфорилирования промежуточного продукта, его фосфатная группа (неорганический фосфат) переносится на
Механизм
После фосфорилирования промежуточного продукта, его фосфатная группа (неорганический фосфат) переносится на
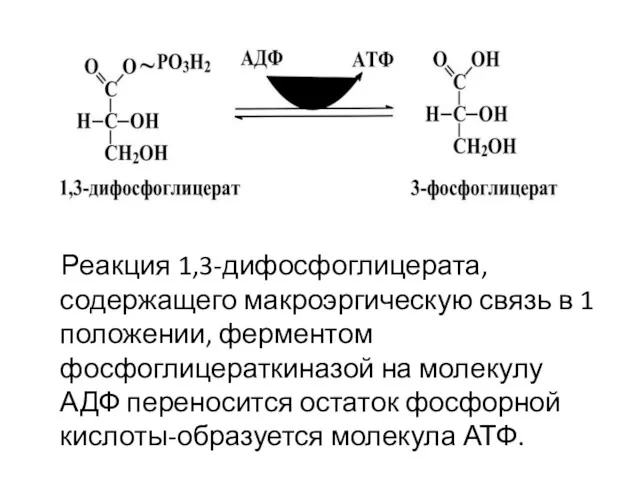
Реакция 1,3-дифосфоглицерата, содержащего макроэргическую связь в 1 положении, ферментом фосфоглицераткиназой
Реакция 1,3-дифосфоглицерата, содержащего макроэргическую связь в 1 положении, ферментом фосфоглицераткиназой

Броже́ние — биохимический процесс, основанный на окислительно-восстановительных превращениях органических соединений в
Броже́ние — биохимический процесс, основанный на окислительно-восстановительных превращениях органических соединений в

Брожение осуществляют многие микроорганизмы, так называемые бродильщики, как прокариотические, так и
Брожение осуществляют многие микроорганизмы, так называемые бродильщики, как прокариотические, так и

В качестве субстрата в процессах брожения могут выступать различные органические соединения,
В качестве субстрата в процессах брожения могут выступать различные органические соединения,

Виды брожения
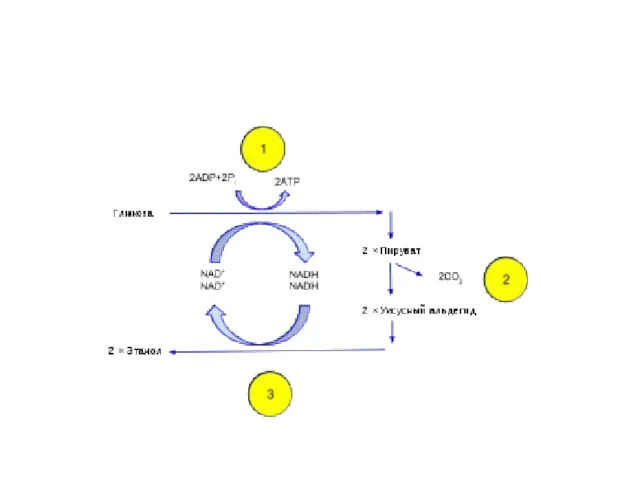
Спиртовое брожение в 90 % случаев осуществляют дрожжи родов Saccharomyces и Schizosaccharomyces. Они сбраживают моно- и дисахариды с образованием
Виды брожения
Спиртовое брожение в 90 % случаев осуществляют дрожжи родов Saccharomyces и Schizosaccharomyces. Они сбраживают моно- и дисахариды с образованием

Молочнокислое брожение осуществляют филогенетически неродственные организмы: представители порядков Lactobacillales, Bacillales а также семейства Bifidobacteriaceae. Эти бактерии живут
Молочнокислое брожение осуществляют филогенетически неродственные организмы: представители порядков Lactobacillales, Bacillales а также семейства Bifidobacteriaceae. Эти бактерии живут

Пропионовокислое брожение осуществляют преимущественно бактерии подпорядка Propionibacterineae класса Actinobacteria, обитающие в рубце и кишечнике жвачных животных.
Пропионовокислое брожение осуществляют преимущественно бактерии подпорядка Propionibacterineae класса Actinobacteria, обитающие в рубце и кишечнике жвачных животных.

Муравьинокислое (также известно как смешанное) брожение осуществляют бактерии порядка Enterobacteriales, большинство из
Муравьинокислое (также известно как смешанное) брожение осуществляют бактерии порядка Enterobacteriales, большинство из

К маслянокислому брожению способны некоторые представители родов Clostridium, Butyrivibrio, Fusobacterium, Eubacterium. Клостридии в качестве субстрата
К маслянокислому брожению способны некоторые представители родов Clostridium, Butyrivibrio, Fusobacterium, Eubacterium. Клостридии в качестве субстрата
 Витамины. Естественные и искусственные витамины
Витамины. Естественные и искусственные витамины Цитология. Клеточная теория. Строение клетки
Цитология. Клеточная теория. Строение клетки Курс подготовки к ЕГЭ по биологии
Курс подготовки к ЕГЭ по биологии Теории возникновения жизни на Земле
Теории возникновения жизни на Земле Деление клетки. Митоз
Деление клетки. Митоз Пора года - лето
Пора года - лето Тестирование по теме Цветок. Соцветие
Тестирование по теме Цветок. Соцветие Тип Плоские черви
Тип Плоские черви Насекомые. Отряд перепончатокрылые
Насекомые. Отряд перепончатокрылые Время посева и глубина заделки семян
Время посева и глубина заделки семян Ядовитые растения
Ядовитые растения Бассүйек кұрылысы, оның милық және беттік бөліктері
Бассүйек кұрылысы, оның милық және беттік бөліктері Митоз. Деление клеток организма
Митоз. Деление клеток организма Плоды. Классификация.
Плоды. Классификация. Введение в физиологию. Физиология клетки
Введение в физиологию. Физиология клетки Генетика. Решение задач
Генетика. Решение задач Сборник заданий № 25 с ответами. ЕГЭ по биологии
Сборник заданий № 25 с ответами. ЕГЭ по биологии Гидротермические условия для выращивания растений
Гидротермические условия для выращивания растений Основные популяционные показатели
Основные популяционные показатели Снежный барс (Ирбис)
Снежный барс (Ирбис) Дневные хищные, совы, куриные
Дневные хищные, совы, куриные Как появился человек на Земле
Как появился человек на Земле Лекарственные растения Мангистауской области
Лекарственные растения Мангистауской области Надкласс II. Рыбы
Надкласс II. Рыбы Cell Structure end Function
Cell Structure end Function Деревья
Деревья Medical Academy Named after S. I. Georgievsky of Crimea
Medical Academy Named after S. I. Georgievsky of Crimea Увеличительные приборы. 6 класс. Урок № 3
Увеличительные приборы. 6 класс. Урок № 3