- Транспорт газов кровью. Газообмен в тканях. Регуляция дыхания

Содержание
- 2. ДЫХАНИЕ. Транспорт газов кровью Газообмен в тканях Регуляция дыхания
- 4. ЗАКОН ФИКА: М = k⋅S/L⋅ΔP: где, коэффициент диффузии (k) зависит от природы газа, температуры и среды,
- 5. КЕК и гемоглобин В крови легких при взаимодействие О2 с Fе2+ гема , образуется легко диссоциирующее
- 6. Газообмен в эритроцитах в крови легких В легких в эритроцит поступает О2 , а из него
- 7. Кривая диссоциации оксигемоглобина Но образование HbО2 не прямо пропорционально уровню РО2 в крови, а идет по
- 8. Биологическое значение указанной кривой В смешанной венозной крови, полученной в покое из правого предсердия, РvО2 =
- 9. Венозный резерв HbО2 И при снижении РvО2 еще до 20 мм рт. ст. в крови остается
- 10. Кривые диссоциации различных видов Hb По активности взаимодействия с Hb: 1 – СО, но СОHb диссоциирует
- 11. Кривые диссоциации НbО2 не постоянны При увеличении в крови рН, снижении СО2 или температуры – кривая
- 12. Газообмен в тканях Доставка О2 к тканям происходит кровотоком - путем конвекции. А в тканях снова
- 13. Количество О2, поступившее к органу, может быть определено по Арт. вен. Разнице содержания О2 (АВР-О2). Зная
- 14. Цилиндр Крога В связи с плохой растворимостью О2 в жидкостях требуется высокие градиенты РО2. Обычно РО2:
- 15. Обычно в большинстве тканей уровень РСО2 близок к 50-60 мм рт. ст. В крови, поступающей в
- 17. Транспорт СО2 В венозной крови содержится около 580 мл/л СО2. Поступивший в эритроциты СО2, при участии
- 18. Для СО2 также имеется зависимость скорости диссоциации, как от уровня РСО2 в крови, так и от
- 19. Реакции, идущие в эритроцитах в венозной крови малого круга кровообращения (в легких) *а) Н++НСО3- H2СО3 Н2О+СО2
- 20. Дыхательные движения выполняются сокращением скелетных мышц, а они иннервируются мотонейронами (С4-Т7) спинного мозга. Поэтому дыхание можно
- 21. Дыхание, являясь вегетативной функцией, но выполняется скелетными мышцами. Поэтому механизмы регуляции его имеют черты, характерные для
- 22. Механизмы, регулирующие дыхание в покое и при физической нагрузке (ФН) несколько отличаются. Это обусловлено: а) при
- 23. Если во внутренних органах (сердце, кишечник) автоматия обусловлена лишь свойствами водителей ритма, то в дыхательном центре
- 24. Дыхательный центр (генератор дыхательного ритма) состоит из: 2 -дорсального ядра (ведущее), 3 – вентрального ядра, 4
- 25. Дыхательные нейроны. Обнаружено 11-типов нейронов, возбуждение в которых можно зарегистрировать во время дыхания. Если они возбуждаются
- 26. Бульбарный отдел дыхательного центра все же обладает и собственной, пейсмекерной автивностью, хотя она и не столь
- 27. Вентральное ядро Дорсальное ядро И Iα Э Межнейронные взаимодействия дыхательного центра /ГЕНЕРАТОР РИТМА ДЫХАНИЯ/ I-α I-β
- 28. Регуляция дыхания в покое Начало возбуждение Iα-нейронов: вдох; выдох : торможение Iα -нейронов возбужденными Iβ-нейронами, (
- 29. Начало – возбуждение Iα-нейронов + возбуждение И-нейронов вентрального центра – глубокий вдох; Форсированный выдох торможение Iα
- 30. Хеморецепторы Центральные хеморецепторы расположены в самом продолговатом мозгу. Они контролируют метаболизм мозга. Периферические в кровеносных сосудах:
- 31. У вентральной поверхности продолговатого мозга на глубине 200-400 мкм расположены центральные хеморецепторы (ЦХР). Нахождение их в
- 32. ПХР находятся в бифуркации общих сонных артерий и в аортальных тельцах, находящихся на верхней и нижней
- 33. Возбуждаются периферические хеморецепторы также при снижении рН или повышении РаСО2. Гипоксия и гиперкапния взаимно усиливают импульсацию
- 34. а) Р. растяжения легких – расположены в гладких мышцах воздухоносных путей (активируя Iβ-нейроны, тормозящие активность Iα-нейронов,
- 35. б) Рецепторы дыхательных мышц . Функционируют по принципу гамма-петли межреберных и мышц стенок живота (при внешнем
- 36. Раздражителями этих рецепторов являются едкие газы Раздражение ирритантных рецепторов вызывает у человека такие неприятные ощущения, как
- 37. Раздражаются они при поступлении биологически активных веществ в сосуды малого круга кровообращения, а так же при
- 39. Скачать презентацию

