- Токсины бактерий. Тема 4

Содержание
- 2. Токсины по механизму действия подразделяют на 5 типов: Повреждающие клеточные мембраны (порообразующие токсины). К ним относят
- 3. 4. Активаторы иммунного ответа. Отдельные бактериальные токсины могут действовать непосредственно на Т-клетки и антигенпрезентирующие клетки иммунной
- 4. Одними из первых охарактеризованных токсинов, продуцируемых бактериями E. coli, были колицины. В 1925 г. А. Грация
- 5. Бактериоцины – это вещества белковой природы или представленные белком в комплексе с липополисахаридами, (за антибактериальную активность
- 6. По механизму действия на бактериальную клетку бактериоцины подразделяют на 4 основные группы: Ингибирующие окислительное фосфорилирование в
- 7. Грамположительные бактерии продуцируют: Бактериоцины класса I (низин и др) ингибируют синтез пептидогликана, связываясь с липидом II,
- 8. Грамотрицательные бактерии продуцируют микроцины, которые ингибируют ДНК-гиразу (MccB17), РНК-полимеразу (MccJ25) и аспартил-тРНК-синтетазу (MccC7-C51)
- 9. Существует два типа колициногенных плазмид, различающихся по размеру, количеству копий в клетке и способности к переносу
- 10. Различные плазмиды могут кодировать сходные колициновые белки. Опероны колицина включают кластер генов, состоящий из двух-трех генов:
- 11. Колицины не вырабатываются в бактериальных клетках в нормальных условиях, но в клетке постоянно присутствует ограниченное количество
- 12. Хорошо изученными являются токсины продуцируемые бактериями Bacillus thuringiensis (ВТ), которые приводят к гибели насекомых. Как правило,
- 13. Бактерии Bacillus thuringiensis продуцируют: α-экзотоксин (фосфолипаза С), является продуктом растущих клеток; предполагают, что эффект данного токсина,
- 14. Клетка Bacillus thuringiensis subsp. israelensis после споруляции (А) и её типичное параспоральное тельце с указанием взаиморасположения
- 15. Электронномикроскопическая фоторнафия B. thuringiensis subsp. thuringiensis berliner 1715 в процессе споруляции. Черным обозначено пароспоральное включение, представляющее
- 16. Группы Cry- и Cyt-белков, выделенные на основании сходства их аминокислотных последовательностей
- 17. Белки Cry состоят из трех доменов: Домен I состоит из 6 α-спиральных белковых молекул, имеющих амфипатическую
- 18. Трехмерная структура Cry1Ab белка
- 19. Кристаллическя структура белков Cry1Aa, Cry2Aa, Cry3Aa и and Cry4Ba Домен I обозначен красным цветом Домен II
