- Трансляция. Генетикалық код

Содержание
- 2. мРНҚ тізбегі үш нуклеотид бойынша оқылады Ақпараттың РНҚ формасынан белок формасына қайта өзгеруі дегеніміз «тасымал» немесе
- 3. мРНҚ молекуласындағы қатар біртіндеп үш нуклеотидтен тұратын топ арқылы оқылады. РНҚ төрт нуклеотидтен тұратын сызықты полимер
- 4. Генетикалық код
- 5. Бұл генетикалық код барлық заманауи ағзаларда бірегей қолданылады. Алайда кодтан бірнеше аздаған айырмашылықтар табылған, олар митохондрияның
- 6. Белок синтезі кезінде мүмкін болатын үш оқу аймағы
- 7. Жасушада белок синтезінің кезеңіне басты шек қоятын трансляция инициациясы – бұл фазада старттық кодонды іздестіру және
- 8. Эукариоттық мРНК- ның көбісі ұқсас құрылысты болып келеді: оларда 5’-соңында кэп-құрылымы болады («инвертирленген » және метилденген
- 9. Трансляция инициациясы процесс ін сипаттау үшін өткен ғасырдың 80-жж аяғында « сканирлейтін модель» ұсынған болатын. Сол
- 10. 40S суббірлікке инициаторлы мРНК қосылуы. Рибосомальді прединициаторлы кешеннің түзілуі. Трансляция инициациясында маңызды рөл инициаторлы тРНК (тРНКiМет)
- 11. Амин қышқылының активациясы
- 12. Аминоацил тРНҚ кешеніндегі байланыс құрылымы
- 14. мРНК –ң трансляция инициациясына дайындалуы Эукариот жасушасында бос мРНК, әдетте РНК байланыстыратын белоктармен бірге болып, рибонуклеопротеид
- 15. Прединициаторлы кешеннің мРНК-ға қосылуы 4-ші топ факторлары көмегімен «Активтенген» мРНК өзіне рибосомальді прединициаторлы кешенді қоса алады.
- 16. мРНК инициаторлы AUG кодонын іздеу барысында сканирлеу Келесі кэп-тәуелді трансляция инициациясы ның маңызы мРНК құрамындағы AUG
- 17. 80S рибосомның AUG-кодонында жаналуы Инициаторлы тРНК антикодоны рибосоманың AUG 40S суббірлігіне дұрыс қосылғаннан кейін инициация факторларынан
- 18. eIF2-GDP-тің eIF2-GTP-ге айналуы Үш жақты кешеннің құрамында GTP гидролизі eIF2-GDP босалуына алып келеді. Көптеген G-белоктар сияқты,
- 19. мРНК-ның 3’-соңы трансляция инициациясына қатысуы 3’-НТО-ғы поли - А- тізбегі мРНКны 3’-5’ деградациясынан тек қорғап қана
- 21. Бактерияларда тек үш инициация факторы белгілі - IF1, IF2 және IF3, эукариоттарда - 9 канондық және
- 22. Белок синтезінің инициациясынан кейін әр жаңа амин қышқылы төрт шагтан тұратын (тРНҚ-ның байланысуы, пептидтік байланыстың түзілуі,
- 23. Трансляция циклының толық кескіні
- 24. Элонгация факторы трансляцияны алға жүргізеді және оның нақтылығын арттырады Элонгацияның екі факторы рибосомаға кіреді және әр
- 25. EF-Tu бір уақытта GTP мен аминоацил-тРНҚ молекуласын байланыстырады. Трансляцияны алға жылжытуға көмектескеннен бөлек, EF-Tu (эукариоттардағы EF1
- 26. Кодон-антикодон сәйкестігінің дұрыстығы РНҚ механизмі негізінде қалыптасқан рибосома арқылы бағаланады. Рибосоманың кіші суббірлігінің рРНҚ-сы кодон-антикодон жұбымен
- 27. Кіші суббірлік рибосомасының рРНҚ молекуласы арқылы дұрыс кодон-антикодон жұбын тану. Кіші суббірліктің рРНҚ молекуласы дұрыс байланысқан
- 28. Эукариоттардағы белок синтезінің инициациясы
- 29. Стоп-кодон трансляцияның аяқталуын белгілейді Стопкодондардың біреуінің болуы кодтаушы белок хабарламасының аяқталғандығын айтады (UAA, UAG немесе UGA).