ДЫХАНИЕ.
Транспорт газов кровью
Газообмен в тканях
Регуляция дыхания
ДЫХАНИЕ.
Транспорт газов кровью
Газообмен в тканях
Регуляция дыхания
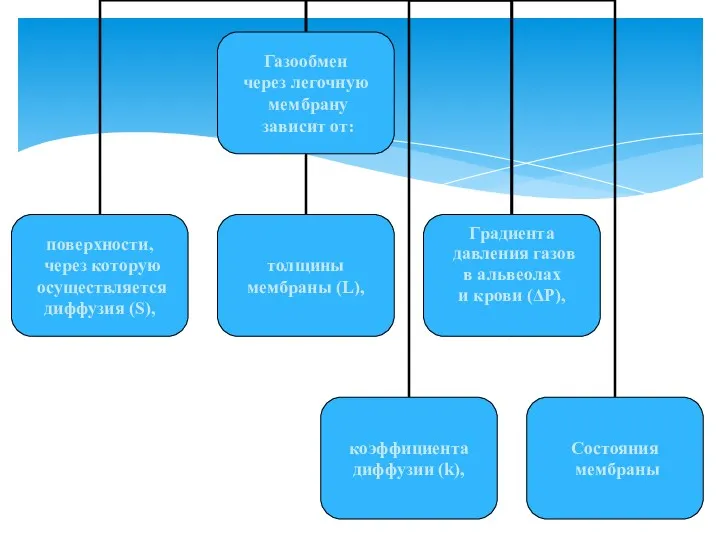

ЗАКОН ФИКА: М = k⋅S/L⋅ΔP:
где, коэффициент диффузии (k)
ЗАКОН ФИКА: М = k⋅S/L⋅ΔP:
где, коэффициент диффузии (k)
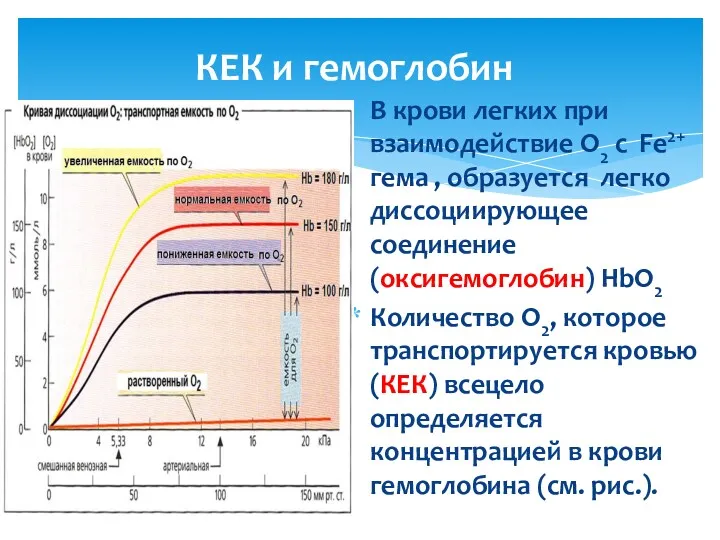
КЕК и гемоглобин
В крови легких при взаимодействие О2 с Fе2+ гема
КЕК и гемоглобин
В крови легких при взаимодействие О2 с Fе2+ гема

