Транспорт асимілятів як основна об΄єднуюча ланка для пересування метаболітів та їх використання в синтетичних процесах презентация
- Транспорт асимілятів як основна об΄єднуюча ланка для пересування метаболітів та їх використання в синтетичних процесах

Содержание
- 2. Продуктами фотосинтезу вважають низку речовин, що утворюються в хлоропластах в результаті засвоєння вуглекислоти. Первинним продуктом фотосинтезу
- 4. Синтез вуглеводів при фотосинтезі
- 5. Відбувається конкуренція між двома ферментними системами: АДФ-глюкопірофосфорилази і спряженої з нею крохмальсинтази та УДФ-глюкопірофосфорилази і спряженої
- 6. Значна частина продуктів фотосинтезу використовується безпосередньо в клітинах мезофілу, решта транспортується в інші органи рослини. Відтік
- 7. Дальнє транспортування продуктів фотосинтезу здійснюється по провідних пучках та по флоемі. Основна транспортна форма асимілятів –
- 8. Рух асимілятів – активний процес, що відбувається за рахунок енергії дихання. Швидкість пересування асимілятів по флоемі
- 10. Транспорт речовин по рослині Проблема транспорту речовин має три аспекти: По яких тканинах відбувається Які є
- 11. Пересування речовин по рослині
- 12. Ближній транспорт – переміщення іонів, метаболітів і води між клітинами й тканинами (на відміну від мембранного
- 13. Рух води з розчиненими речовинами може відбуватись: по провідних пучках; по клітинних стінках (апопласту); по цитоплазмі
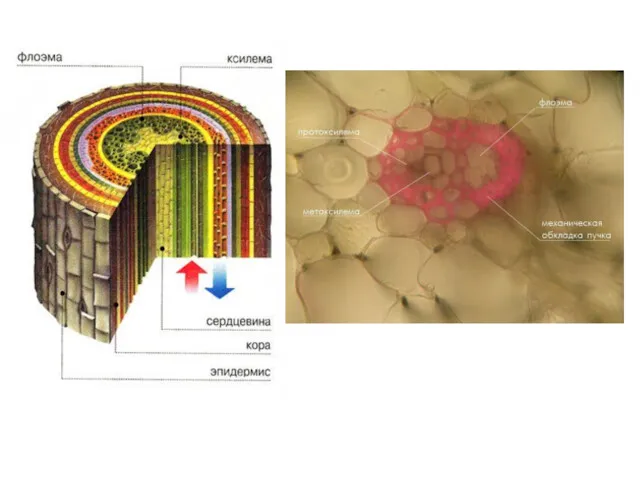
- 14. У всіх вищих наземних рослин є провідні пучки, які складаються з трахеїд і судин ксилеми; ситовидних
- 15. Рух розчинених речовин по провідних пучках здійснюється: по ксилемі (висхідний потік); по флоемі (низхідний потік від
- 16. ксилема флоема
- 18. Транспорт продуктів фотосинтезу у генеративні органи Надходження вуглекислого газу, транспірація Утворення продуктів фотосинтезу Надходження елементів живлення
- 20. паренхіма ендодерма
- 21. Ксилемний транспорт Судини і трахеїди в деревині клена (х350)
- 22. Ксилемний сік – розчин неорганічних речовин. Однак у пасоці із ксилеми пенька виявили різні азотисті сполуки
- 23. Механізми ксилемного транспорту Завантажування ксилеми найінтенсивніше відбувається в зоні кореневих волосків. Тут працює декілька насосів, на
- 24. В паренхімних клітинах пучків діє інша помпа, котра перекачує мінеральні речовини через пори трахеїд до їх
- 25. Вода і розчинені в ній речовини рухаються по судинах ксилеми з великою швидкістю ≈ 1-20 м/год.
- 26. Вміст органічних та неорганічних речовин у ксилемі залежить від виду рослини та умов мінерального живлення. По
- 27. Регуляція ксилемного транспорту здійснюється через функціонування Н+-помп і інших іонних насосів, що пов'язано з енергетикою дихання
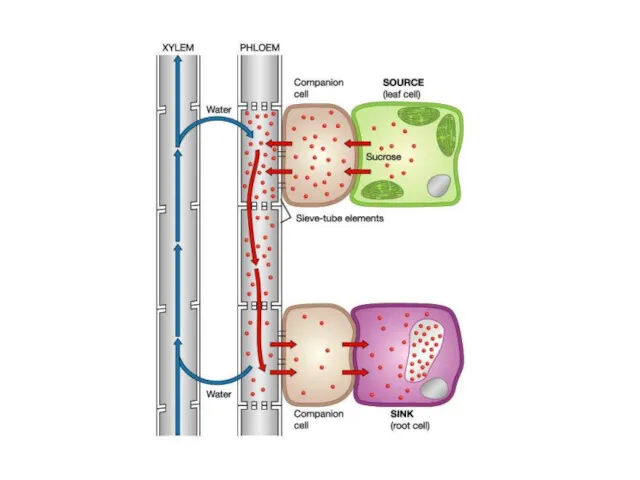
- 28. Флоемний транспорт Асиміляти із листків надходять у флоему, яка складається із спеціалізованих клітин. Транспортну функцію здійснюють
- 29. Клітини ситовидних трубок містять невелику кількість органоїдів, тонопласт зруйнований, у зрілих клітин ядро відсутнє. Поперечні стінки-пластинки
- 31. Провідна система складає 25 % від загального об'єму листка. Загальна довжина провідних пучків в 1 см2
- 33. Склад флоемного соку. Основна транспортна форма цукрів – сахароза. Її вміст у флоемному сокові може досягати
- 34. Наявні фітогормони, органічні кислоти, вітаміни. рН ~ 8,0-8,5. АТФ міститься в концентрації 0,5 ммоль/л і надходить,
- 35. Механізми флоемного транспорту Рух асимілятів по ситовидних трубках відбувається зі швидкістю 50-100 см/год і включає в
- 36. Флоемний ексудат містить різноманітні іони. З ксилемного соку у флоемний легко циркулюють К, Na, Mg, P,
- 37. Транспорт речовин по ситовидних трубках. Рух асимілятів по ситовидних елементах – особливий випадок симпластичного транспорту. Механізм
- 38. ІІ-га гіпотеза. Рушійною силою переміщення флоемної рідини з однієї ситовидної трубки в іншу через пори може
- 39. Для розвантажування флоеми має значення підвищений гідростатичний тиск у ситовидних трубках. Крім того, органи, які інтенсивно
- 40. Регуляція флоемного транспорту Флоемний транспорт включає в себе ряд транспортних систем із своїми механізмами регуляції. На
- 41. При завантажуванні флоемних закінчень регуляція здійснюється на рівні Н+-помпи, яка активується фітогормонами (ауксин). АБК блокує Н+/К+
- 43. Скачать презентацию