- 30. Трансляцияның инициация машинасы бастамас бұрын 5´-кэп және полиА-ұшты таниды. Трансляцияның басталуына дейін экзондардың байланысу кешені (EJC)
- 31. мРНҚ-ның нонсенс-жанама ыдырауы
- 32. Белок құрамына кіретін амин қышқылы тізбегін құру үшін генетикалық код қолданылған кезде ген экспрессиясы процесі аяқталмайды.
- 33. Функционалды белсенді белоктың қалыптасу кезеңдері
- 34. Полипептидтік тізбектің жинақталған құрылымға айналуы кезінде көптеген гидрофобты қалдықтар оның ішінде қалып қояды. Сонымен қатар молекуланың
- 35. Еріген глобула құрылысы
- 36. Барлық белоктар өзінің синтезі кезінде жұмыршақталмайды. Рибосомадан шығар кезде оларды арнайы тағайындалған белоктар-молекулалық шаперондар күтіп алады.
- 37. Барлық Hsp белоктары толықтай емес оралған белоктардың беткейіндегі гидрофобтық учасоктарға ұқсас болып келеді және АТФ гидролазасының
- 38. Белоктың котрансляциялық шиыршықталуы
- 39. Шамадан тыс экспонирленген гидрофобты аймағы бар көптеген белоктар жасушада үлкен агрегаттарды түзе алады. Егер рефолдинг әрекеті
- 40. Белок фолдингіне заманауи түрде қарау
- 41. Протеасома көптеген белоктық суббірліктерден түзілген және төрт гептамерлік сақинадан тұратын квазицилиндрлік түтік түрінде жиналған орталық қуыс
- 42. Молекулалық шаперонның Hsp70 тобы
- 43. Протеасоманың қалпақшасында сақиналы құрылымы бар белоктар ААА белоктары ретінде танымал белоктық «анфолдаздың» үлкен класына жатады. 19S-қалпақшалар
- 44. Молекулалық шаперонның Hsp60 тобының құрылымы мен қызметі
- 45. Белоктың синтезінен кейінгі белок сапасын қадағалау процесі
- 46. Протеасома
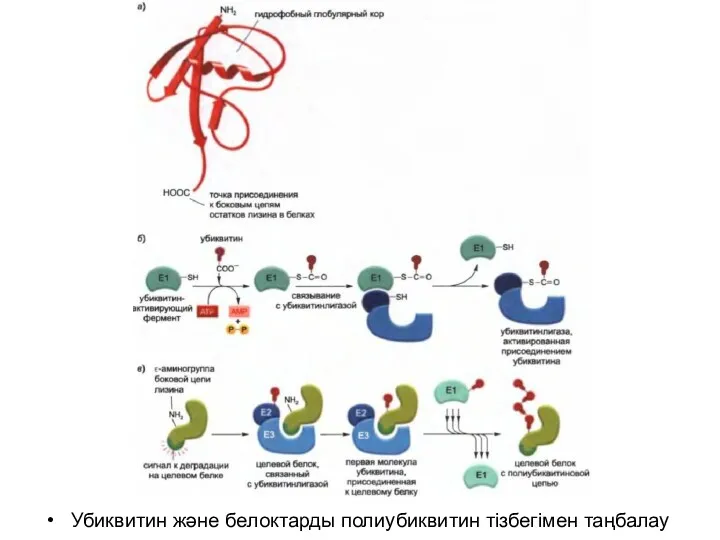
- 47. Убиквитиннің басқа белоктармен конъюгациясы үшін дайындығын АТФ тәуелді убиквитин-белсендіруші фермент (Е1) жүргізеді- ол белсендірілген Е1-байланысқан убиквитинді
- 48. Протеасома арқылы белоктың процессивті ыдырауы
- 49. Гексамерлі белоктық анфолдаза («развертаза»)
- 50. Убиквитин және белоктарды полиубиквитин тізбегімен таңбалау
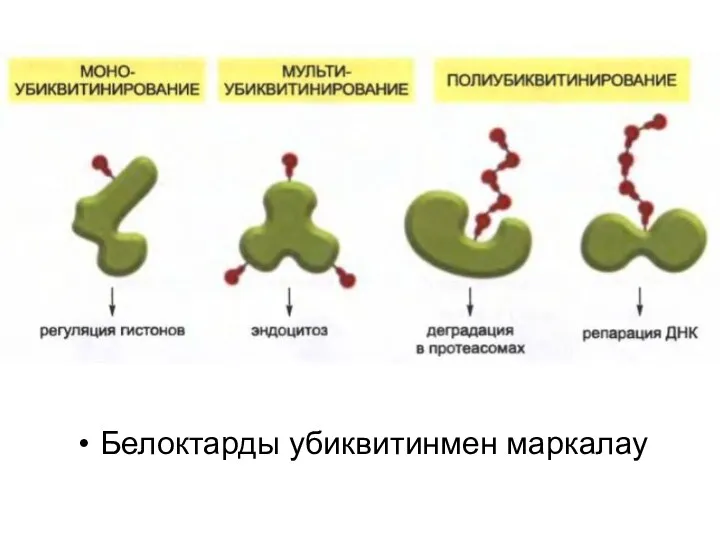
- 51. Белоктарды убиквитинмен маркалау
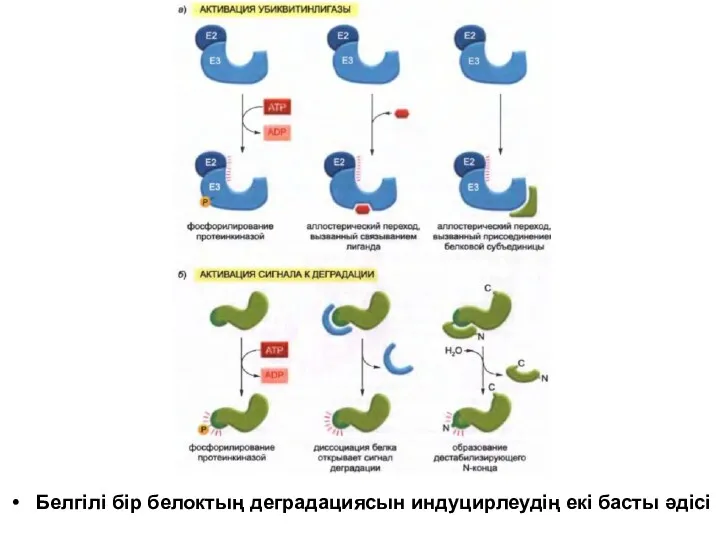
- 52. Белгілі бір белоктың деградациясын индуцирлеудің екі басты әдісі
- 53. Сау дарақтардағы жасушада кездесетін белок сапасын тексеретін жүйенің біртіндеп әлсіреуі салдарынан қалыпты белоктар арқылы агрегат түзіліп,
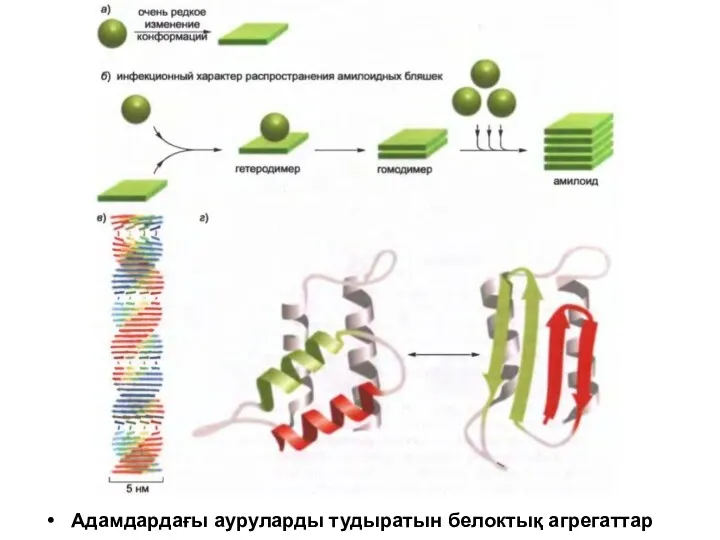
- 54. Адамдардағы ауруларды тудыратын белоктық агрегаттар
- 55. Адамдардағы аурудың туындауына әкеп соғатын белоктық шоғырланудың көбісі полипептидтік тізбектен тұратын, бір біріне β-жапырақшаларының үйіндісі түрінде
- 56. Приондық аурулар Хантингтон және Альцгеймер ауруларымен салыстырғанда приондық аурулар бір ағзадан екінші ағзаға берілуі мүмкін, егер
- 57. PrP тіптен спецификалық аномальды конформацияға өту тенденциясына ие. Бұл конформация протеазаға төзімді «кросс-бета» құрылымдарды ғана қалыптастырып
- 58. in vitro жағыдайында әртүрлі приондық штамдарды жасау
- 59. Эукариот жасушасындағы белоктың түзілуі
- 61. Скачать презентацию

