- Ультраструктура бактериальной клетки

Содержание
- 2. Особенности организации прокариотической клетки Обязательными компонентами бактериальной клетки являются: Клеточная стенка Цитоплазматическая мембрана Цитоплазма с органоидами
- 3. Клеточная стенка бактерий (КС) КС обеспечивает постоянную форму клетки, механическую и осмотическую защиту, взаимосвязь с окружающей
- 4. Клеточная стенка бактерий (КС) Основу клеточной стенки всех бактерий составляет специфический гетерополимер -пептидогликан, обеспечивающий ригидность и
- 5. Синтез клеточной стенки Образование мономера – дисахарид-пептидного звена Перенос образовавшегося мономера из цитоплазмы на наружную поверхность
- 6. Пептидогликан представляет собой трехмерную сеть из поперечно сшитых полисахаридных цепей, которая легко пропускает различные низкомолекулярные и
- 8. Клеточная стенка грам + бактерий Липотейховые к-т пептидогликан тейховые к-т Цитоплазматическая мембрана Клеточная стенка грамположительных бактерий
- 9. Клеточная стенка грамположительных бактерий Тейхоевые кислоты характеризуются большой вариабельностью строения могут связываться с клеточными мембранами животных
- 11. Клеточная стенка грамотрицательных бактерий устроена намного сложнее. Пептидогликан образует только тонкий внутренний слой клеточной стенки, неплотно
- 12. ЛПС построен из липида А, который собственно формирует наружный листок мембраны, олигосахаридного ядра, обычно содержащего заряженные
- 13. Пептидогликан отделен от ЦПМ и наружной мембраны периплазматическим пространством – специализированная метаболически активная область или компартмент
- 15. Кислотоустойчивый тип клеточной стенки Для многих представителей порядка Corynebacteriales характерен особый тип строения клеточной стенки –
- 16. Клеточная стенка бактерий. L-формы Если бактерии частично или полностью утратили клеточную стенку, но сохранили способность к
- 17. S -слой (от англ. surface - поверхность): сравнительно тонкая оболочка, образованная белками, способными к самоорганизации (самосборке)
- 18. Поверхностные структуры бактериальной клетки Капсула - слизистая структура толщиной более 0,2мкм, прочно связанная с клеточной стенкой
- 19. Поверхностные структуры бактериальной клетки Капсулы имеют консистенцию геля и плохо удерживают краситель, для их выявления чаще
- 20. Поверхностные структуры бактериальной клетки Функции капсул и слизей: защитная (предохраняют клетку от механических повреждений, высыхания и
- 21. Цитоплазматическая мембрана (ЦПМ) ЦПМ – обязательный структурный элемент любой клетки, нарушение целостности которого приводит к потере
- 22. барьерная (избирательный перенос молекул и ионов); синтетическая (в ней локализованы ферменты, катализирующие конечные этапы синтеза мембранных
- 23. Проницаемость ЦПМ
- 24. Figure 12.15. Проницаемость бислойной фосфолипидной мембраны Газы, гидрофобные молекулы (бензол), малые полярные не заряженные молекулы могут
- 25. Транспортные системы Выделяют 4 типа транспортных систем, с участием которых происходит проникновение молекул в бактериальную клетку:
- 26. Жгутики бактерий поверхностные структуры, определяющие способность клетки к движению в жидкой среде. их число, размеры, расположение
- 28. Основную массу жгутика составляет длинная спирально закрученная нить (фибрилла), у поверхности клеточной стенки переходящая в утолщенную
- 29. Размеры жгутиков (толщина 10-20 нм, длина 3-15 мкм) не позволяют их увидеть в обычном световом микроскопе
- 30. Ворсинки (фимбрии и пили) Белковые образования на поверхности ряда бактерий При электронной микроскопии они выглядят как
- 31. Пили Пили типа 1 - common pili (пили общего типа, ответственные за адгезию) - прочно связаны
- 32. Амилоидные белки поверхности микроорганизмов Амилоиды (т. е. вещество, сходное с крахмалом — amylum) — особый способ
- 33. Амилоидные белки поверхности микроорганизмов Многие микроорганизмы имеют на своей поверхности амилоидные белки. Тонкие аггрегативные фимбрии (Thin
- 34. Курли Курли E. coli связываются со многими белками человека, в частности с фибронектином, ламинином, коллагеном I
- 35. Эпителиальные клетки кишечника и иммунная система «узнают» курли Salmonella and E. coli - обитатели кишечника, чьи
- 37. Цитоплазма -содержимое клетки, окруженное ЦПМ Состоит из цитозоля и органоидов (нуклеоид, рибосомы) и включений Цитозоль –
- 38. Рибосомы бактерий Рибосома представляет собой компактную частицу специфической формы, лишенную внутренней и внешней симметрии Состоят из
- 39. Основная морфологическая черта электронно-микроскопических изображений рибосомы - борозда, разделяющая две рибосомные субчастицы Эта борозда сильно расширяется
- 40. Функционально рибосома - это молекулярная машина, протягивающая вдоль себя цепь мРНК, считывающая закодированную в мРНК генетическую
- 41. В процессе элонгации рибосома удерживает мРНК и движется относительно ее (или протягивает ее сквозь себя) в
- 42. Синтез белка. Отличия от эукариот У прокариот транскрипция и трансляция идеально сопряжены при помощи механизма управления
- 43. Центральная догма биологии
- 44. Генетический аппарат бактерий Модель конденсации бактериальной хромосомы Основной генетический аппарат бактерий- нуклеоид- представлен одной хромосомой (гаплоидный
- 45. Отличительной особенностью организации генетического аппарата прокариот является то, что помимо нуклеоида, в цитоплазме могут находиться особого
- 46. У некоторых групп микроорганизмов организация хромосом и плазмид значительно отличается от классической. Например, представители родов Borrelia
- 47. ДНК ДНК-биополимер, мономерами которого являются нуклеотиды Первичная структура- линейная последовательность дезоксирибонуклеотидов в одной цепочке определяет все
- 48. Транскрипция ДНК в ходе деления клеток начинается с разделения двух цепей, каждая из которых становится матрицей,
- 49. Хеликаза, топоизомераза и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК.
- 50. Каждый фрагмент Оказаки начинается с короткой РНК-затравки (праймера), необходимой для функционирования ДНК-полимеразы. Праймер синтезируется специальной РНК-полимеразой
- 51. Цитоплазматические включения активно функционирующие структуры (структуры, содержащие пигменты у фотосинтезирующих бактерий) продукты клеточного метаболизма, не выделяющиеся
- 52. Для прокариот, метаболизм которых связан с соединениями серы, характерно отложение в клетках молекулярной серы. Сера накапливается,
- 53. Споры бактерий Споры бактерий можно рассматривать как форму сохранения наследственной информации бактериальной клетки в неблагоприятных условиях
- 54. Спорообразование Споробразование начинается при изменении температуры, влажности, недостатке питательных веществ или накоплении продуктов обмена Процесс начинается
- 55. Спорообразование В ходе созревания будущая спора подвергается обезвоживанию – содержание воды в цитоплазме падает до 25-55%,
- 56. Эндоспоры Эндоспоры в теле клетки может располагаться: 1. центрально — возбудитель сибирской язвы Вacillus anthracis; 2.
- 57. Прорастание споры Активация споры: прорастание споры активируется при прогревании активируют прорастание глюкоза и другие углеводы ,
- 58. Другие покоящиеся формы (цисты) Многие неспорообразующие бактерии, в том числе и патогенные микроорганизмы, в определенных условиях
- 60. Скачать презентацию
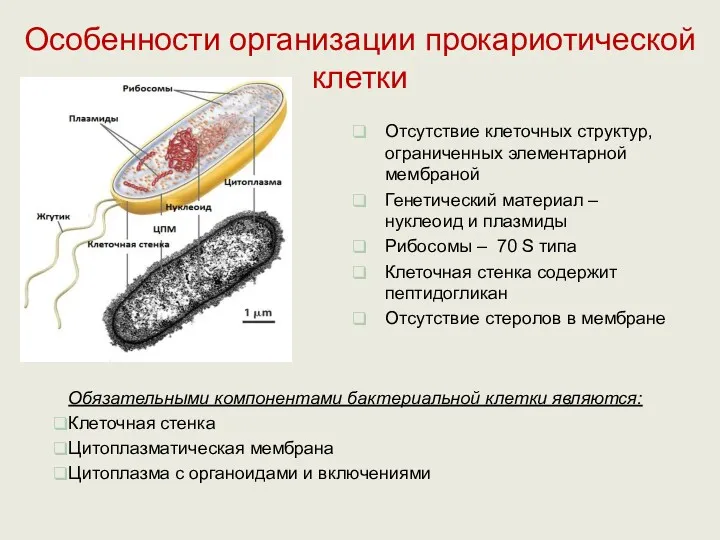
Особенности организации прокариотической клетки
Обязательными компонентами бактериальной клетки являются:
Клеточная стенка
Цитоплазматическая мембрана
Цитоплазма с
Особенности организации прокариотической клетки
Обязательными компонентами бактериальной клетки являются:
Клеточная стенка
Цитоплазматическая мембрана
Цитоплазма с