- 20. Белки Cyt включают один домен Белки cyt-семейства, в отличие от cry-токсинов, способны вызывать лизис многих типов
- 21. Процесс действия δ-эндотоксинов Cry-семейства включает несколько этапов: Этап 1. Растворение кристаллических включений, содержащих молекулы протоксина, в
- 22. Этап 3. Конформационное изменение молекулы токсина, индуцированное контактом с первым специфическим рецептором, приводит к отщеплению небольшого
- 23. Схема действия δ-эндотоксина на клетку-мишень на примере Cry1А токсина по модели Bravo (По Soberon, Bravo, 2008)
- 24. Пути возникновения устойчивости насекомых к δ-эндотоксинам. На активность инсектицидных токсинов BT могут оказывать действие протеазы кишечника
- 25. Всего предложено три модели механизма цитоцидного действия δ-эндотоксинов на клетки кишечного эпителия насекомых, которые являются не
- 26. Помимо рассмотренных механизмов действия δ-эндотоксинов установлено, что они способны: в 2-5 раз снижать сопротивление искусственных фосфолипидных
- 27. Токсины семейств Vip и Cyt также разрушают клетки по механизму осмотического лизиса, формируя в мембране поры,
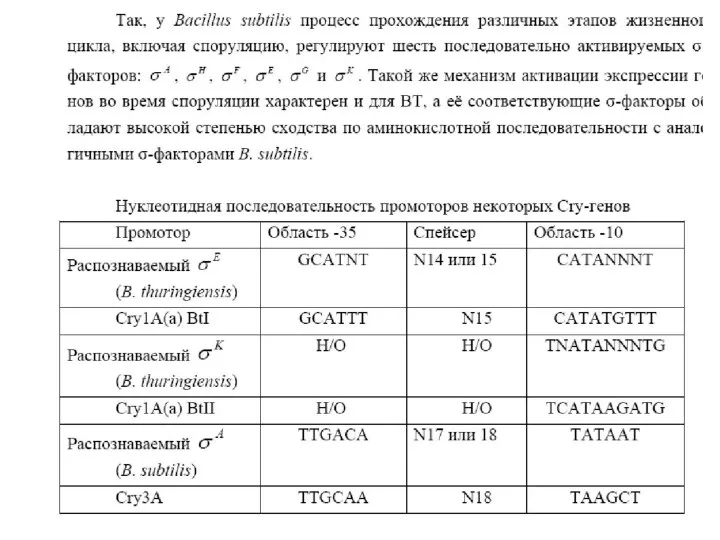
- 28. Многообразие групп токсинов, а также отдельных токсинов является следствием высокой генетической пластичности Bacillus thuringiensis. Большинство генов,
- 29. Размер генома различных штаммов ВТ варьирует от 2,4 до 5,7 миллионов п.н. Для большинства штаммов ВТ
- 32. Биотехнологическое использование бактерий Bacillus thuringiensis 1) использование различных штаммов ВТ в качестве источника генетических детерминант токсинов
- 33. Трансгенные растения, способные экспрессировать Cry-гены, обладают высокой степенью устойчивости к своим специфическим вредителям, за исключением случаев
- 34. Трансгенные бактерии используют в качестве биологического средства борьбы с различными вредителями сельского хозяйства и переносчиками определённых
- 35. Повышение инсектицидной активности штаммов ВТ может быть достигнуто путём создания рекомбинантных бактерий, Cry- и Cyt-гены которых
- 36. Недостатком непосредственного использования клеток ВТ после споруляции в качестве биопестицида или их споро-кристаллических комплексов является засорение
- 37. Генно-инженерные манипуляции, обеспечивающие направленное изменения определённых последовательностей генов, детерминирующих токсины, могут изменять свойства продукта этих генов.
- 38. Сферы использования Bt и генов, детерминирующих синтез токсинов: Создание и использование рекомбинантных штаммов бактерий для защиты
- 40. Скачать презентацию