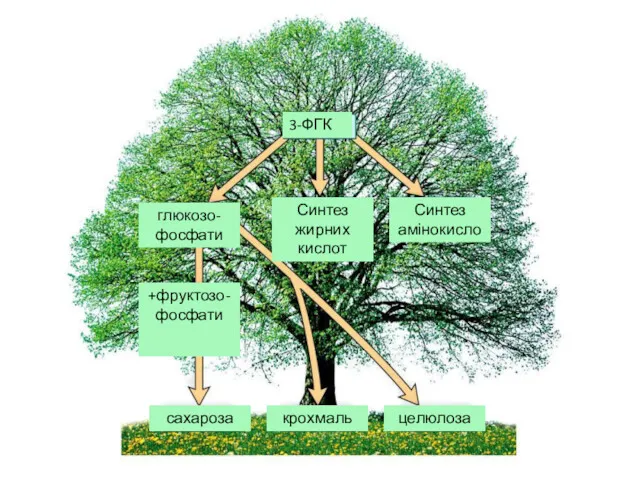
Продуктами фотосинтезу вважають низку речовин, що утворюються в хлоропластах в результаті
Продуктами фотосинтезу вважають низку речовин, що утворюються в хлоропластах в результаті
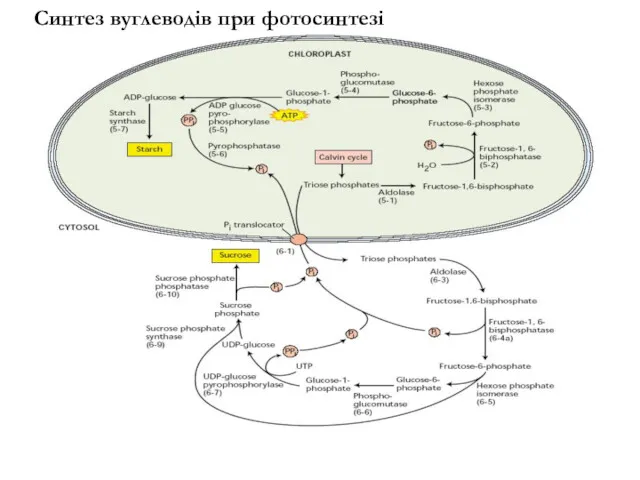
Синтез вуглеводів при фотосинтезі
Синтез вуглеводів при фотосинтезі
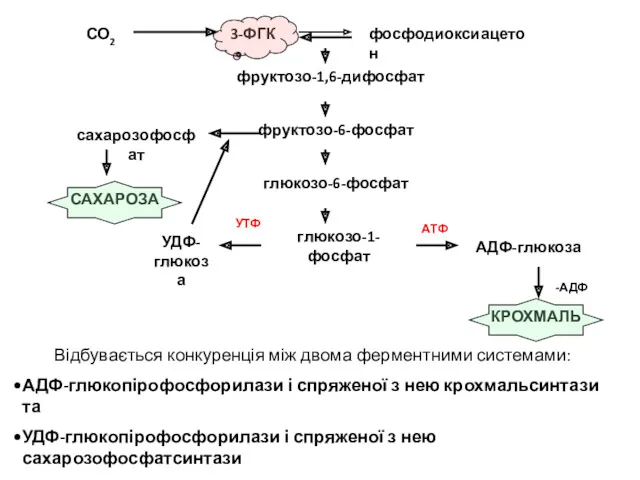
Відбувається конкуренція між двома ферментними системами:
АДФ-глюкопірофосфорилази і спряженої з нею
Відбувається конкуренція між двома ферментними системами:
АДФ-глюкопірофосфорилази і спряженої з нею