Клеточная стенка бактерий (КС)
КС обеспечивает постоянную форму клетки, механическую и осмотическую
Клеточная стенка бактерий (КС)
КС обеспечивает постоянную форму клетки, механическую и осмотическую

Клеточная стенка бактерий (КС)
Основу клеточной стенки всех бактерий составляет специфический гетерополимер
Клеточная стенка бактерий (КС)
Основу клеточной стенки всех бактерий составляет специфический гетерополимер
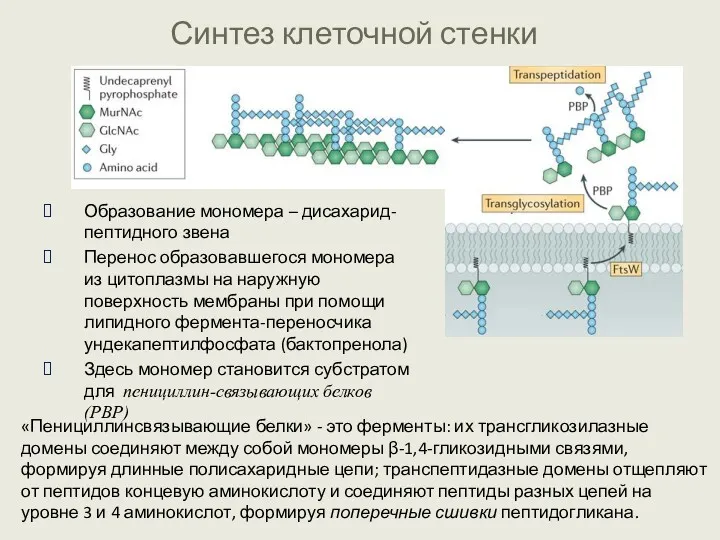
Синтез клеточной стенки
Образование мономера – дисахарид-пептидного звена
Перенос образовавшегося мономера из цитоплазмы
Синтез клеточной стенки
Образование мономера – дисахарид-пептидного звена
Перенос образовавшегося мономера из цитоплазмы
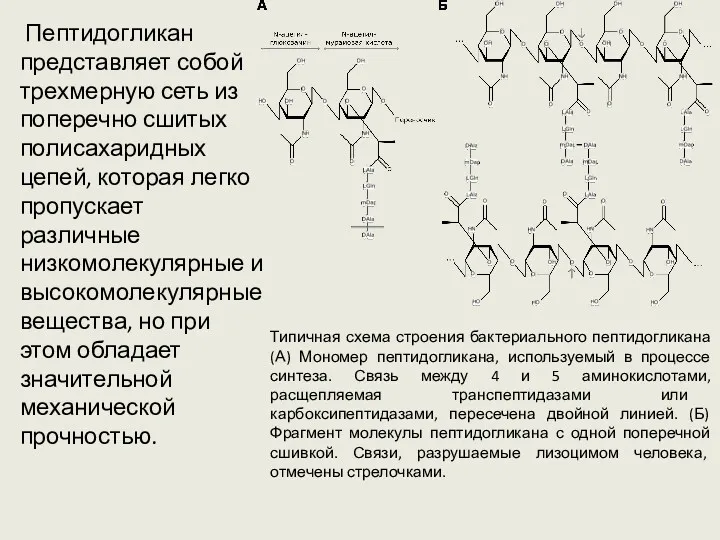
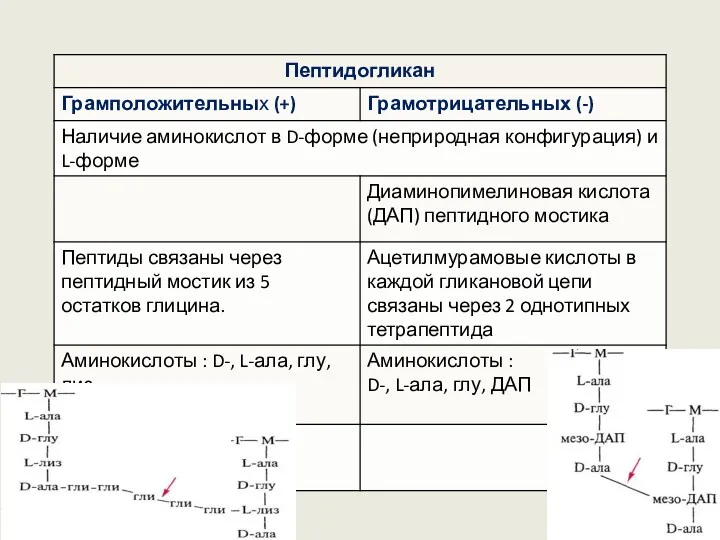
Пептидогликан представляет собой трехмерную сеть из поперечно сшитых полисахаридных цепей,
Пептидогликан представляет собой трехмерную сеть из поперечно сшитых полисахаридных цепей,
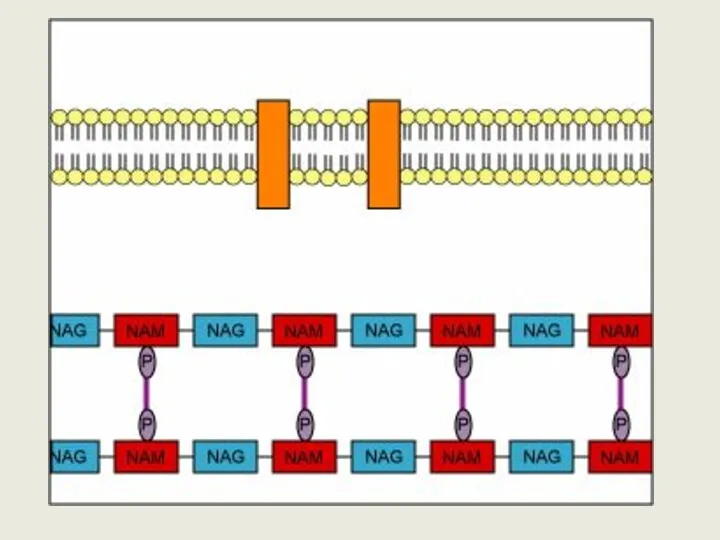
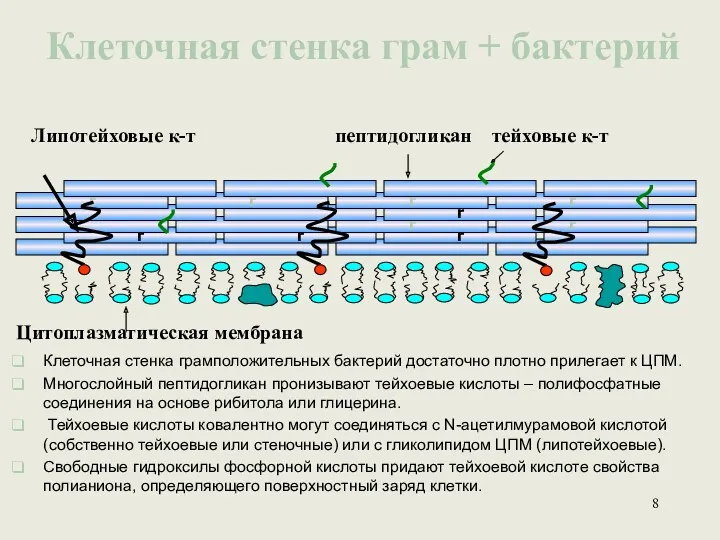
