- Температурные адаптации животных

Содержание
- 2. Химические адаптации Температурные условия в первую очередь влияют на скорость метаболизма, которая способна изменять величину теплопродукции.
- 3. Выживаемость животных при низких температурах среды определяется двумя факторами: теплоизолирующими свойствами покровных тканей и способностью животных
- 4. Афферентный поток, возникающий в результате возбуждения холодовых рецепторов (телец Краузе), через таламус и гипоталамус активизирует продукцию
- 6. Физические адаптации На уровень теплоотдачи влияет развитие теплозащитных покровов, особое устройстве кровеносной системы, распределение жировых запасов
- 7. Показательна и реакция сердечно-сосудистой системы на изменения температуры среды. Изменения частоты дыхания сопровождаются изменением частоты сердечных
- 8. Помимо сердечной мышцы реактивностью на термический фактор обладает и сосудистая система. У привычных к холоду животных
- 9. Параллельное и близкое расположение артерий и вен позволяет эффективно выводить из организма избыточное тепло. Артериальная кровь
- 10. Системы противоточного теплообмена, помогающие поддерживать постоянную температуру внутренних органов, обнаружены в лапах и хвостах у сумчатых,
- 11. У крупного рогатого скота и близких к нему видов животных противоточный механизм кровообращения имеется в межреберных
- 12. Жировая ткань разновидность соединительной ткани животных организмов, образующаяся из мезенхимы и состоящая из жировых клеток —
- 14. Белая жировая ткань - представлена крупными округлыми клетками с большой светлой каплей нейтрального жира. Размеры клеток
- 15. Бурая жировая ткань — один из двух видов жировой ткани у млекопитающих. Хорошо развита у новорожденных
- 16. Среди животных бурая жировая ткань лучше всего развита у тех, которые зимой впадают в спячку. Во
- 17. У морских млекопитающих – ластоногих и китов – слой подкожной жировой клетчатки распределен по всему телу.
- 18. У человека бурая жировая ткань хорошо развита только у новорожденных (примерно 5 % от массы тела)
- 19. Теплозащитные покровы сохранение тепла за счет физической терморегуляции не требует дополнительных пищевых ресурсов, оно является основным
- 21. Поведенческие адаптации Поведенческие способы регуляции теплообмена для теплокровных животных не менее важны, чем для пойкилотермных, и
- 23. В ряде случаев гомойотермные животные используют в целях терморегуляции групповое поведение. Например, некоторые пингвины в сильный
- 24. Правило Баргмана в пределах вида или достаточно однородной группы близких видов животные (теплокровные) с более крупными
- 25. Мелвильский островной волк (арктических островов в Северной Америке и в северной части Гренландии) Длина средней особи
- 26. Аравийский волк (живёт на ограниченных ареалах в южном Израиле, Омане, Йемене, Иордании, Саудовской Аравии и, возможно,
- 27. Человек также подчиняется правилу Бергмана.
- 28. Правило Аллена Согласно с этим правилом среди родственных форм гомойотермных (теплокровных) животных, ведущих сходный образ жизни,
- 29. Примером данного правила являются представители семейства Собачьи из различных регионов. Наименьшие (относительно длины тела) уши и
- 30. Правило Глогера состоит в том, что среди родственных друг другу форм (разных рас или подвидов одного
- 32. Скачать презентацию

Химические адаптации
Температурные условия в первую очередь влияют на скорость метаболизма, которая
Химические адаптации
Температурные условия в первую очередь влияют на скорость метаболизма, которая

Выживаемость животных при низких температурах среды определяется двумя факторами: теплоизолирующими свойствами
Выживаемость животных при низких температурах среды определяется двумя факторами: теплоизолирующими свойствами

Афферентный поток, возникающий в результате возбуждения холодовых рецепторов (телец Краузе), через
Афферентный поток, возникающий в результате возбуждения холодовых рецепторов (телец Краузе), через
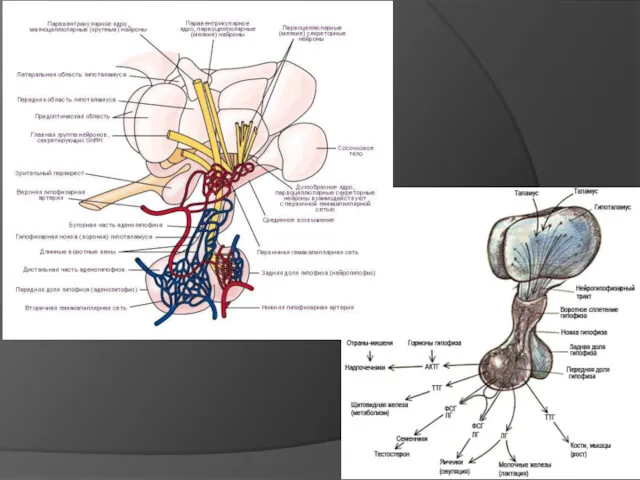