Значна частина продуктів фотосинтезу використовується безпосередньо в клітинах мезофілу, решта транспортується
Значна частина продуктів фотосинтезу використовується безпосередньо в клітинах мезофілу, решта транспортується

Дальнє транспортування продуктів фотосинтезу здійснюється по провідних пучках та по флоемі.
Дальнє транспортування продуктів фотосинтезу здійснюється по провідних пучках та по флоемі.

Рух асимілятів – активний процес, що відбувається за рахунок енергії дихання.
Рух асимілятів – активний процес, що відбувається за рахунок енергії дихання.
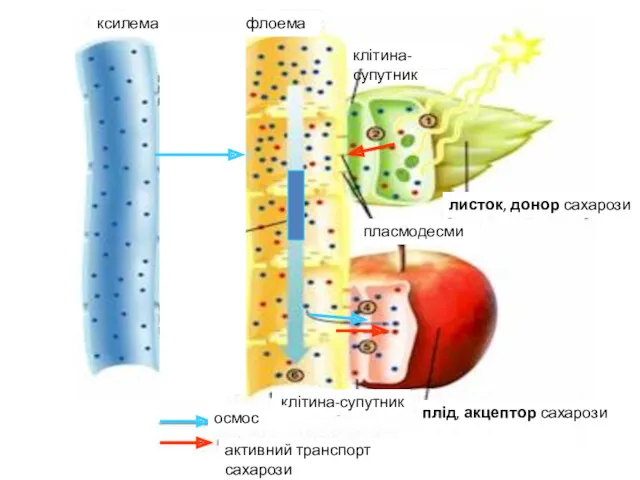

Транспорт речовин по рослині
Проблема транспорту речовин має три аспекти:
По яких тканинах
Транспорт речовин по рослині
Проблема транспорту речовин має три аспекти:
По яких тканинах
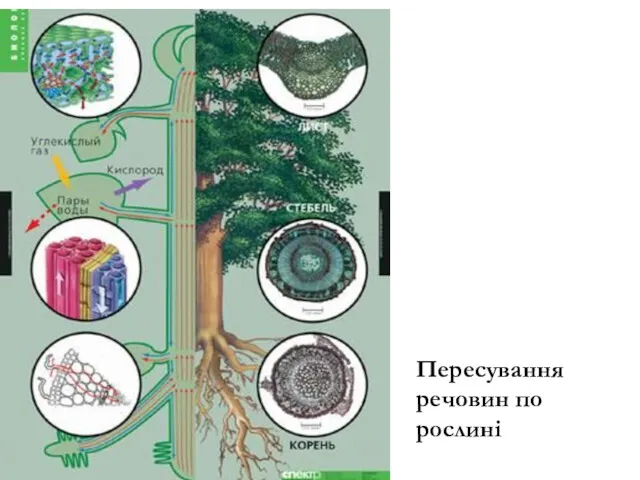
Пересування речовин по рослині
Пересування речовин по рослині