Газообмен в эритроцитах в крови легких
В легких в эритроцит поступает О2
Газообмен в эритроцитах в крови легких
В легких в эритроцит поступает О2
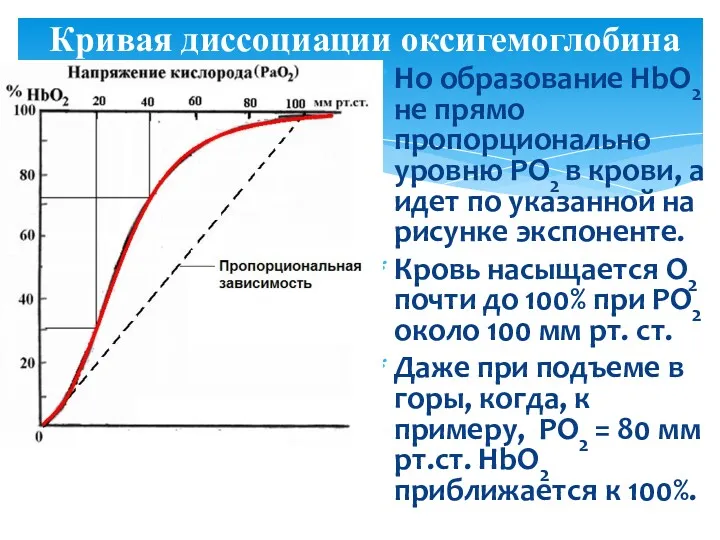
Кривая диссоциации оксигемоглобина
Но образование HbО2 не прямо пропорционально уровню РО2 в
Кривая диссоциации оксигемоглобина
Но образование HbО2 не прямо пропорционально уровню РО2 в

Биологическое значение указанной кривой
В смешанной венозной крови, полученной в покое из
Биологическое значение указанной кривой
В смешанной венозной крови, полученной в покое из

Венозный резерв HbО2
И при снижении РvО2 еще до 20 мм рт.
Венозный резерв HbО2
И при снижении РvО2 еще до 20 мм рт.

Кривые диссоциации различных видов Hb
По активности взаимодействия с Hb:
1 – СО,
Кривые диссоциации различных видов Hb
По активности взаимодействия с Hb:
1 – СО,

Кривые диссоциации НbО2 не постоянны
При увеличении в крови
рН, снижении СО2 или
Кривые диссоциации НbО2 не постоянны
При увеличении в крови
рН, снижении СО2 или
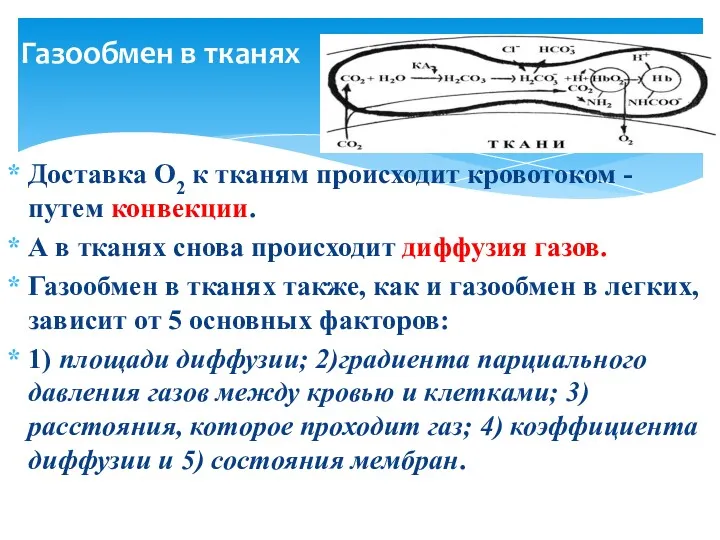
Газообмен в тканях
Доставка О2 к тканям происходит кровотоком - путем конвекции.
Газообмен в тканях
Доставка О2 к тканям происходит кровотоком - путем конвекции.

Количество О2, поступившее к органу, может быть определено по Арт. вен.
Количество О2, поступившее к органу, может быть определено по Арт. вен.