мРНҚ тізбегі үш нуклеотид бойынша оқылады
Ақпараттың РНҚ формасынан белок формасына
мРНҚ тізбегі үш нуклеотид бойынша оқылады
Ақпараттың РНҚ формасынан белок формасына

мРНҚ молекуласындағы қатар біртіндеп үш нуклеотидтен тұратын топ арқылы оқылады. РНҚ
мРНҚ молекуласындағы қатар біртіндеп үш нуклеотидтен тұратын топ арқылы оқылады. РНҚ
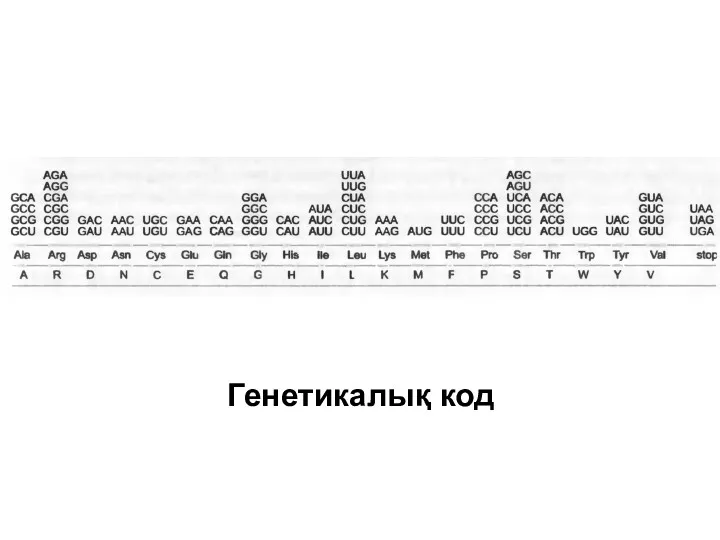
Генетикалық код
Генетикалық код

Бұл генетикалық код барлық заманауи ағзаларда бірегей қолданылады.
Алайда кодтан бірнеше
Бұл генетикалық код барлық заманауи ағзаларда бірегей қолданылады.
Алайда кодтан бірнеше
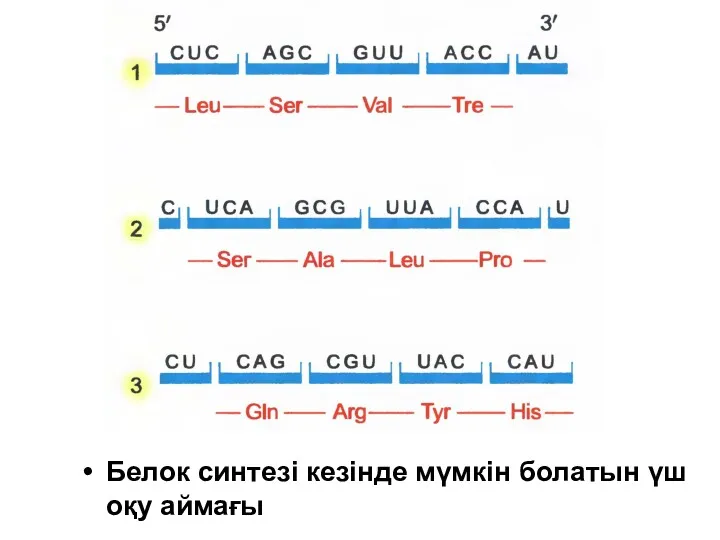
Белок синтезі кезінде мүмкін болатын үш оқу аймағы
Белок синтезі кезінде мүмкін болатын үш оқу аймағы

Жасушада белок синтезінің кезеңіне басты шек қоятын трансляция инициациясы – бұл
Жасушада белок синтезінің кезеңіне басты шек қоятын трансляция инициациясы – бұл
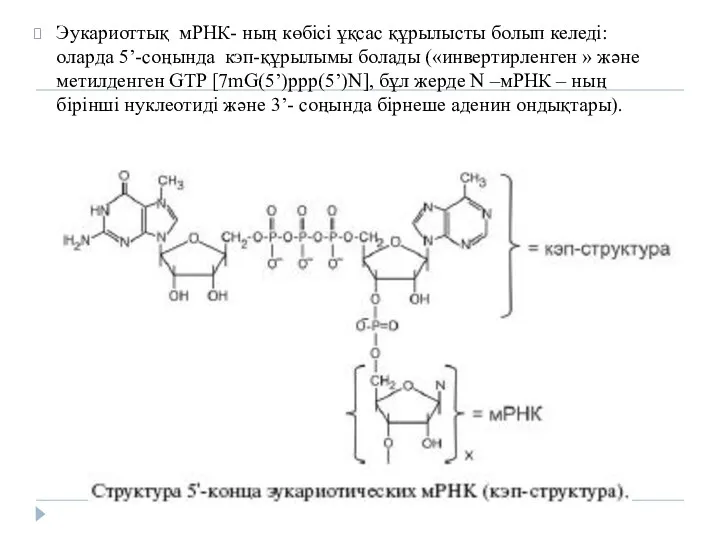
Эукариоттық мРНК- ның көбісі ұқсас құрылысты болып келеді: оларда 5’-соңында кэп-құрылымы
Эукариоттық мРНК- ның көбісі ұқсас құрылысты болып келеді: оларда 5’-соңында кэп-құрылымы

Трансляция инициациясы процесс ін сипаттау үшін өткен ғасырдың 80-жж аяғында «
Трансляция инициациясы процесс ін сипаттау үшін өткен ғасырдың 80-жж аяғында «

40S суббірлікке инициаторлы мРНК қосылуы. Рибосомальді прединициаторлы кешеннің түзілуі.
Трансляция инициациясында маңызды
40S суббірлікке инициаторлы мРНК қосылуы. Рибосомальді прединициаторлы кешеннің түзілуі.
Трансляция инициациясында маңызды
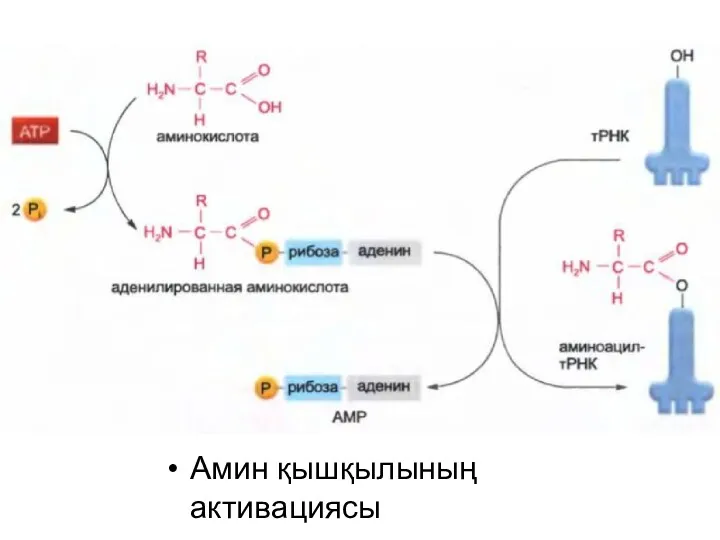
Амин қышқылының активациясы
Амин қышқылының активациясы
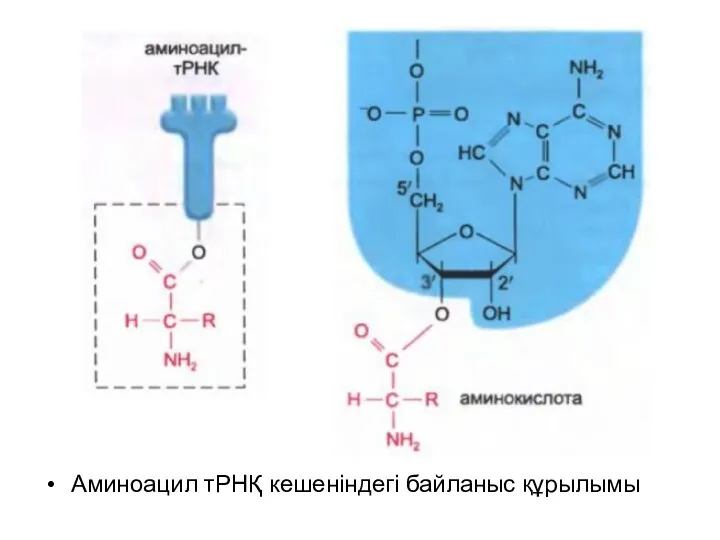
Аминоацил тРНҚ кешеніндегі байланыс құрылымы
Аминоацил тРНҚ кешеніндегі байланыс құрылымы