Клеточная стенка грам + бактерий
Липотейховые к-т
пептидогликан тейховые к-т
Цитоплазматическая мембрана
Клеточная стенка грамположительных
Клеточная стенка грам + бактерий
Липотейховые к-т
пептидогликан тейховые к-т
Цитоплазматическая мембрана
Клеточная стенка грамположительных
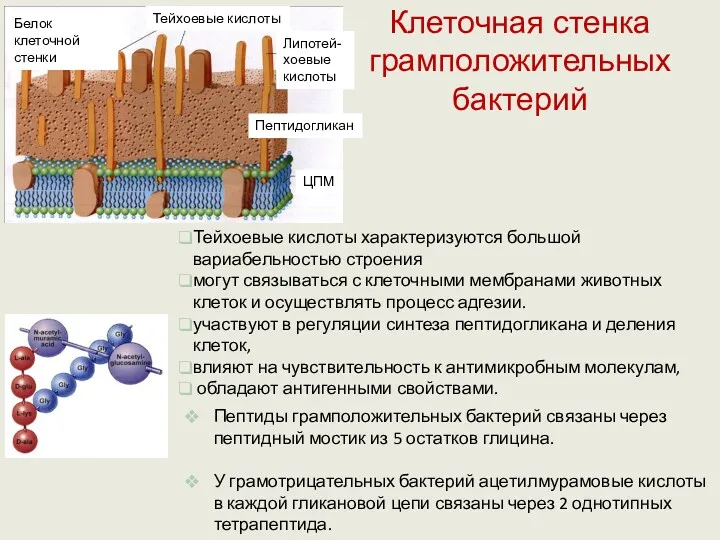
Клеточная стенка грамположительных бактерий
Тейхоевые кислоты характеризуются большой вариабельностью строения
могут связываться
Клеточная стенка грамположительных бактерий
Тейхоевые кислоты характеризуются большой вариабельностью строения
могут связываться
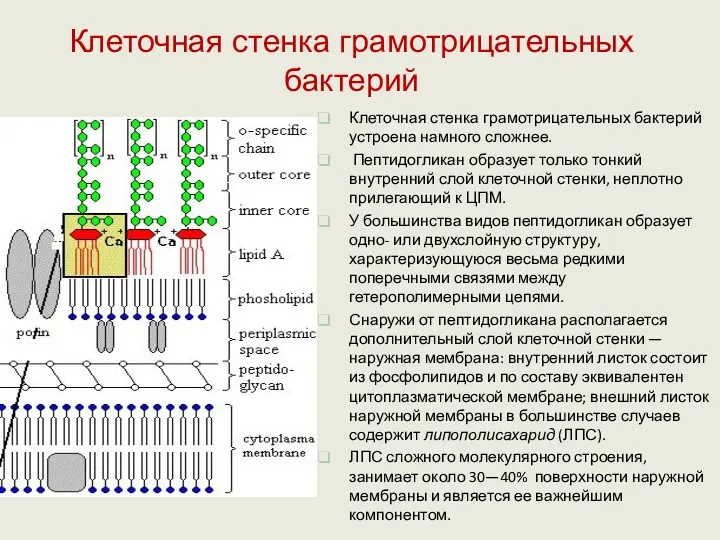
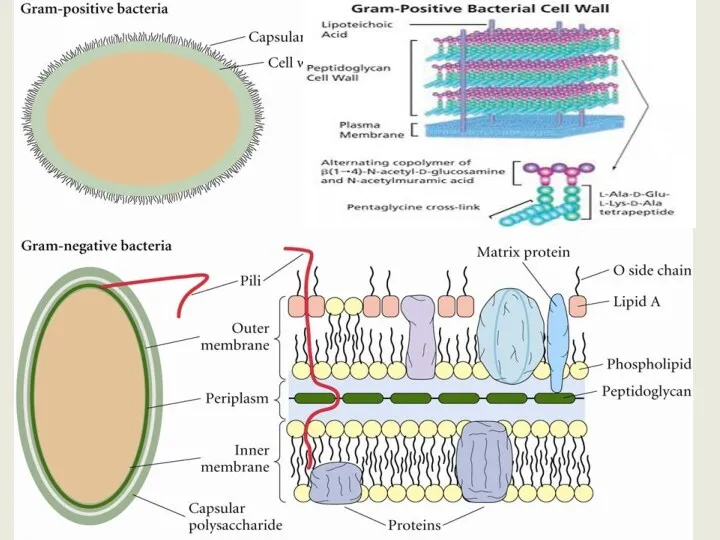
Клеточная стенка грамотрицательных бактерий устроена намного сложнее.
Пептидогликан образует только тонкий
Клеточная стенка грамотрицательных бактерий устроена намного сложнее.
Пептидогликан образует только тонкий
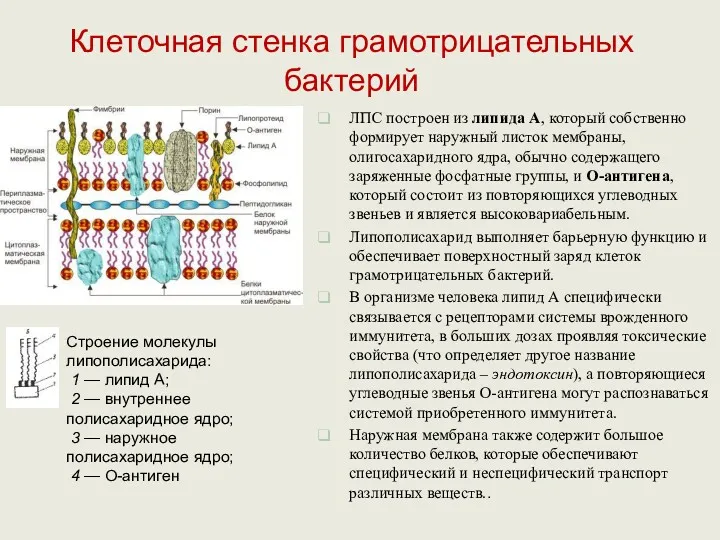
ЛПС построен из липида А, который собственно формирует наружный листок мембраны,
ЛПС построен из липида А, который собственно формирует наружный листок мембраны,