Цилиндр Крога
В связи с плохой растворимостью О2 в жидкостях требуется высокие
Цилиндр Крога
В связи с плохой растворимостью О2 в жидкостях требуется высокие

Обычно в большинстве тканей уровень РСО2 близок к 50-60 мм рт.
Обычно в большинстве тканей уровень РСО2 близок к 50-60 мм рт.


Транспорт СО2
В венозной крови содержится около 580 мл/л СО2. Поступивший в
Транспорт СО2
В венозной крови содержится около 580 мл/л СО2. Поступивший в

Для СО2 также имеется зависимость скорости диссоциации, как от уровня РСО2
Для СО2 также имеется зависимость скорости диссоциации, как от уровня РСО2

Реакции, идущие в эритроцитах в венозной крови малого круга кровообращения (в
Реакции, идущие в эритроцитах в венозной крови малого круга кровообращения (в

Дыхательные движения выполняются сокращением скелетных мышц, а они иннервируются мотонейронами (С4-Т7)
Дыхательные движения выполняются сокращением скелетных мышц, а они иннервируются мотонейронами (С4-Т7)

Дыхание, являясь вегетативной функцией, но выполняется скелетными мышцами. Поэтому механизмы регуляции
Дыхание, являясь вегетативной функцией, но выполняется скелетными мышцами. Поэтому механизмы регуляции

Механизмы, регулирующие дыхание в покое и при физической нагрузке (ФН) несколько
Механизмы, регулирующие дыхание в покое и при физической нагрузке (ФН) несколько

Если во внутренних органах (сердце, кишечник) автоматия обусловлена лишь свойствами водителей
Если во внутренних органах (сердце, кишечник) автоматия обусловлена лишь свойствами водителей

Дыхательный центр (генератор дыхательного ритма) состоит из:
2 -дорсального ядра (ведущее),
3
Дыхательный центр (генератор дыхательного ритма) состоит из:
2 -дорсального ядра (ведущее),
3

Дыхательные нейроны.
Обнаружено 11-типов нейронов, возбуждение в которых можно зарегистрировать во время
Дыхательные нейроны.
Обнаружено 11-типов нейронов, возбуждение в которых можно зарегистрировать во время

Бульбарный отдел дыхательного центра все же обладает и собственной, пейсмекерной автивностью,
Бульбарный отдел дыхательного центра все же обладает и собственной, пейсмекерной автивностью,
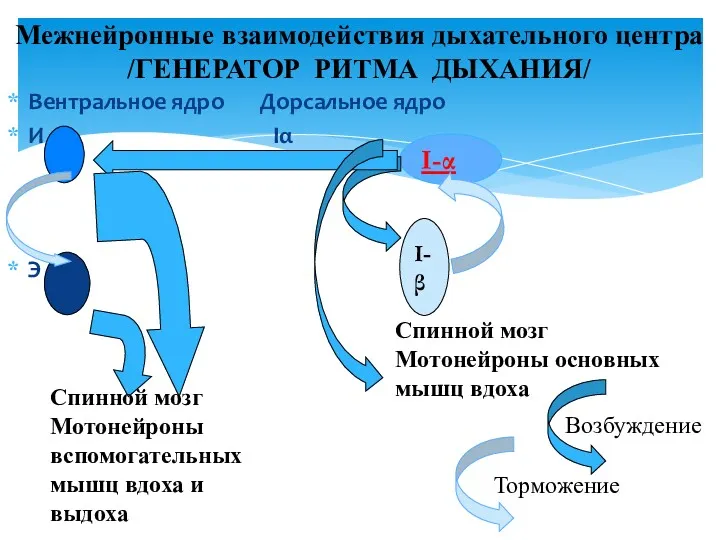
Вентральное ядро Дорсальное ядро
И Iα
Э
Межнейронные взаимодействия дыхательного центра
/ГЕНЕРАТОР РИТМА ДЫХАНИЯ/
I-α
I-β
Спинной
Вентральное ядро Дорсальное ядро
И Iα
Э
Межнейронные взаимодействия дыхательного центра
/ГЕНЕРАТОР РИТМА ДЫХАНИЯ/
I-α
I-β
Спинной

Регуляция дыхания в покое
Начало возбуждение Iα-нейронов: вдох;
выдох : торможение Iα -нейронов
Регуляция дыхания в покое
Начало возбуждение Iα-нейронов: вдох;
выдох : торможение Iα -нейронов