мРНК –ң трансляция инициациясына дайындалуы
Эукариот жасушасында бос мРНК, әдетте РНК
мРНК –ң трансляция инициациясына дайындалуы
Эукариот жасушасында бос мРНК, әдетте РНК

Прединициаторлы кешеннің мРНК-ға қосылуы
4-ші топ факторлары көмегімен «Активтенген» мРНК өзіне рибосомальді
Прединициаторлы кешеннің мРНК-ға қосылуы
4-ші топ факторлары көмегімен «Активтенген» мРНК өзіне рибосомальді

мРНК инициаторлы AUG кодонын іздеу барысында сканирлеу
Келесі кэп-тәуелді трансляция инициациясы ның
мРНК инициаторлы AUG кодонын іздеу барысында сканирлеу
Келесі кэп-тәуелді трансляция инициациясы ның

80S рибосомның AUG-кодонында жаналуы
Инициаторлы тРНК антикодоны рибосоманың AUG 40S суббірлігіне
80S рибосомның AUG-кодонында жаналуы
Инициаторлы тРНК антикодоны рибосоманың AUG 40S суббірлігіне

eIF2-GDP-тің eIF2-GTP-ге айналуы
Үш жақты кешеннің құрамында GTP гидролизі eIF2-GDP босалуына алып
eIF2-GDP-тің eIF2-GTP-ге айналуы
Үш жақты кешеннің құрамында GTP гидролизі eIF2-GDP босалуына алып

мРНК-ның 3’-соңы трансляция инициациясына қатысуы
3’-НТО-ғы поли - А- тізбегі мРНКны
мРНК-ның 3’-соңы трансляция инициациясына қатысуы
3’-НТО-ғы поли - А- тізбегі мРНКны
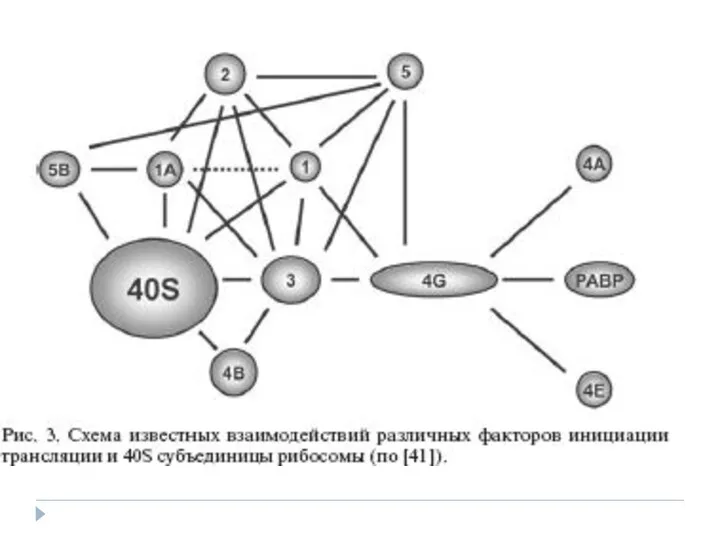

Бактерияларда тек үш инициация факторы белгілі - IF1, IF2 және IF3,
Бактерияларда тек үш инициация факторы белгілі - IF1, IF2 және IF3,

Белок синтезінің инициациясынан кейін әр жаңа амин қышқылы төрт шагтан тұратын
Белок синтезінің инициациясынан кейін әр жаңа амин қышқылы төрт шагтан тұратын
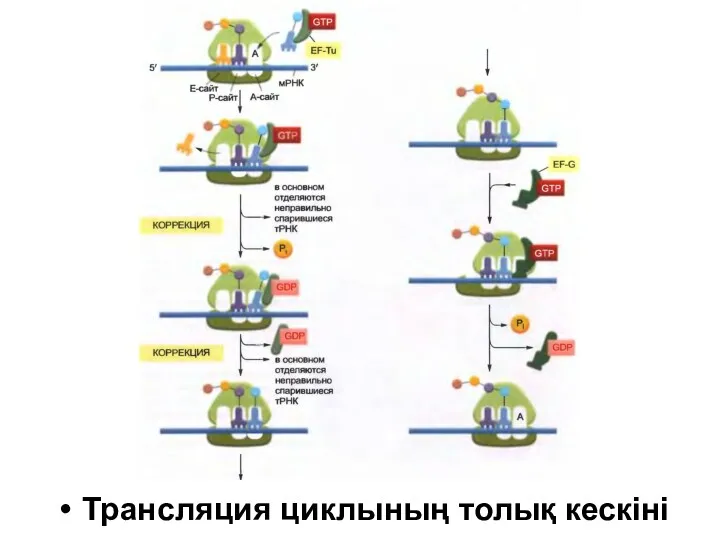
Трансляция циклының толық кескіні
Трансляция циклының толық кескіні

Элонгация факторы трансляцияны алға жүргізеді және оның нақтылығын арттырады
Элонгацияның екі факторы
Элонгация факторы трансляцияны алға жүргізеді және оның нақтылығын арттырады
Элонгацияның екі факторы

EF-Tu бір уақытта GTP мен аминоацил-тРНҚ молекуласын байланыстырады.
Трансляцияны алға жылжытуға
EF-Tu бір уақытта GTP мен аминоацил-тРНҚ молекуласын байланыстырады.
Трансляцияны алға жылжытуға