Пептидогликан отделен от ЦПМ и наружной мембраны периплазматическим пространством – специализированная
Пептидогликан отделен от ЦПМ и наружной мембраны периплазматическим пространством – специализированная
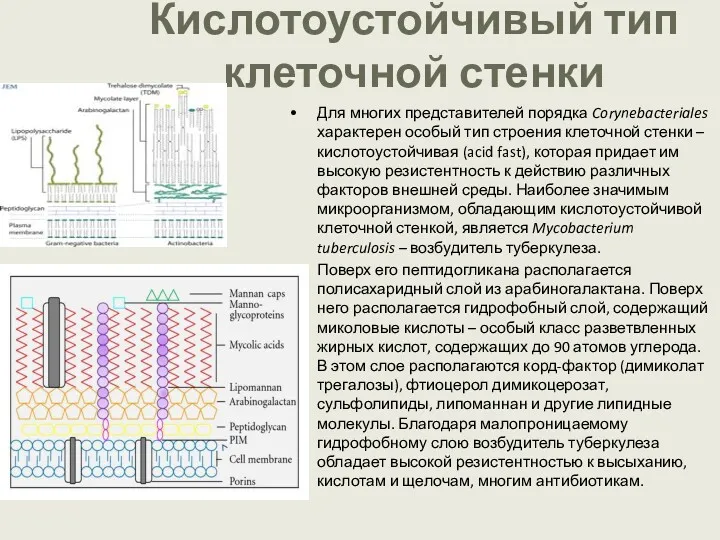
Кислотоустойчивый тип клеточной стенки
Для многих представителей порядка Corynebacteriales характерен особый тип
Кислотоустойчивый тип клеточной стенки
Для многих представителей порядка Corynebacteriales характерен особый тип
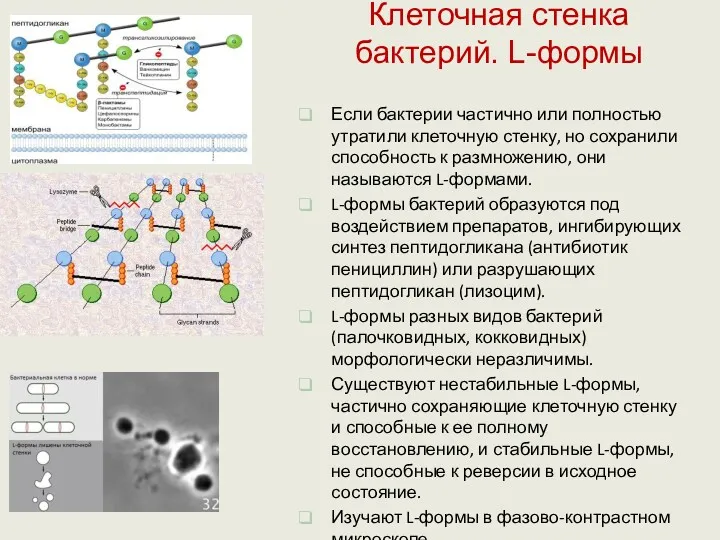
Клеточная стенка бактерий. L-формы
Если бактерии частично или полностью утратили клеточную стенку,
Клеточная стенка бактерий. L-формы
Если бактерии частично или полностью утратили клеточную стенку,

S -слой (от англ. surface - поверхность): сравнительно тонкая оболочка, образованная белками,
S -слой (от англ. surface - поверхность): сравнительно тонкая оболочка, образованная белками,
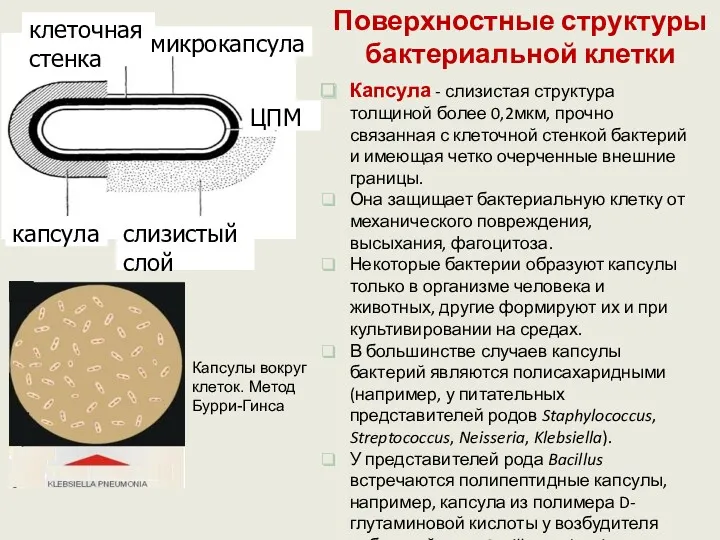
Поверхностные структуры бактериальной клетки
Капсула - слизистая структура толщиной более 0,2мкм, прочно
Поверхностные структуры бактериальной клетки
Капсула - слизистая структура толщиной более 0,2мкм, прочно
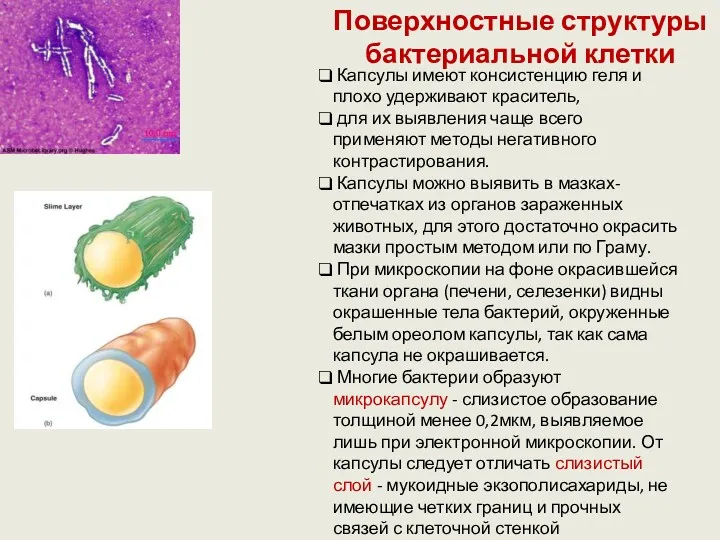
Поверхностные структуры бактериальной клетки
Капсулы имеют консистенцию геля и плохо удерживают
Поверхностные структуры бактериальной клетки
Капсулы имеют консистенцию геля и плохо удерживают