Ближній транспорт – переміщення іонів, метаболітів і води між клітинами
Ближній транспорт – переміщення іонів, метаболітів і води між клітинами

Рух води з розчиненими речовинами може відбуватись:
по провідних пучках;
по
Рух води з розчиненими речовинами може відбуватись:
по провідних пучках;
по

У всіх вищих наземних рослин є провідні пучки, які складаються з
У всіх вищих наземних рослин є провідні пучки, які складаються з

Рух розчинених речовин по провідних пучках здійснюється:
по ксилемі (висхідний потік);
Рух розчинених речовин по провідних пучках здійснюється:
по ксилемі (висхідний потік);
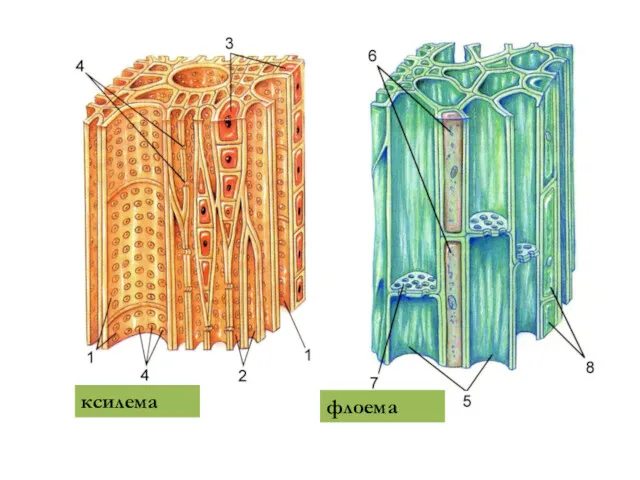
ксилема
флоема
ксилема
флоема
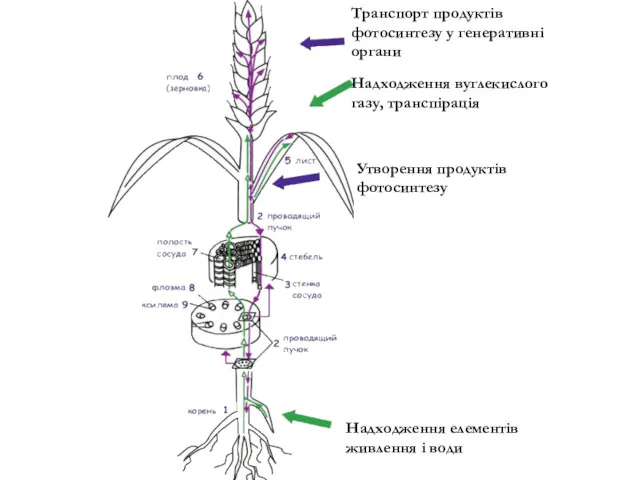
Транспорт продуктів фотосинтезу у генеративні органи
Надходження вуглекислого газу, транспірація
Утворення продуктів фотосинтезу
Надходження
Транспорт продуктів фотосинтезу у генеративні органи
Надходження вуглекислого газу, транспірація
Утворення продуктів фотосинтезу
Надходження
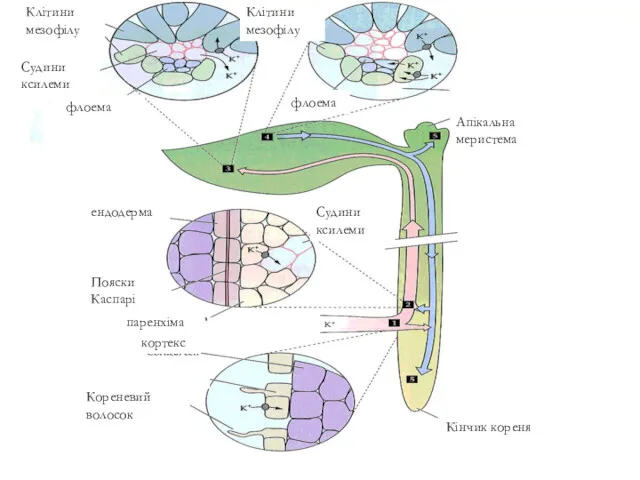
паренхіма
ендодерма
паренхіма
ендодерма
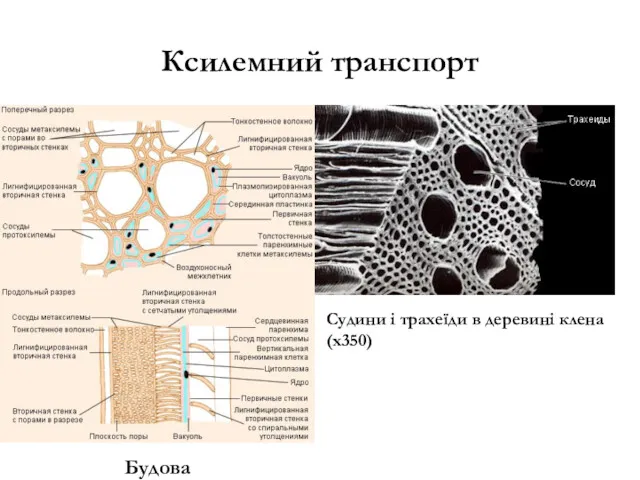
Ксилемний транспорт
Судини і трахеїди в деревині клена (х350)
Ксилемний транспорт
Судини і трахеїди в деревині клена (х350)