Токсины по механизму действия подразделяют на 5 типов:
Повреждающие клеточные мембраны (порообразующие
Токсины по механизму действия подразделяют на 5 типов:
Повреждающие клеточные мембраны (порообразующие

4. Активаторы иммунного ответа. Отдельные бактериальные токсины могут действовать непосредственно на
4. Активаторы иммунного ответа. Отдельные бактериальные токсины могут действовать непосредственно на

Одними из первых охарактеризованных токсинов, продуцируемых бактериями E. coli, были колицины.
Одними из первых охарактеризованных токсинов, продуцируемых бактериями E. coli, были колицины.

Бактериоцины – это вещества белковой природы или представленные белком в комплексе
Бактериоцины – это вещества белковой природы или представленные белком в комплексе

По механизму действия на бактериальную клетку бактериоцины подразделяют на 4 основные
По механизму действия на бактериальную клетку бактериоцины подразделяют на 4 основные
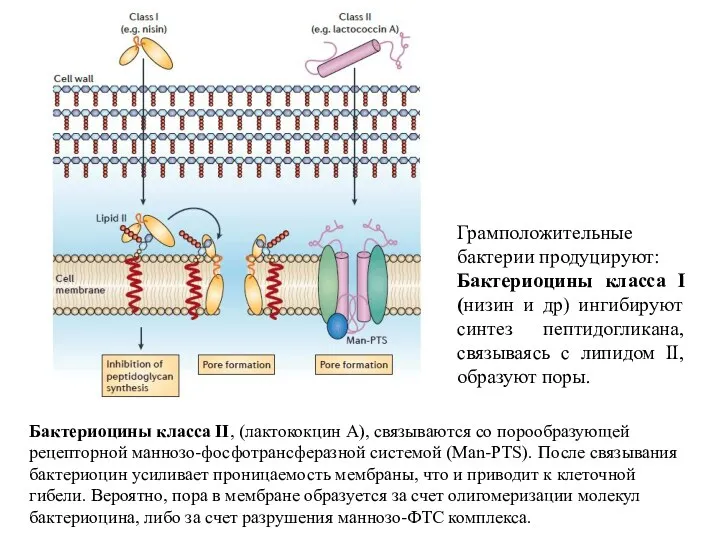
Грамположительные бактерии продуцируют:
Бактериоцины класса I (низин и др) ингибируют синтез пептидогликана,
Грамположительные бактерии продуцируют:
Бактериоцины класса I (низин и др) ингибируют синтез пептидогликана,
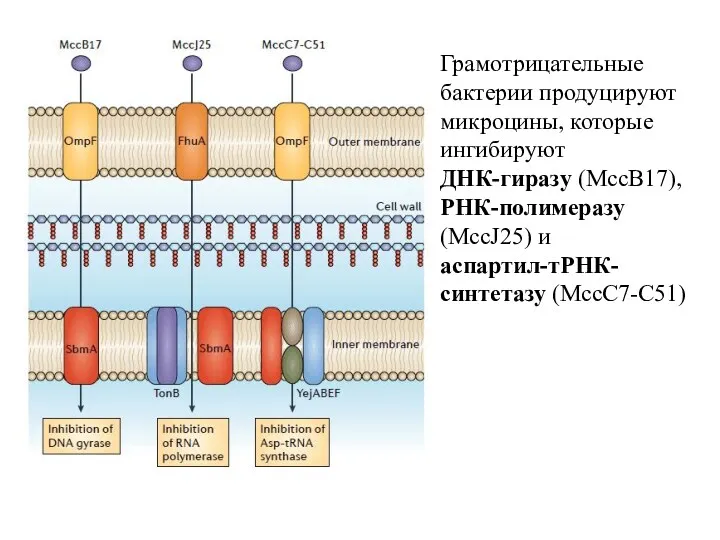
Грамотрицательные бактерии продуцируют микроцины, которые ингибируют
ДНК-гиразу (MccB17), РНК-полимеразу (MccJ25) и
Грамотрицательные бактерии продуцируют микроцины, которые ингибируют
ДНК-гиразу (MccB17), РНК-полимеразу (MccJ25) и

Существует два типа колициногенных плазмид, различающихся по размеру, количеству копий в
Существует два типа колициногенных плазмид, различающихся по размеру, количеству копий в

Различные плазмиды могут кодировать сходные колициновые белки.
Опероны колицина включают кластер генов,
Различные плазмиды могут кодировать сходные колициновые белки.
Опероны колицина включают кластер генов,

Колицины не вырабатываются в бактериальных клетках в нормальных условиях, но в
Колицины не вырабатываются в бактериальных клетках в нормальных условиях, но в

Хорошо изученными являются токсины продуцируемые бактериями Bacillus thuringiensis (ВТ), которые приводят
Хорошо изученными являются токсины продуцируемые бактериями Bacillus thuringiensis (ВТ), которые приводят