Поверхностные структуры бактериальной клетки
Функции капсул и слизей:
защитная (предохраняют
Поверхностные структуры бактериальной клетки
Функции капсул и слизей:
защитная (предохраняют

Цитоплазматическая мембрана (ЦПМ)
ЦПМ – обязательный структурный элемент любой клетки, нарушение целостности
Цитоплазматическая мембрана (ЦПМ)
ЦПМ – обязательный структурный элемент любой клетки, нарушение целостности

барьерная (избирательный перенос молекул и ионов);
синтетическая (в ней локализованы ферменты,
барьерная (избирательный перенос молекул и ионов);
синтетическая (в ней локализованы ферменты,
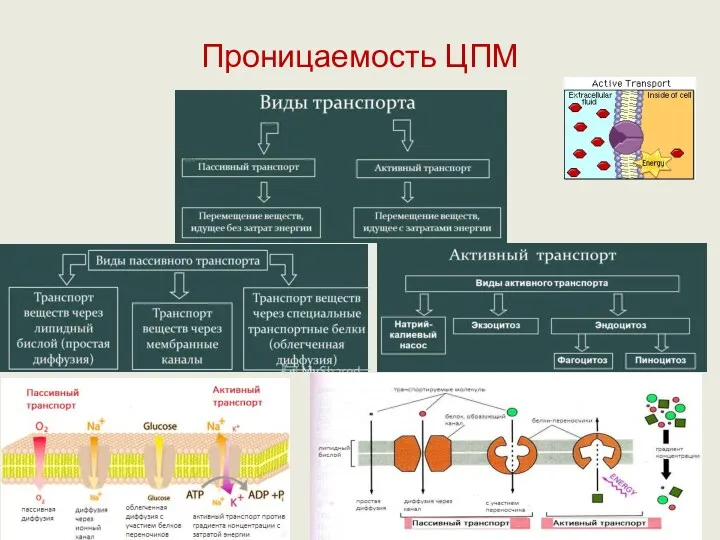
Проницаемость ЦПМ
Проницаемость ЦПМ
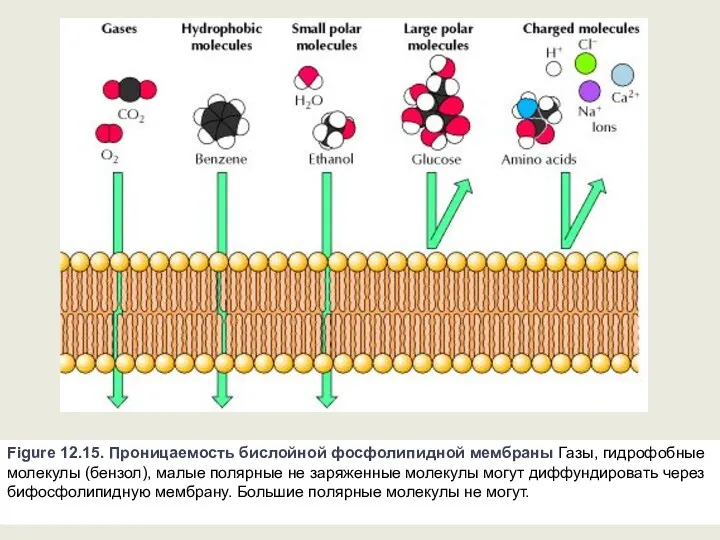
Figure 12.15. Проницаемость бислойной фосфолипидной мембраны Газы, гидрофобные молекулы (бензол), малые
Figure 12.15. Проницаемость бислойной фосфолипидной мембраны Газы, гидрофобные молекулы (бензол), малые

Транспортные системы
Выделяют 4 типа транспортных систем, с участием которых происходит проникновение
Транспортные системы
Выделяют 4 типа транспортных систем, с участием которых происходит проникновение

Жгутики бактерий
поверхностные структуры, определяющие способность клетки к движению в жидкой
Жгутики бактерий
поверхностные структуры, определяющие способность клетки к движению в жидкой
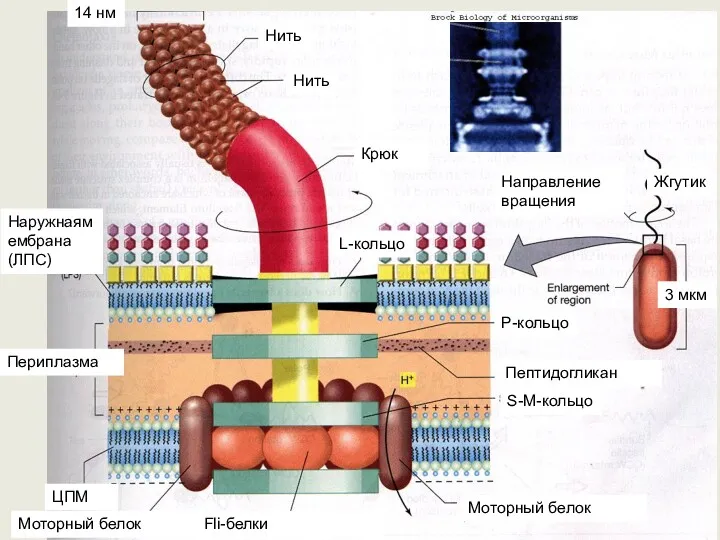

Основную массу жгутика составляет длинная спирально закрученная нить (фибрилла), у поверхности
Основную массу жгутика составляет длинная спирально закрученная нить (фибрилла), у поверхности

Размеры жгутиков (толщина 10-20 нм, длина 3-15 мкм) не позволяют их
Размеры жгутиков (толщина 10-20 нм, длина 3-15 мкм) не позволяют их

Ворсинки (фимбрии и пили)
Белковые образования на поверхности ряда бактерий
При электронной
Ворсинки (фимбрии и пили)
Белковые образования на поверхности ряда бактерий
При электронной