Кодон-антикодон сәйкестігінің дұрыстығы РНҚ механизмі негізінде қалыптасқан рибосома арқылы бағаланады.
Рибосоманың
Кодон-антикодон сәйкестігінің дұрыстығы РНҚ механизмі негізінде қалыптасқан рибосома арқылы бағаланады.
Рибосоманың
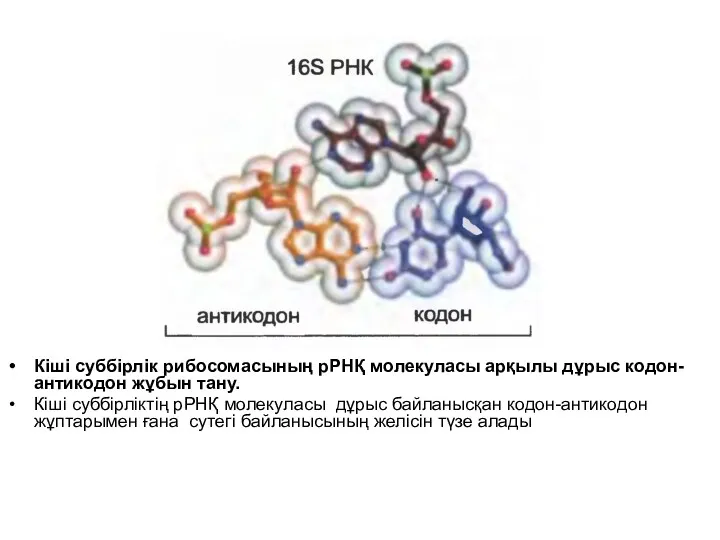
Кіші суббірлік рибосомасының рРНҚ молекуласы арқылы дұрыс кодон-антикодон жұбын тану.
Кіші
Кіші суббірлік рибосомасының рРНҚ молекуласы арқылы дұрыс кодон-антикодон жұбын тану.
Кіші
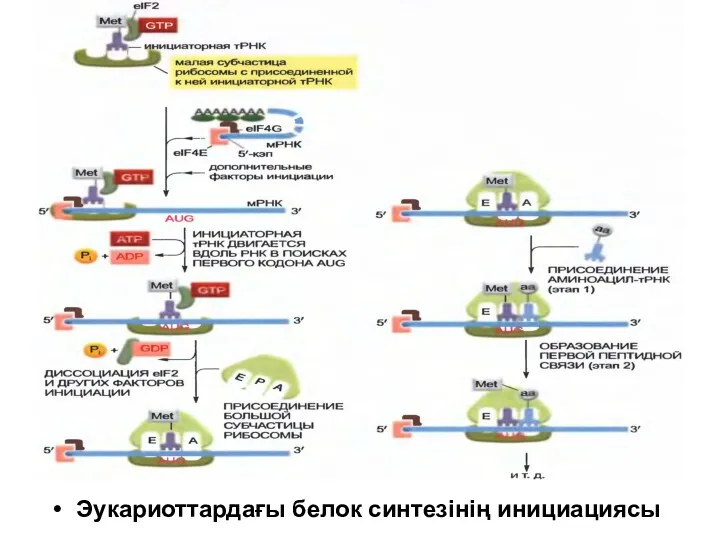
Эукариоттардағы белок синтезінің инициациясы
Эукариоттардағы белок синтезінің инициациясы

Стоп-кодон трансляцияның аяқталуын белгілейді
Стопкодондардың біреуінің болуы кодтаушы белок хабарламасының аяқталғандығын айтады
Стоп-кодон трансляцияның аяқталуын белгілейді
Стопкодондардың біреуінің болуы кодтаушы белок хабарламасының аяқталғандығын айтады

Трансляцияның инициация машинасы бастамас бұрын 5´-кэп және полиА-ұшты таниды.
Трансляцияның басталуына
Трансляцияның инициация машинасы бастамас бұрын 5´-кэп және полиА-ұшты таниды.
Трансляцияның басталуына
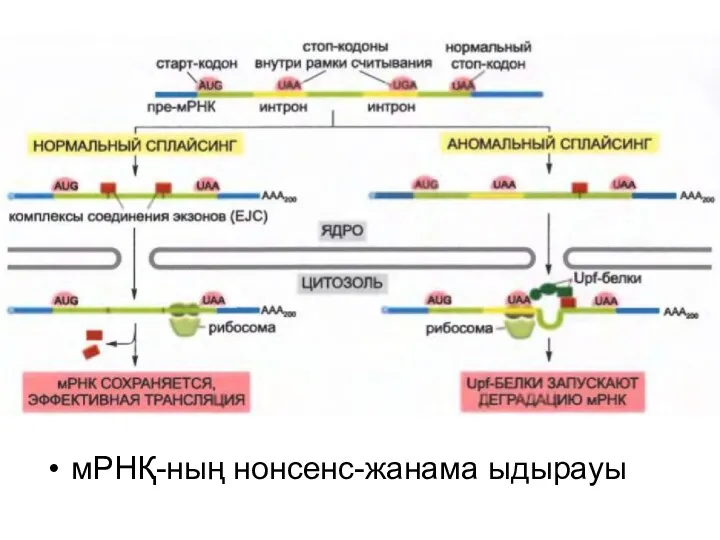
мРНҚ-ның нонсенс-жанама ыдырауы
мРНҚ-ның нонсенс-жанама ыдырауы

Белок құрамына кіретін амин қышқылы тізбегін құру үшін генетикалық код қолданылған
Белок құрамына кіретін амин қышқылы тізбегін құру үшін генетикалық код қолданылған
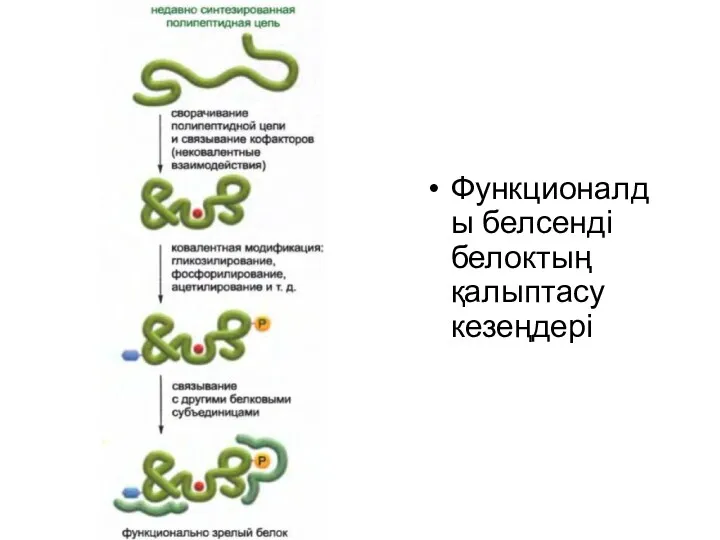
Функционалды белсенді белоктың қалыптасу кезеңдері
Функционалды белсенді белоктың қалыптасу кезеңдері