Бактерии Bacillus thuringiensis продуцируют:
α-экзотоксин (фосфолипаза С), является продуктом растущих клеток;
Бактерии Bacillus thuringiensis продуцируют:
α-экзотоксин (фосфолипаза С), является продуктом растущих клеток;
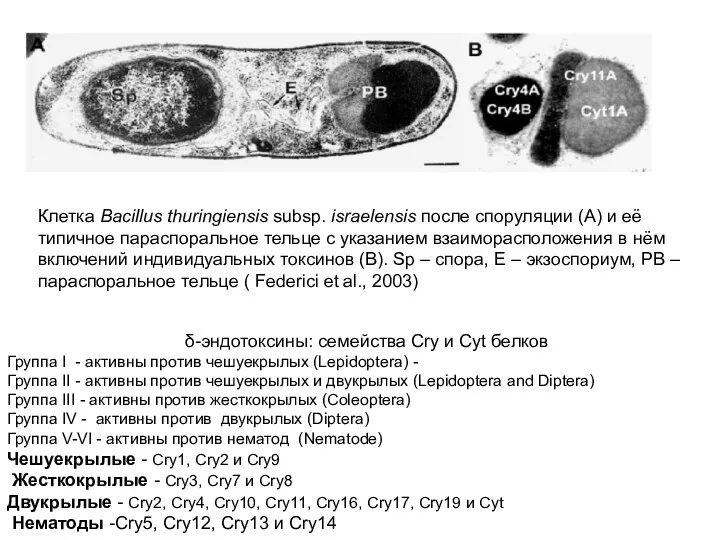
Клетка Bacillus thuringiensis subsp. israelensis после споруляции (А) и её типичное
Клетка Bacillus thuringiensis subsp. israelensis после споруляции (А) и её типичное
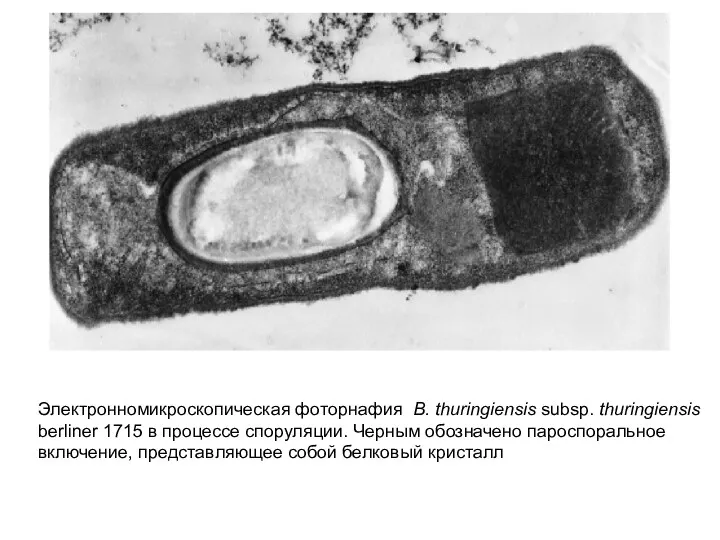
Электронномикроскопическая фоторнафия B. thuringiensis subsp. thuringiensis berliner 1715 в процессе споруляции.
Электронномикроскопическая фоторнафия B. thuringiensis subsp. thuringiensis berliner 1715 в процессе споруляции.

Группы Cry- и Cyt-белков, выделенные на основании сходства их
аминокислотных последовательностей
Группы Cry- и Cyt-белков, выделенные на основании сходства их
аминокислотных последовательностей