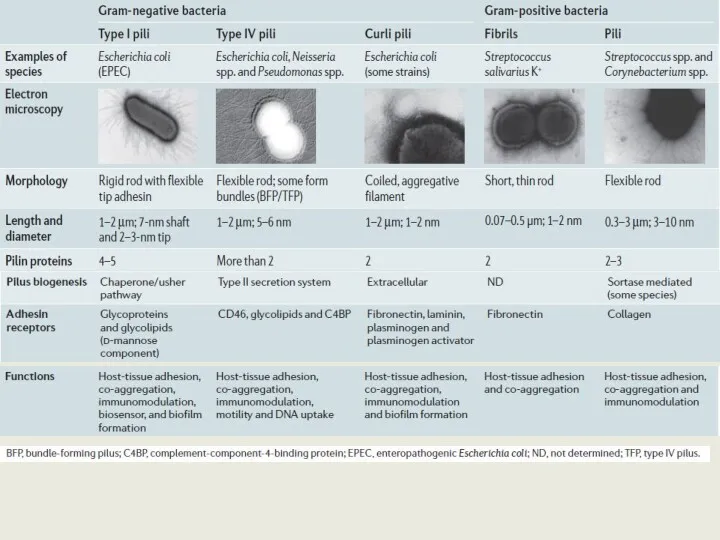
Пили
Пили типа 1 - common pili (пили общего типа, ответственные за
Пили
Пили типа 1 - common pili (пили общего типа, ответственные за
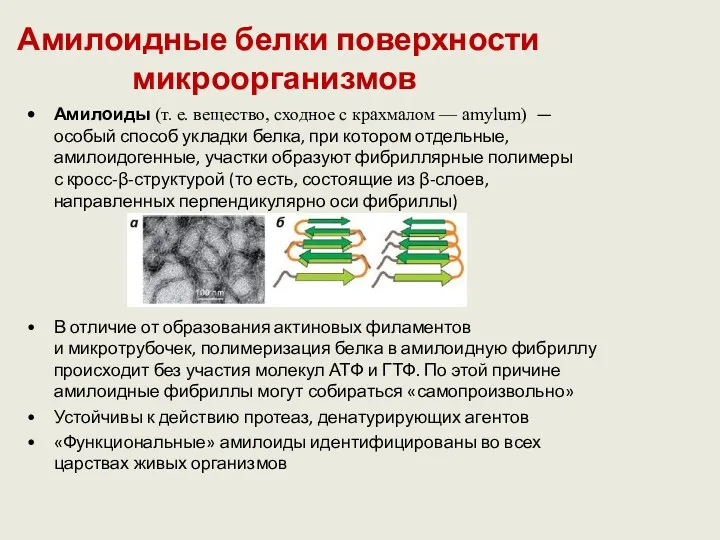
Амилоидные белки поверхности микроорганизмов
Амилоиды (т. е. вещество, сходное с крахмалом — amylum) — особый способ укладки белка,
Амилоидные белки поверхности микроорганизмов
Амилоиды (т. е. вещество, сходное с крахмалом — amylum) — особый способ укладки белка,

Амилоидные белки поверхности микроорганизмов
Многие микроорганизмы имеют на своей поверхности амилоидные
Амилоидные белки поверхности микроорганизмов
Многие микроорганизмы имеют на своей поверхности амилоидные

Курли
Курли E. coli связываются со многими белками человека, в частности с
Курли
Курли E. coli связываются со многими белками человека, в частности с
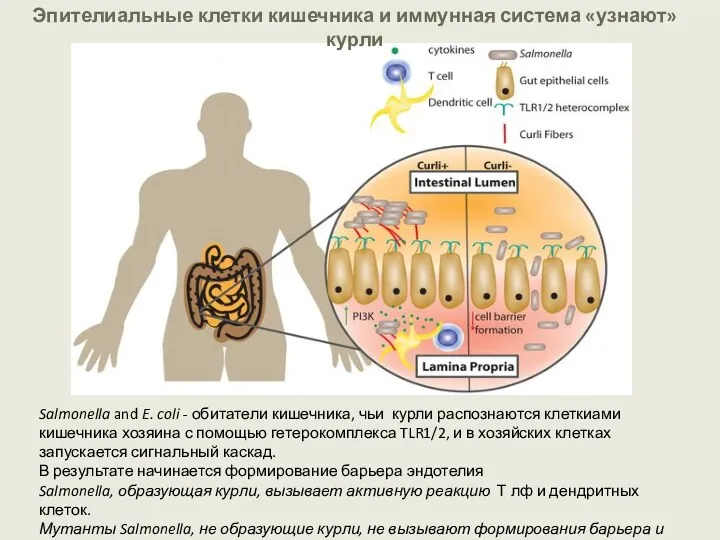
Эпителиальные клетки кишечника и иммунная система «узнают» курли
Salmonella and E. coli - обитатели кишечника,
Эпителиальные клетки кишечника и иммунная система «узнают» курли
Salmonella and E. coli - обитатели кишечника,

Цитоплазма
-содержимое клетки, окруженное ЦПМ
Состоит из цитозоля и органоидов (нуклеоид, рибосомы) и
Цитоплазма
-содержимое клетки, окруженное ЦПМ
Состоит из цитозоля и органоидов (нуклеоид, рибосомы) и

Рибосомы бактерий
Рибосома представляет собой компактную частицу специфической формы, лишенную внутренней и
Рибосомы бактерий
Рибосома представляет собой компактную частицу специфической формы, лишенную внутренней и
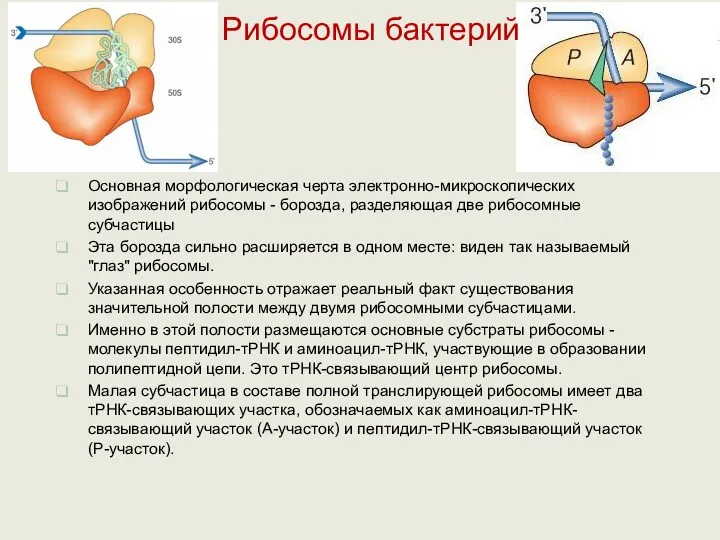
Основная морфологическая черта электронно-микроскопических изображений рибосомы - борозда, разделяющая две рибосомные
Основная морфологическая черта электронно-микроскопических изображений рибосомы - борозда, разделяющая две рибосомные
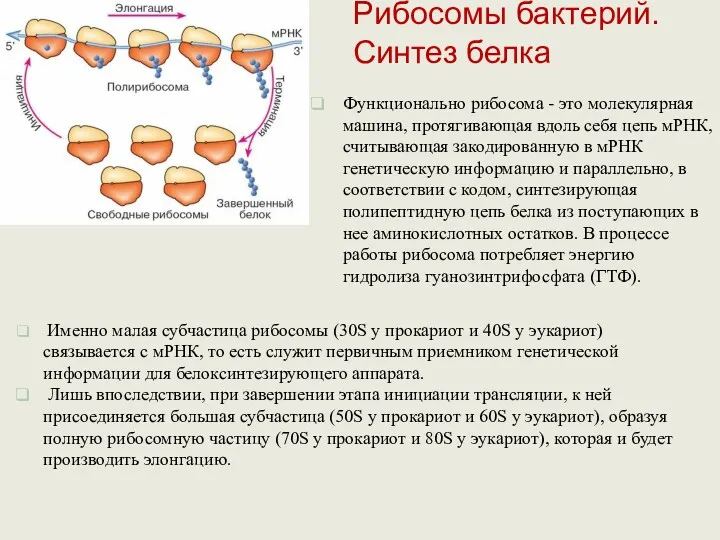
Функционально рибосома - это молекулярная машина, протягивающая вдоль себя цепь мРНК,
Функционально рибосома - это молекулярная машина, протягивающая вдоль себя цепь мРНК,