Полипептидтік тізбектің жинақталған құрылымға айналуы кезінде көптеген гидрофобты қалдықтар оның ішінде
Полипептидтік тізбектің жинақталған құрылымға айналуы кезінде көптеген гидрофобты қалдықтар оның ішінде
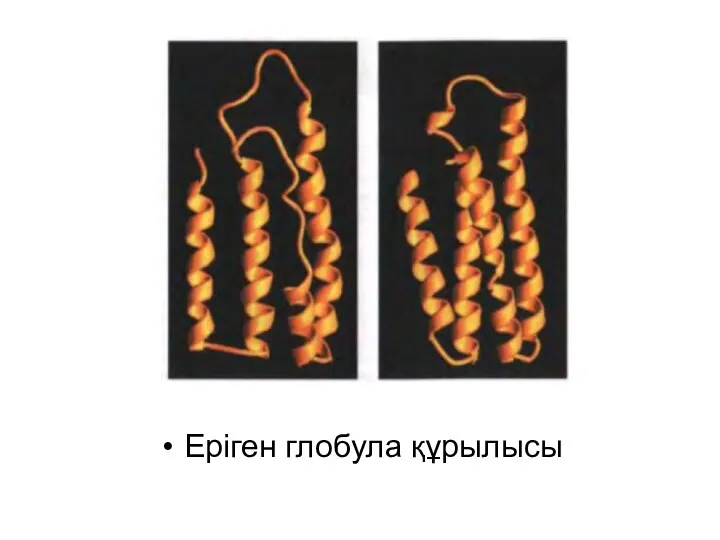
Еріген глобула құрылысы
Еріген глобула құрылысы

Барлық белоктар өзінің синтезі кезінде жұмыршақталмайды. Рибосомадан шығар кезде оларды арнайы
Барлық белоктар өзінің синтезі кезінде жұмыршақталмайды. Рибосомадан шығар кезде оларды арнайы

Барлық Hsp белоктары толықтай емес оралған белоктардың беткейіндегі гидрофобтық учасоктарға ұқсас
Барлық Hsp белоктары толықтай емес оралған белоктардың беткейіндегі гидрофобтық учасоктарға ұқсас
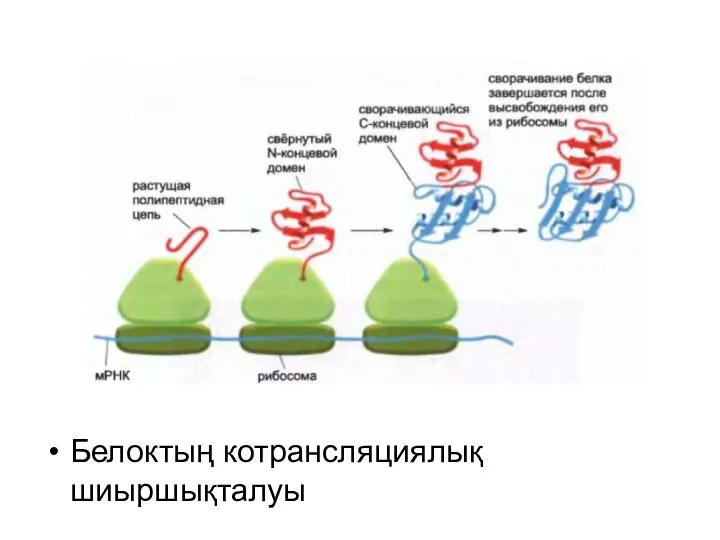
Белоктың котрансляциялық шиыршықталуы
Белоктың котрансляциялық шиыршықталуы

Шамадан тыс экспонирленген гидрофобты аймағы бар көптеген белоктар жасушада үлкен агрегаттарды
Шамадан тыс экспонирленген гидрофобты аймағы бар көптеген белоктар жасушада үлкен агрегаттарды
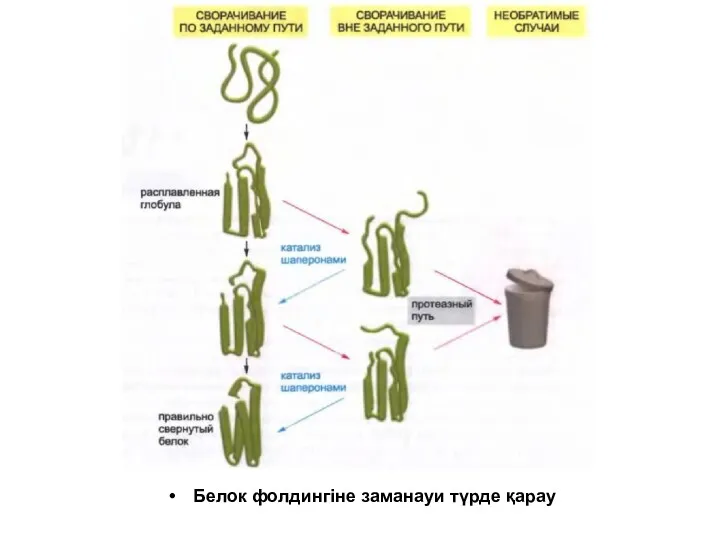
Белок фолдингіне заманауи түрде қарау
Белок фолдингіне заманауи түрде қарау

Протеасома көптеген белоктық суббірліктерден түзілген және төрт гептамерлік сақинадан тұратын квазицилиндрлік
Протеасома көптеген белоктық суббірліктерден түзілген және төрт гептамерлік сақинадан тұратын квазицилиндрлік
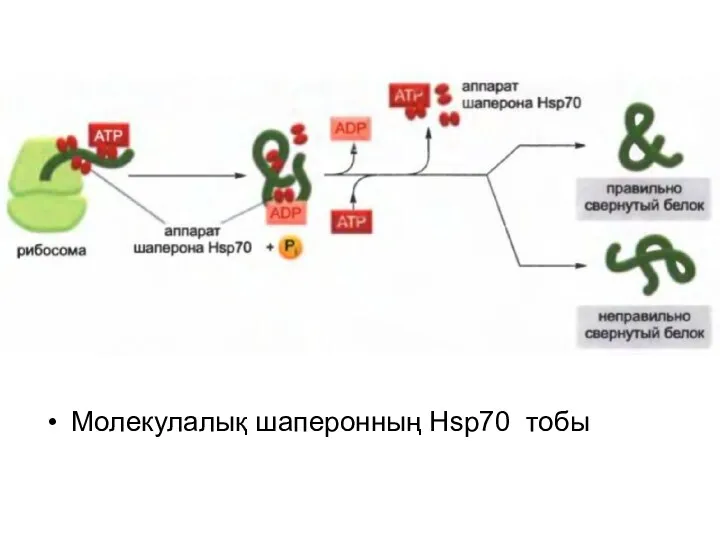
Молекулалық шаперонның Hsp70 тобы
Молекулалық шаперонның Hsp70 тобы