Ксилемний сік – розчин неорганічних речовин. Однак у пасоці із ксилеми
Ксилемний сік – розчин неорганічних речовин. Однак у пасоці із ксилеми

Механізми ксилемного транспорту
Завантажування ксилеми найінтенсивніше відбувається в зоні кореневих волосків.
Тут
Механізми ксилемного транспорту
Завантажування ксилеми найінтенсивніше відбувається в зоні кореневих волосків.
Тут

В паренхімних клітинах пучків діє інша помпа, котра перекачує мінеральні речовини
В паренхімних клітинах пучків діє інша помпа, котра перекачує мінеральні речовини

Вода і розчинені в ній речовини рухаються по судинах ксилеми з
Вода і розчинені в ній речовини рухаються по судинах ксилеми з

Вміст органічних та неорганічних речовин у ксилемі залежить від виду рослини
Вміст органічних та неорганічних речовин у ксилемі залежить від виду рослини

Регуляція ксилемного транспорту здійснюється через функціонування Н+-помп і інших іонних насосів,
Регуляція ксилемного транспорту здійснюється через функціонування Н+-помп і інших іонних насосів,

Флоемний транспорт
Асиміляти із листків надходять у флоему, яка складається із
Флоемний транспорт
Асиміляти із листків надходять у флоему, яка складається із
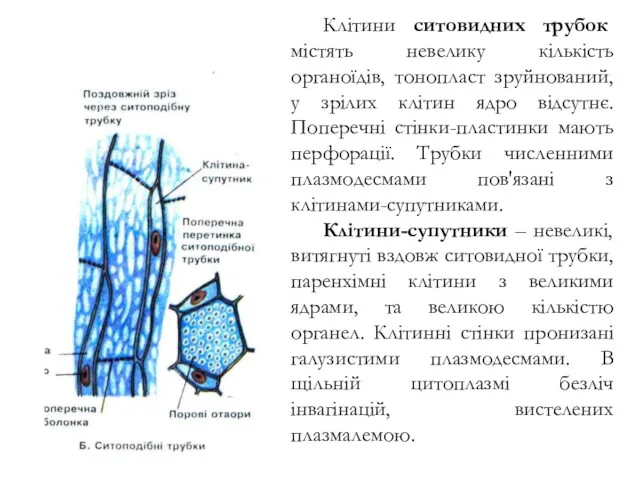
Клітини ситовидних трубок містять невелику кількість органоїдів, тонопласт зруйнований, у зрілих
Клітини ситовидних трубок містять невелику кількість органоїдів, тонопласт зруйнований, у зрілих
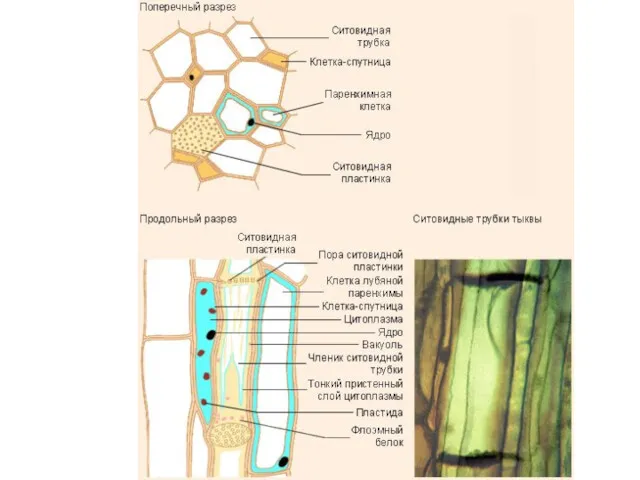