В процессе элонгации рибосома удерживает мРНК и движется относительно ее
В процессе элонгации рибосома удерживает мРНК и движется относительно ее

Синтез белка. Отличия от эукариот
У прокариот транскрипция и трансляция идеально сопряжены
Синтез белка. Отличия от эукариот
У прокариот транскрипция и трансляция идеально сопряжены
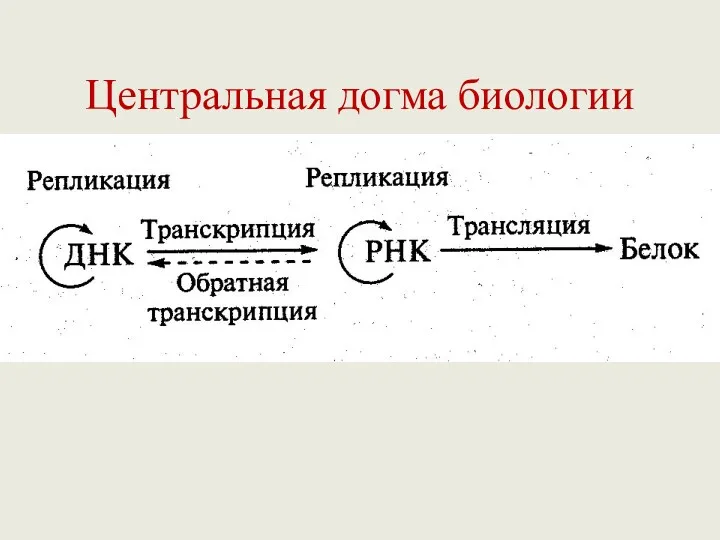
Центральная догма биологии
Центральная догма биологии

Генетический аппарат бактерий
Модель конденсации бактериальной хромосомы
Основной генетический аппарат бактерий- нуклеоид- представлен
Генетический аппарат бактерий
Модель конденсации бактериальной хромосомы
Основной генетический аппарат бактерий- нуклеоид- представлен
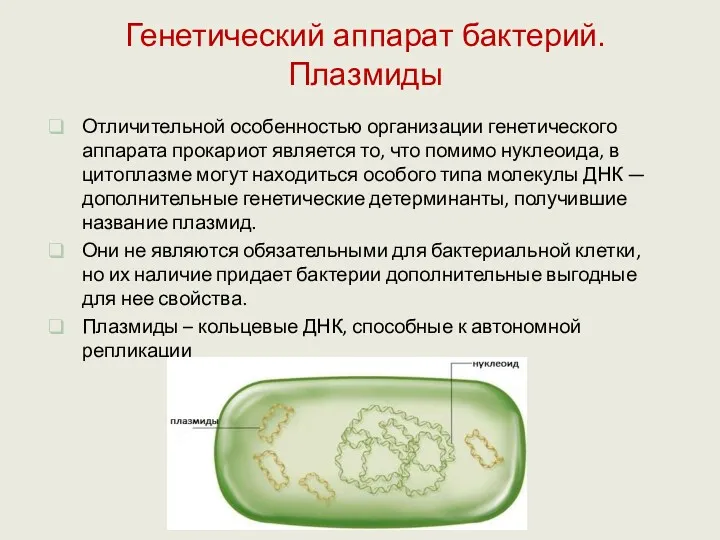
Отличительной особенностью организации генетического аппарата прокариот является то, что помимо нуклеоида,
Отличительной особенностью организации генетического аппарата прокариот является то, что помимо нуклеоида,
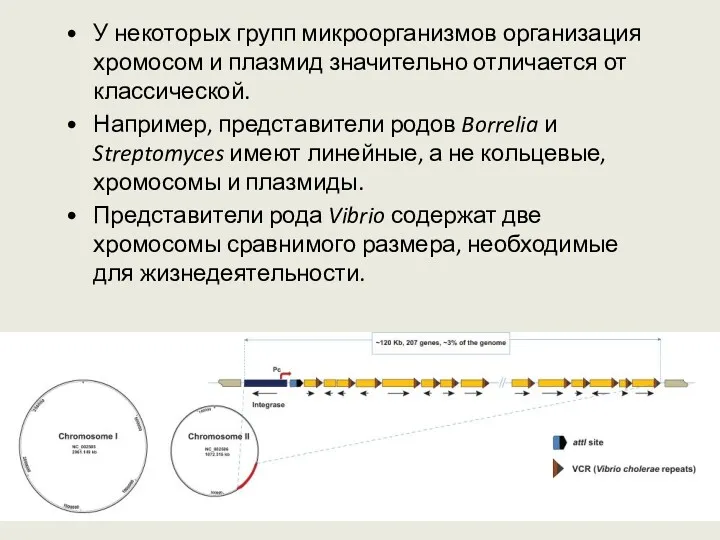
У некоторых групп микроорганизмов организация хромосом и плазмид значительно отличается от
У некоторых групп микроорганизмов организация хромосом и плазмид значительно отличается от
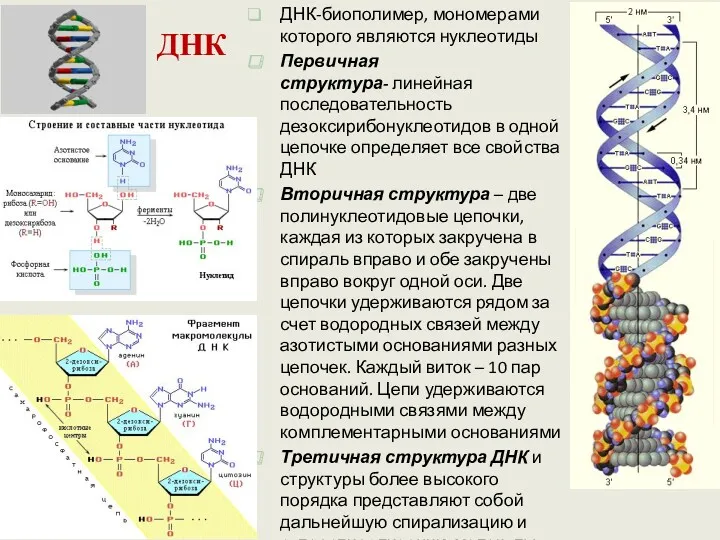
ДНК
ДНК-биополимер, мономерами которого являются нуклеотиды
Первичная структура- линейная последовательность дезоксирибонуклеотидов в одной цепочке
ДНК
ДНК-биополимер, мономерами которого являются нуклеотиды
Первичная структура- линейная последовательность дезоксирибонуклеотидов в одной цепочке