Физические адаптации
На уровень теплоотдачи влияет развитие теплозащитных покровов, особое устройстве кровеносной
Физические адаптации
На уровень теплоотдачи влияет развитие теплозащитных покровов, особое устройстве кровеносной

Показательна и реакция сердечно-сосудистой системы на изменения температуры среды. Изменения частоты
Показательна и реакция сердечно-сосудистой системы на изменения температуры среды. Изменения частоты

Помимо сердечной мышцы реактивностью на термический фактор обладает и сосудистая система.
Помимо сердечной мышцы реактивностью на термический фактор обладает и сосудистая система.
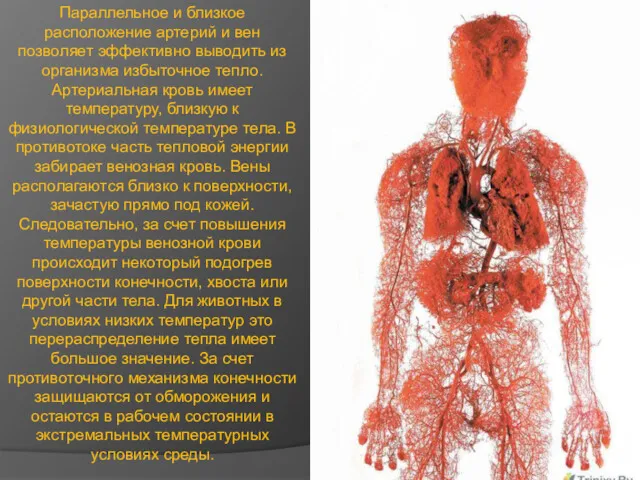
Параллельное и близкое расположение артерий и вен позволяет эффективно выводить из
Параллельное и близкое расположение артерий и вен позволяет эффективно выводить из

Системы противоточного теплообмена, помогающие поддерживать постоянную температуру внутренних органов, обнаружены в
Системы противоточного теплообмена, помогающие поддерживать постоянную температуру внутренних органов, обнаружены в

У крупного рогатого скота и близких к нему видов животных противоточный
У крупного рогатого скота и близких к нему видов животных противоточный
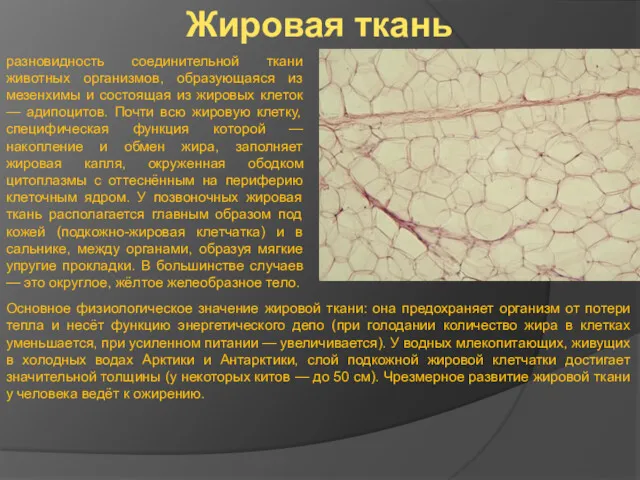
Жировая ткань
разновидность соединительной ткани животных организмов, образующаяся из мезенхимы и состоящая
Жировая ткань
разновидность соединительной ткани животных организмов, образующаяся из мезенхимы и состоящая
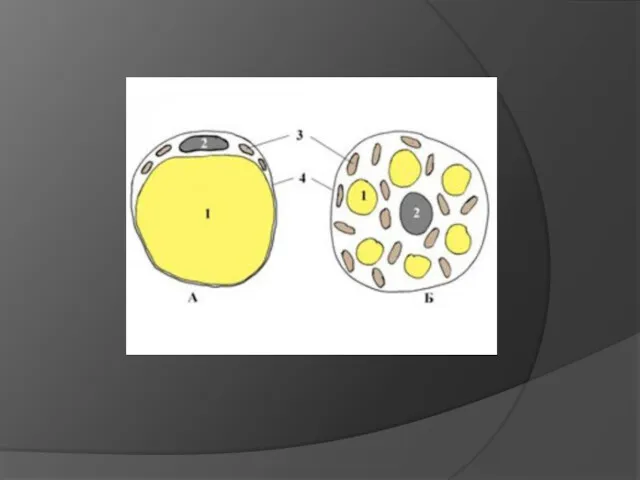
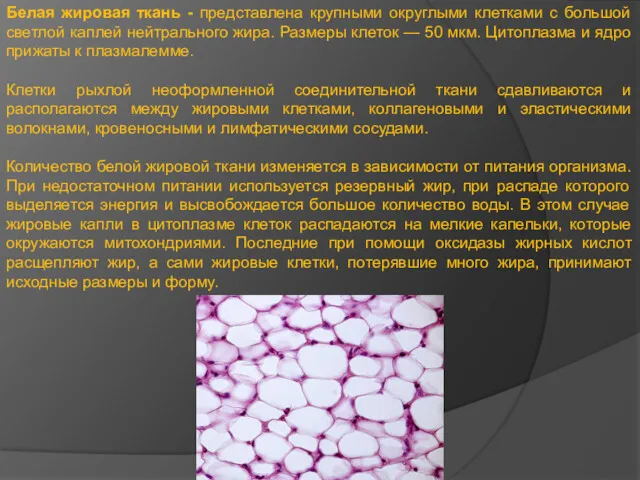
Белая жировая ткань - представлена крупными округлыми клетками с большой светлой
Белая жировая ткань - представлена крупными округлыми клетками с большой светлой