Протеасоманың қалпақшасында сақиналы құрылымы бар белоктар ААА белоктары ретінде танымал белоктық
Протеасоманың қалпақшасында сақиналы құрылымы бар белоктар ААА белоктары ретінде танымал белоктық
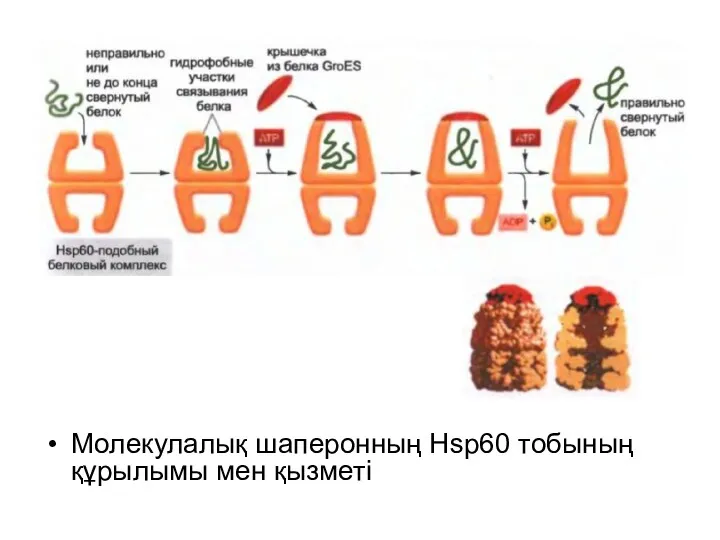
Молекулалық шаперонның Hsp60 тобының құрылымы мен қызметі
Молекулалық шаперонның Hsp60 тобының құрылымы мен қызметі
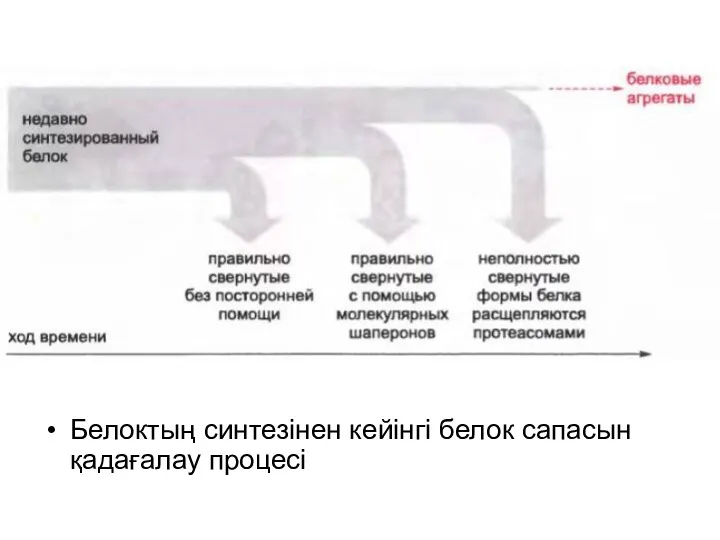
Белоктың синтезінен кейінгі белок сапасын қадағалау процесі
Белоктың синтезінен кейінгі белок сапасын қадағалау процесі
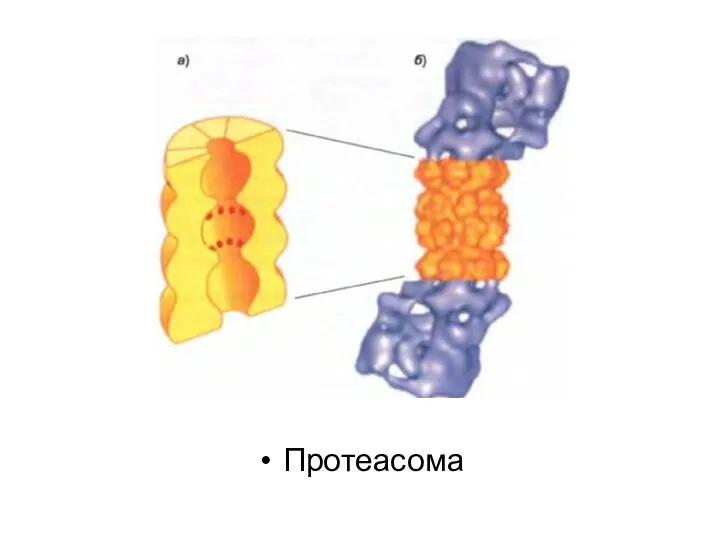
Протеасома
Протеасома

Убиквитиннің басқа белоктармен конъюгациясы үшін дайындығын АТФ тәуелді убиквитин-белсендіруші фермент (Е1)
Убиквитиннің басқа белоктармен конъюгациясы үшін дайындығын АТФ тәуелді убиквитин-белсендіруші фермент (Е1)
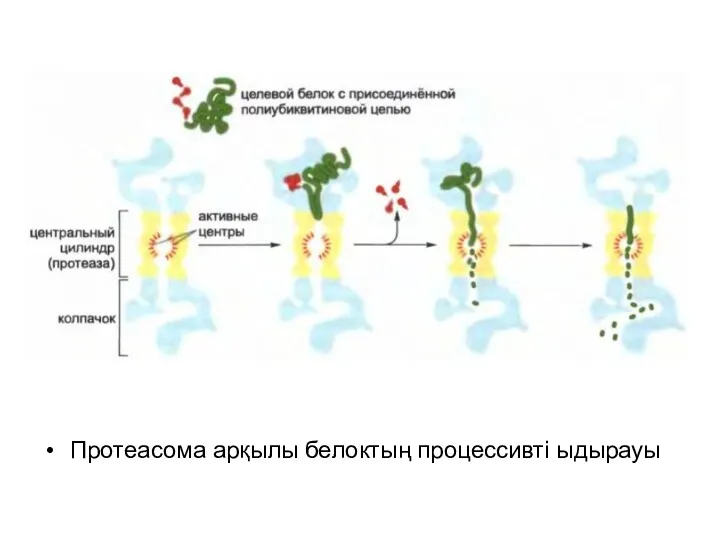
Протеасома арқылы белоктың процессивті ыдырауы
Протеасома арқылы белоктың процессивті ыдырауы
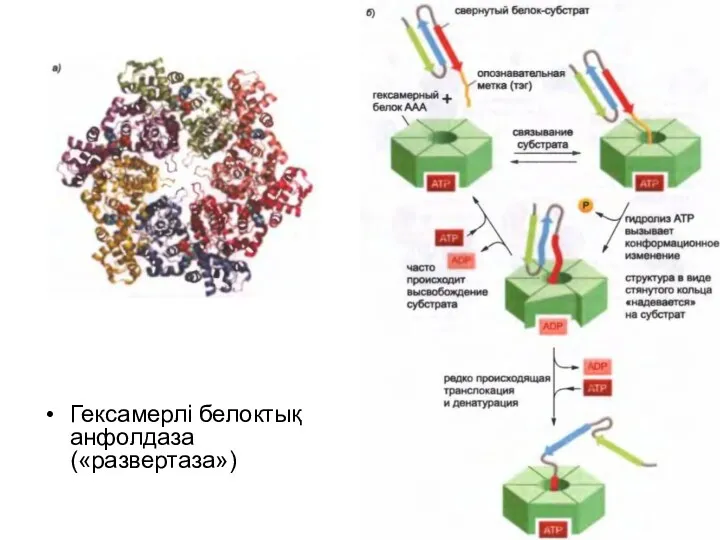
Гексамерлі белоктық анфолдаза («развертаза»)
Гексамерлі белоктық анфолдаза («развертаза»)
Убиквитин және белоктарды полиубиквитин тізбегімен таңбалау
Убиквитин және белоктарды полиубиквитин тізбегімен таңбалау
Белоктарды убиквитинмен маркалау
Белоктарды убиквитинмен маркалау
Белгілі бір белоктың деградациясын индуцирлеудің екі басты әдісі
Белгілі бір белоктың деградациясын индуцирлеудің екі басты әдісі