Провідна система складає 25 % від загального об'єму листка. Загальна довжина
Провідна система складає 25 % від загального об'єму листка. Загальна довжина


Склад флоемного соку.
Основна транспортна форма цукрів – сахароза. Її вміст
Склад флоемного соку.
Основна транспортна форма цукрів – сахароза. Її вміст

Наявні фітогормони, органічні кислоти, вітаміни. рН ~ 8,0-8,5. АТФ міститься в
Наявні фітогормони, органічні кислоти, вітаміни. рН ~ 8,0-8,5. АТФ міститься в

Механізми флоемного транспорту
Рух асимілятів по ситовидних трубках відбувається зі швидкістю
Механізми флоемного транспорту
Рух асимілятів по ситовидних трубках відбувається зі швидкістю

Флоемний ексудат містить різноманітні іони. З ксилемного соку у флоемний легко
Флоемний ексудат містить різноманітні іони. З ксилемного соку у флоемний легко

Транспорт речовин по ситовидних трубках.
Рух асимілятів по ситовидних елементах –
Транспорт речовин по ситовидних трубках.
Рух асимілятів по ситовидних елементах –

ІІ-га гіпотеза. Рушійною силою переміщення флоемної рідини з однієї ситовидної трубки
ІІ-га гіпотеза. Рушійною силою переміщення флоемної рідини з однієї ситовидної трубки

Для розвантажування флоеми має значення підвищений гідростатичний тиск у ситовидних трубках.
Для розвантажування флоеми має значення підвищений гідростатичний тиск у ситовидних трубках.

Регуляція флоемного транспорту
Флоемний транспорт включає в себе ряд транспортних систем із
Регуляція флоемного транспорту
Флоемний транспорт включає в себе ряд транспортних систем із

При завантажуванні флоемних закінчень регуляція здійснюється на рівні Н+-помпи, яка активується
При завантажуванні флоемних закінчень регуляція здійснюється на рівні Н+-помпи, яка активується
 Класс Паукообразные, особенности строения и жизнедеятельности
Класс Паукообразные, особенности строения и жизнедеятельности Нейро-гуморальная регуляция процессов жизнедеятельности организма
Нейро-гуморальная регуляция процессов жизнедеятельности организма Строение и свойства биогеоценоза
Строение и свойства биогеоценоза Biochimia. Enzimele
Biochimia. Enzimele Животные Южной Америки
Животные Южной Америки Биотехнология в селекции растений. Часть 1. Селекция на важнейшие хозяйственные свойства
Биотехнология в селекции растений. Часть 1. Селекция на важнейшие хозяйственные свойства Морфология и строение вирусов
Морфология и строение вирусов Клетка: строение, химический состав, жизнедеятельность
Клетка: строение, химический состав, жизнедеятельность Основы рационального питания
Основы рационального питания Селекція тварин
Селекція тварин Царство растения
Царство растения Витамин В1
Витамин В1 Перелетные птицы
Перелетные птицы Строение волос
Строение волос У нас на даче. Кроссворд
У нас на даче. Кроссворд сезонные явления в жизни птиц
сезонные явления в жизни птиц Изучение морфологического критерия вида на примере растений рода Клевер
Изучение морфологического критерия вида на примере растений рода Клевер Закономерности изменчивости
Закономерности изменчивости Уникальные грибы
Уникальные грибы Генетика. Третий закон Менделя
Генетика. Третий закон Менделя Биология как наука. Методы познания живой природы
Биология как наука. Методы познания живой природы презентация Виртуальное путешествие по реке Белая НАО
презентация Виртуальное путешествие по реке Белая НАО Зоология позвоночных. Амниоты, первично-наземные позвоночные. (Лекция 8)
Зоология позвоночных. Амниоты, первично-наземные позвоночные. (Лекция 8) Отряд бабочки
Отряд бабочки Морфофункциональная организация скелетных мышц
Морфофункциональная организация скелетных мышц Разнообразие грибов. Значение грибов в природе и жизни человека
Разнообразие грибов. Значение грибов в природе и жизни человека Царство Растения
Царство Растения Уходы за посевами и сеянцами в посевном отделении питомника. Лекция-9
Уходы за посевами и сеянцами в посевном отделении питомника. Лекция-9