Бурая жировая ткань — один из двух видов жировой ткани у
Бурая жировая ткань — один из двух видов жировой ткани у

Среди животных бурая жировая ткань лучше всего развита у тех, которые
Среди животных бурая жировая ткань лучше всего развита у тех, которые

У морских млекопитающих – ластоногих и китов – слой подкожной жировой
У морских млекопитающих – ластоногих и китов – слой подкожной жировой
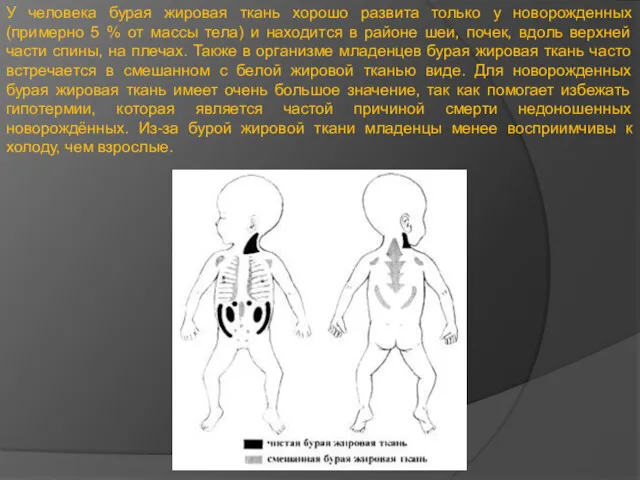
У человека бурая жировая ткань хорошо развита только у новорожденных (примерно
У человека бурая жировая ткань хорошо развита только у новорожденных (примерно

Теплозащитные покровы
сохранение тепла за счет физической терморегуляции не требует дополнительных пищевых
Теплозащитные покровы
сохранение тепла за счет физической терморегуляции не требует дополнительных пищевых


Поведенческие адаптации
Поведенческие способы регуляции теплообмена для теплокровных животных не менее важны,
Поведенческие адаптации
Поведенческие способы регуляции теплообмена для теплокровных животных не менее важны,


В ряде случаев гомойотермные животные используют в целях терморегуляции групповое поведение.
В ряде случаев гомойотермные животные используют в целях терморегуляции групповое поведение.