Сау дарақтардағы жасушада кездесетін белок сапасын тексеретін жүйенің біртіндеп әлсіреуі салдарынан
Сау дарақтардағы жасушада кездесетін белок сапасын тексеретін жүйенің біртіндеп әлсіреуі салдарынан
Адамдардағы ауруларды тудыратын белоктық агрегаттар
Адамдардағы ауруларды тудыратын белоктық агрегаттар

Адамдардағы аурудың туындауына әкеп соғатын белоктық шоғырланудың көбісі полипептидтік тізбектен тұратын,
Адамдардағы аурудың туындауына әкеп соғатын белоктық шоғырланудың көбісі полипептидтік тізбектен тұратын,

Приондық аурулар
Хантингтон және Альцгеймер ауруларымен салыстырғанда приондық аурулар бір ағзадан екінші
Приондық аурулар
Хантингтон және Альцгеймер ауруларымен салыстырғанда приондық аурулар бір ағзадан екінші

PrP тіптен спецификалық аномальды конформацияға өту тенденциясына ие.
Бұл конформация протеазаға
PrP тіптен спецификалық аномальды конформацияға өту тенденциясына ие.
Бұл конформация протеазаға
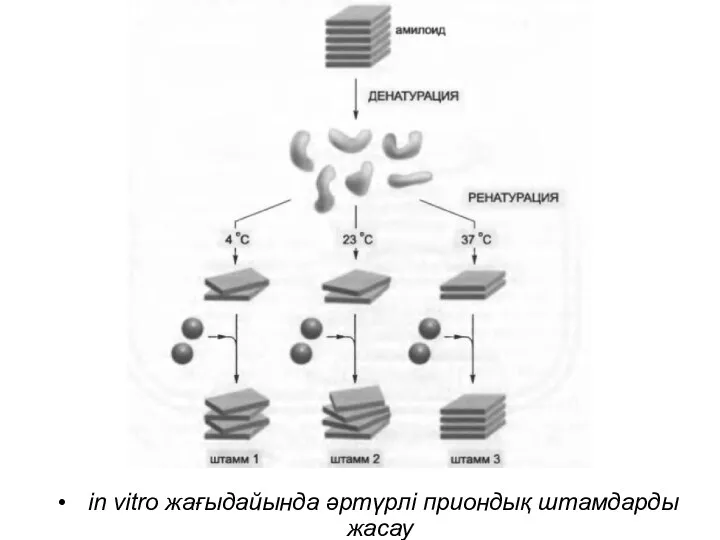
in vitro жағыдайында әртүрлі приондық штамдарды жасау
in vitro жағыдайында әртүрлі приондық штамдарды жасау
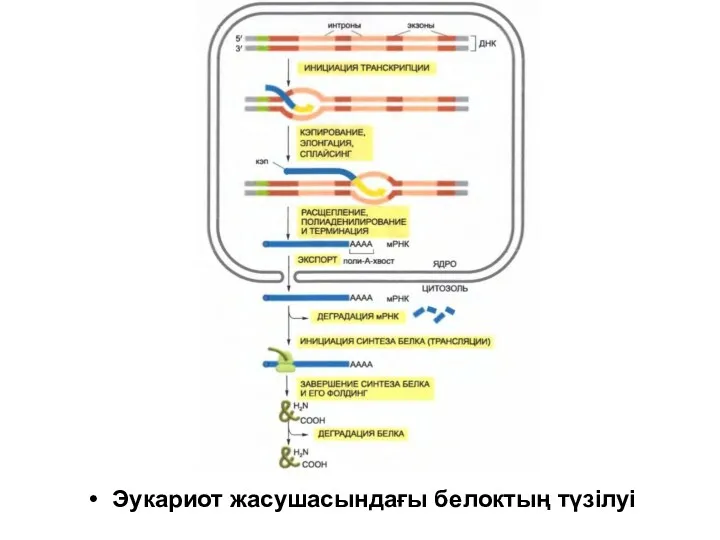
Эукариот жасушасындағы белоктың түзілуі
Эукариот жасушасындағы белоктың түзілуі
 Nucleic acids
Nucleic acids Развитие органического мира на Земле
Развитие органического мира на Земле Эволюция опорно- двигательной системы у животных
Эволюция опорно- двигательной системы у животных Апаптоз кезіндегі молекулалық оқиғалардың жалпы сипаттамасы
Апаптоз кезіндегі молекулалық оқиғалардың жалпы сипаттамасы Минирующие насекомые. (Лекция 8)
Минирующие насекомые. (Лекция 8) Фотосинтез
Фотосинтез Уголок живой природы как средство формирования исследовательских способностей обучающихся по биологии
Уголок живой природы как средство формирования исследовательских способностей обучающихся по биологии Индивидуальное развитие организмов
Индивидуальное развитие организмов Информация и ее роль в обществе
Информация и ее роль в обществе Структура и содержание курса общей биологии. 10 класс
Структура и содержание курса общей биологии. 10 класс Бактериалық жасушаның химиялық құрамы
Бактериалық жасушаның химиялық құрамы Class ear
Class ear Среда обитания
Среда обитания Цветы. Уход за цветами
Цветы. Уход за цветами Цветочная карусель
Цветочная карусель Стадии онтогенеза почв
Стадии онтогенеза почв Дыхательная система
Дыхательная система Wild and domestic animals
Wild and domestic animals Лимфатическая система
Лимфатическая система Учебный проект Экспериментальное определение качества молочной продукции
Учебный проект Экспериментальное определение качества молочной продукции Животные Севера
Животные Севера Cад (окружающий мир, 1 класс)
Cад (окружающий мир, 1 класс) Эндокринная система человека
Эндокринная система человека Первоцветы. Биологические особенности раннецветущих растений
Первоцветы. Биологические особенности раннецветущих растений Дикая природа России
Дикая природа России Водоросли. Общая характеристика
Водоросли. Общая характеристика Сторінками Червоної книги України
Сторінками Червоної книги України Основные загрязнители почвы
Основные загрязнители почвы