Белки Cry состоят из трех доменов:
Домен I состоит из 6 α-спиральных
Белки Cry состоят из трех доменов:
Домен I состоит из 6 α-спиральных
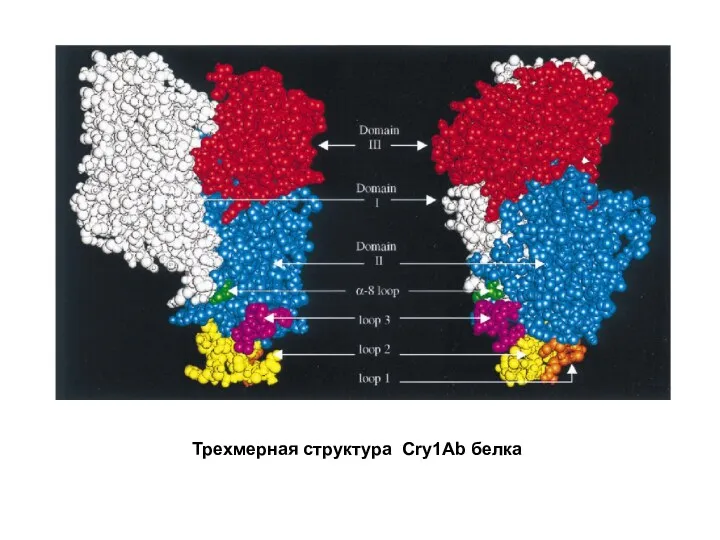
Трехмерная структура Cry1Ab белка
Трехмерная структура Cry1Ab белка
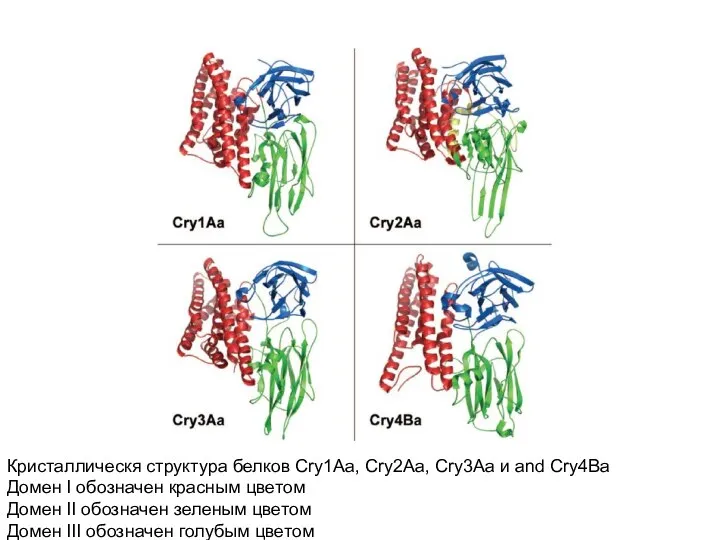
Кристаллическя структура белков Cry1Aa, Cry2Aa, Cry3Aa и and Cry4Ba
Домен I обозначен
Кристаллическя структура белков Cry1Aa, Cry2Aa, Cry3Aa и and Cry4Ba
Домен I обозначен

Белки Cyt включают один домен
Белки cyt-семейства, в отличие от cry-токсинов, способны
Белки Cyt включают один домен
Белки cyt-семейства, в отличие от cry-токсинов, способны

Процесс действия δ-эндотоксинов Cry-семейства включает несколько этапов:
Этап 1. Растворение кристаллических включений,
Процесс действия δ-эндотоксинов Cry-семейства включает несколько этапов:
Этап 1. Растворение кристаллических включений,

Этап 3. Конформационное изменение молекулы токсина, индуцированное контактом с первым специфическим
Этап 3. Конформационное изменение молекулы токсина, индуцированное контактом с первым специфическим
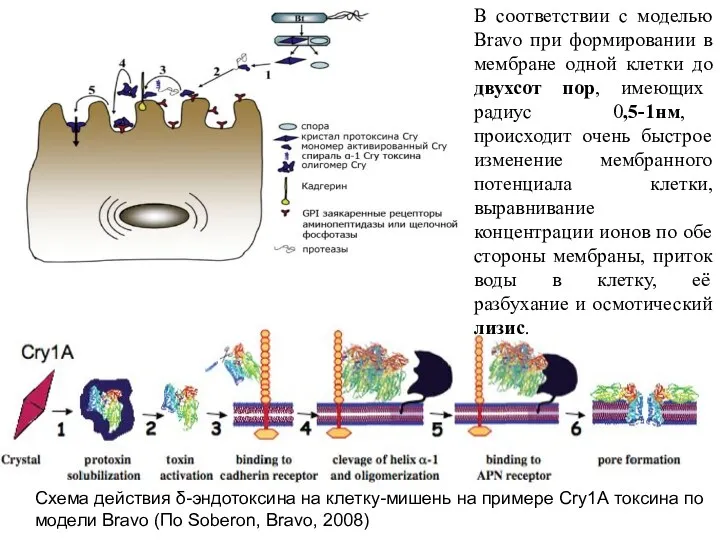
Схема действия δ-эндотоксина на клетку-мишень на примере Cry1А токсина по модели
Схема действия δ-эндотоксина на клетку-мишень на примере Cry1А токсина по модели