Правило Баргмана
в пределах вида или достаточно однородной группы близких видов животные
Правило Баргмана
в пределах вида или достаточно однородной группы близких видов животные

Мелвильский островной волк
(арктических островов в Северной Америке и в северной части
Мелвильский островной волк
(арктических островов в Северной Америке и в северной части

Аравийский волк
(живёт на ограниченных ареалах в южном Израиле, Омане, Йемене, Иордании,
Аравийский волк
(живёт на ограниченных ареалах в южном Израиле, Омане, Йемене, Иордании,
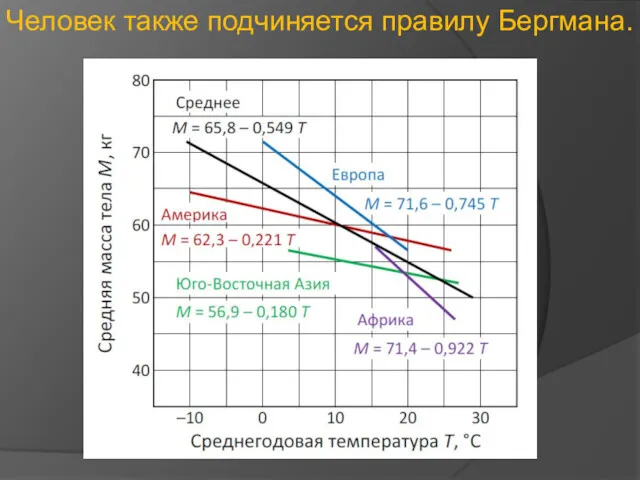
Человек также подчиняется правилу Бергмана.
Человек также подчиняется правилу Бергмана.

Правило Аллена
Согласно с этим правилом среди родственных форм гомойотермных (теплокровных) животных,
Правило Аллена
Согласно с этим правилом среди родственных форм гомойотермных (теплокровных) животных,

Примером данного правила являются представители семейства Собачьи из различных регионов. Наименьшие
Примером данного правила являются представители семейства Собачьи из различных регионов. Наименьшие

Правило Глогера
состоит в том, что среди родственных друг другу форм (разных
Правило Глогера
состоит в том, что среди родственных друг другу форм (разных
 Красная книга. Коала
Красная книга. Коала Самые необычные животные планеты
Самые необычные животные планеты Индивидуальное развитие организмов (онтогенез)
Индивидуальное развитие организмов (онтогенез) Викопні рослини
Викопні рослини Ріст та розвиток вегетативних органів рослин
Ріст та розвиток вегетативних органів рослин Презентация по биологии на тему Класс Пресмыкающиеся. Особенности внешнего и внутреннего строения. Многообразие. Происхождение
Презентация по биологии на тему Класс Пресмыкающиеся. Особенности внешнего и внутреннего строения. Многообразие. Происхождение Репликация. Эукариоты. Часть 2
Репликация. Эукариоты. Часть 2 Антропология, как комплексное учение о человеке. Место человека в животном мире и этапы антропогенеза
Антропология, как комплексное учение о человеке. Место человека в животном мире и этапы антропогенеза Развитие ребёнка 7-9 месяцев
Развитие ребёнка 7-9 месяцев Биология, как наука
Биология, как наука Презентация к уроку Основные свойства жизни
Презентация к уроку Основные свойства жизни Общие вопросы анатомии и физиологии сердечно-сосудистой системы
Общие вопросы анатомии и физиологии сердечно-сосудистой системы Презентация опыта
Презентация опыта Учебно-методический комплекс дисциплины Методы электрофоретического разделения белков
Учебно-методический комплекс дисциплины Методы электрофоретического разделения белков Все о пище с точки зрения химика
Все о пище с точки зрения химика Индивидуальное развитие организма. Индивидуальное развитие человека
Индивидуальное развитие организма. Индивидуальное развитие человека Презентация к уроку Эволюционное учение Ч. Дарвина
Презентация к уроку Эволюционное учение Ч. Дарвина Крапива - удивительное растение
Крапива - удивительное растение ПБ_Отр_Вороб_синицы 2019
ПБ_Отр_Вороб_синицы 2019 Презентация по теме ''Лесные пожары''
Презентация по теме ''Лесные пожары'' Взаимодействие генов
Взаимодействие генов Вимерлі тварини: найбільші динозаври
Вимерлі тварини: найбільші динозаври Что ни страница - то слон, то львица
Что ни страница - то слон, то львица Патология клеток. Морфология лейкоцитов: нейтрофилы
Патология клеток. Морфология лейкоцитов: нейтрофилы дикий интерактив конец
дикий интерактив конец Берёза
Берёза Кое-что о птицах - презентации.
Кое-что о птицах - презентации. Полюс недоступности. Органический мир
Полюс недоступности. Органический мир