Начало – возбуждение Iα-нейронов + возбуждение И-нейронов вентрального центра – глубокий

Хеморецепторы
Центральные хеморецепторы расположены в самом продолговатом мозгу. Они контролируют метаболизм мозга.
Периферические
Хеморецепторы
Центральные хеморецепторы расположены в самом продолговатом мозгу. Они контролируют метаболизм мозга.
Периферические

У вентральной поверхности продолговатого мозга на глубине 200-400 мкм расположены центральные
У вентральной поверхности продолговатого мозга на глубине 200-400 мкм расположены центральные

ПХР находятся в бифуркации общих сонных артерий и в аортальных тельцах,
ПХР находятся в бифуркации общих сонных артерий и в аортальных тельцах,

Возбуждаются периферические хеморецепторы также при снижении рН или повышении РаСО2. Гипоксия
Возбуждаются периферические хеморецепторы также при снижении рН или повышении РаСО2. Гипоксия

а) Р. растяжения легких – расположены в гладких мышцах воздухоносных путей
а) Р. растяжения легких – расположены в гладких мышцах воздухоносных путей

б) Рецепторы дыхательных мышц . Функционируют по принципу гамма-петли межреберных и
б) Рецепторы дыхательных мышц . Функционируют по принципу гамма-петли межреберных и

Раздражителями этих рецепторов являются едкие газы Раздражение ирритантных рецепторов вызывает у
Раздражителями этих рецепторов являются едкие газы Раздражение ирритантных рецепторов вызывает у

Раздражаются они при поступлении биологически активных веществ в сосуды малого круга
Раздражаются они при поступлении биологически активных веществ в сосуды малого круга
 Белки. Содержание белков в различных тканях человека
Белки. Содержание белков в различных тканях человека Гендік инженерия
Гендік инженерия Растительный мир Кольского полуострова. Редкие, исчезающие и занесенные в Красную книгу виды
Растительный мир Кольского полуострова. Редкие, исчезающие и занесенные в Красную книгу виды Гетерофункциональные органические соединения. Основные классы и особенности реакционной способности. (Лекция 3)
Гетерофункциональные органические соединения. Основные классы и особенности реакционной способности. (Лекция 3) Цікаві факти про тварин. Незвичайні рослини світу
Цікаві факти про тварин. Незвичайні рослини світу Интересные растения
Интересные растения Живые организмы. Признаки живых организмов
Живые организмы. Признаки живых организмов Размножение организмов
Размножение организмов Класс Млекопитающие. Внешнее строение
Класс Млекопитающие. Внешнее строение Дыхание растений. Взаимосвязь процессов дыхания и фотосинтеза
Дыхание растений. Взаимосвязь процессов дыхания и фотосинтеза Післязародковий розвиток організмів
Післязародковий розвиток організмів Мутация (3)
Мутация (3) Мышцы груди и живота
Мышцы груди и живота Основы гистологии. Виды тканей
Основы гистологии. Виды тканей Сера и её роль в организме человека
Сера и её роль в организме человека Общие вопросы анатомии и физиологии аппарата движения человека. Лекция № 5
Общие вопросы анатомии и физиологии аппарата движения человека. Лекция № 5 Хто такі комахи? Урок №118. Я досліджую світ
Хто такі комахи? Урок №118. Я досліджую світ Қамыс өсімдігіне жалпы сипаттама
Қамыс өсімдігіне жалпы сипаттама Значение витамина С в питании. БАД компании NSP
Значение витамина С в питании. БАД компании NSP Растительные ткани
Растительные ткани презентация Самые высокие горы
презентация Самые высокие горы Синтез белков в клетке
Синтез белков в клетке Культивирование отдельных (одиночных) клеток
Культивирование отдельных (одиночных) клеток Макроэволюция органического мира
Макроэволюция органического мира Проект Животные Красной книги Московской области г. Долгопрудного
Проект Животные Красной книги Московской области г. Долгопрудного Общая характеристика Голосеменных
Общая характеристика Голосеменных Хищники
Хищники Правнук царского дуба. Проект по окружающему миру
Правнук царского дуба. Проект по окружающему миру