Пути возникновения устойчивости насекомых к δ-эндотоксинам.
На активность инсектицидных токсинов BT
Пути возникновения устойчивости насекомых к δ-эндотоксинам.
На активность инсектицидных токсинов BT

Всего предложено три модели механизма цитоцидного действия δ-эндотоксинов на клетки кишечного
Всего предложено три модели механизма цитоцидного действия δ-эндотоксинов на клетки кишечного

Помимо рассмотренных механизмов действия δ-эндотоксинов установлено, что они способны:
в 2-5 раз
Помимо рассмотренных механизмов действия δ-эндотоксинов установлено, что они способны:
в 2-5 раз

Токсины семейств Vip и Cyt также разрушают клетки по механизму осмотического
Токсины семейств Vip и Cyt также разрушают клетки по механизму осмотического

Многообразие групп токсинов, а также отдельных токсинов является следствием высокой генетической
Многообразие групп токсинов, а также отдельных токсинов является следствием высокой генетической
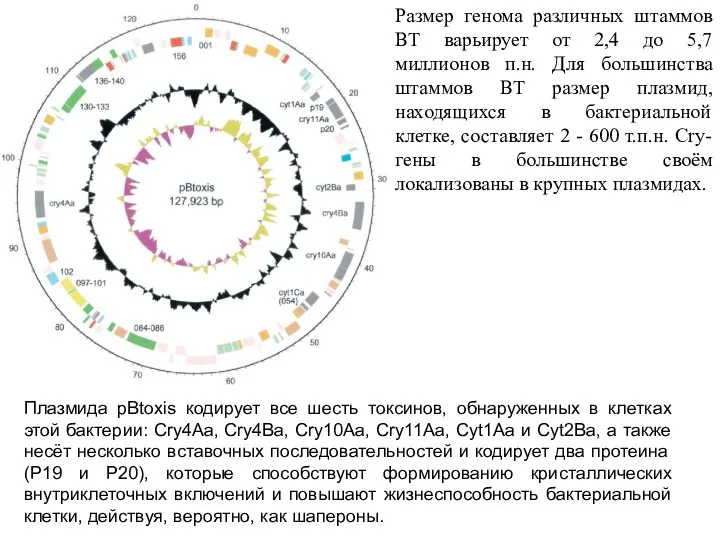
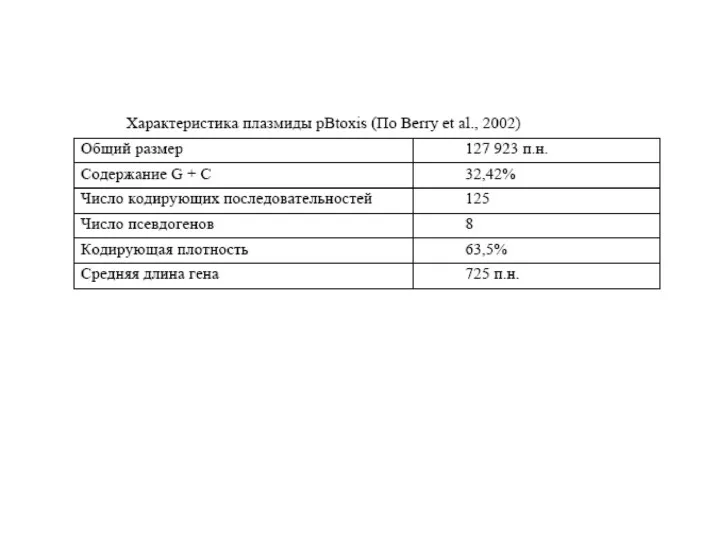
Размер генома различных штаммов ВТ варьирует от 2,4 до 5,7 миллионов
Размер генома различных штаммов ВТ варьирует от 2,4 до 5,7 миллионов

Биотехнологическое использование бактерий Bacillus thuringiensis
1) использование различных штаммов ВТ в
Биотехнологическое использование бактерий Bacillus thuringiensis
1) использование различных штаммов ВТ в

Трансгенные растения, способные экспрессировать Cry-гены, обладают высокой степенью устойчивости к своим
Трансгенные растения, способные экспрессировать Cry-гены, обладают высокой степенью устойчивости к своим

Трансгенные бактерии используют в качестве биологического средства борьбы с различными вредителями
Трансгенные бактерии используют в качестве биологического средства борьбы с различными вредителями

Повышение инсектицидной активности штаммов ВТ может быть достигнуто путём создания рекомбинантных
Повышение инсектицидной активности штаммов ВТ может быть достигнуто путём создания рекомбинантных

Недостатком непосредственного использования клеток ВТ после споруляции в качестве биопестицида или
Недостатком непосредственного использования клеток ВТ после споруляции в качестве биопестицида или

Генно-инженерные манипуляции, обеспечивающие направленное изменения определённых последовательностей генов, детерминирующих токсины, могут
Генно-инженерные манипуляции, обеспечивающие направленное изменения определённых последовательностей генов, детерминирующих токсины, могут

Сферы использования Bt и генов, детерминирующих синтез токсинов:
Создание и использование
Сферы использования Bt и генов, детерминирующих синтез токсинов:
Создание и использование
 Водоёмы по происхождению
Водоёмы по происхождению Поле и его обитатели. (3 класс)
Поле и его обитатели. (3 класс) Гигантская сколопендра
Гигантская сколопендра Значення риб. Рибництво
Значення риб. Рибництво Развитие жизни на Земле
Развитие жизни на Земле Микробиология - наука о микроорганизмах. История становления
Микробиология - наука о микроорганизмах. История становления Процессы жизнедеятельности растений
Процессы жизнедеятельности растений Полезный мед - натуральный мед
Полезный мед - натуральный мед Кровообращение. Строение и функции сердца
Кровообращение. Строение и функции сердца Размножение. Начало новой жизни
Размножение. Начало новой жизни Филогенетическое разнообразие грибоподобных организмов. Разнообразие. Систематика. Жизненные циклы
Филогенетическое разнообразие грибоподобных организмов. Разнообразие. Систематика. Жизненные циклы Обобщающий урок по теме Питание живых организмов по учебнику Природоведение 5 класс. Авторы Т.С.Сухова, В.И.Строганов
Обобщающий урок по теме Питание живых организмов по учебнику Природоведение 5 класс. Авторы Т.С.Сухова, В.И.Строганов Стати собак
Стати собак Тип Кольчатые черви
Тип Кольчатые черви Дельфиндер
Дельфиндер Тип Моллюски
Тип Моллюски Высшие споровые растения. Отдел Моховидные
Высшие споровые растения. Отдел Моховидные Сообщество, биоценоз, биогеоценоз , экосистема, биотоп
Сообщество, биоценоз, биогеоценоз , экосистема, биотоп Молекулярные основы наследственности
Молекулярные основы наследственности Биологические ритмы ,часы
Биологические ритмы ,часы Обобщающий урок по теме Цитология (презентация)
Обобщающий урок по теме Цитология (презентация) Пищеварительная система
Пищеварительная система Основы селекции организмов
Основы селекции организмов Бүйрек физиологиясы. Несеп түзілуінің механизмі
Бүйрек физиологиясы. Несеп түзілуінің механизмі Cистема кровообращения
Cистема кровообращения Презентация к игре Биологическая викторина
Презентация к игре Биологическая викторина Белки. Структура белков. Физические и химические свойства
Белки. Структура белков. Физические и химические свойства Введение в микробиологию
Введение в микробиологию