Транскрипция ДНК в ходе деления клеток начинается с разделения двух цепей,
Транскрипция ДНК в ходе деления клеток начинается с разделения двух цепей,

Хеликаза, топоизомераза и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом
Хеликаза, топоизомераза и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом

Каждый фрагмент Оказаки начинается с короткой РНК-затравки (праймера), необходимой для функционирования
Каждый фрагмент Оказаки начинается с короткой РНК-затравки (праймера), необходимой для функционирования

Цитоплазматические включения
активно функционирующие структуры (структуры, содержащие пигменты у фотосинтезирующих бактерий)
продукты клеточного
Цитоплазматические включения
активно функционирующие структуры (структуры, содержащие пигменты у фотосинтезирующих бактерий)
продукты клеточного

Для прокариот, метаболизм которых связан с соединениями серы, характерно отложение в
Для прокариот, метаболизм которых связан с соединениями серы, характерно отложение в

Споры бактерий
Споры бактерий можно рассматривать как форму сохранения наследственной информации бактериальной
Споры бактерий
Споры бактерий можно рассматривать как форму сохранения наследственной информации бактериальной
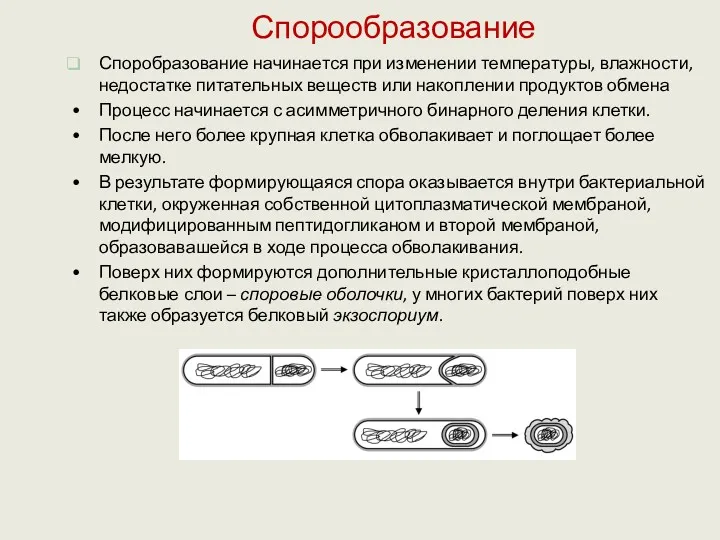
Спорообразование
Споробразование начинается при изменении температуры, влажности, недостатке питательных веществ или накоплении
Спорообразование
Споробразование начинается при изменении температуры, влажности, недостатке питательных веществ или накоплении

Спорообразование
В ходе созревания будущая спора подвергается обезвоживанию – содержание воды в
Спорообразование
В ходе созревания будущая спора подвергается обезвоживанию – содержание воды в
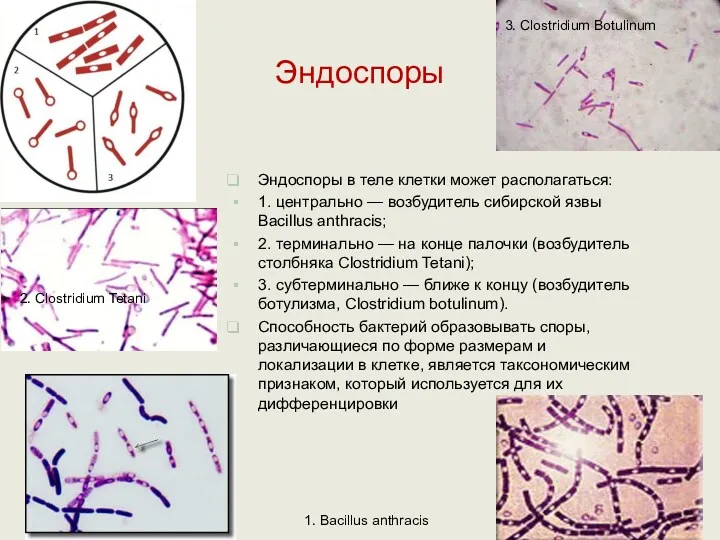
Эндоспоры
Эндоспоры в теле клетки может располагаться:
1. центрально — возбудитель сибирской
Эндоспоры
Эндоспоры в теле клетки может располагаться:
1. центрально — возбудитель сибирской

Прорастание споры
Активация споры:
прорастание споры активируется при прогревании
активируют прорастание глюкоза и
Прорастание споры
Активация споры:
прорастание споры активируется при прогревании
активируют прорастание глюкоза и

Другие покоящиеся формы (цисты)
Многие неспорообразующие бактерии, в том числе и патогенные
Другие покоящиеся формы (цисты)
Многие неспорообразующие бактерии, в том числе и патогенные
 Методы исследования биологии
Методы исследования биологии Ферменты в генетической инженерии. Тема 2
Ферменты в генетической инженерии. Тема 2 Внешнее строение листа
Внешнее строение листа Клиническая физиология КЩС
Клиническая физиология КЩС Происхождение человека
Происхождение человека Цветок – орган генеративного размножения
Цветок – орган генеративного размножения General Musculoskeletal Screening: Upper Extremities
General Musculoskeletal Screening: Upper Extremities Эволюционные теории и взгляды
Эволюционные теории и взгляды Систематика растений
Систематика растений Дуб - царь деревьев
Дуб - царь деревьев Эпизоотологическое и эпидемиологическое значение грызунов. Дератизация
Эпизоотологическое и эпидемиологическое значение грызунов. Дератизация Значение водорослей в природе и жизни человека
Значение водорослей в природе и жизни человека презентация к уроку по теме: Цветок. Строение и значение цветка
презентация к уроку по теме: Цветок. Строение и значение цветка Антропогенез. Гипотезы возникновения человека. Сходство и различия человека и животных. Часть 4
Антропогенез. Гипотезы возникновения человека. Сходство и различия человека и животных. Часть 4 Живі фільтри
Живі фільтри Слух. Среднее и внутреннее ухо. Функции среднего уха. Строение улитки
Слух. Среднее и внутреннее ухо. Функции среднего уха. Строение улитки Анатомия и физиология ЦНС. Свойства и принципы функционирования нервных центров. Строение СМ. Цереброспинальная жидкость
Анатомия и физиология ЦНС. Свойства и принципы функционирования нервных центров. Строение СМ. Цереброспинальная жидкость Ферменты,структура и механизм действия. Классификация и номенклатура. (Лекция 4)
Ферменты,структура и механизм действия. Классификация и номенклатура. (Лекция 4) Практика по общей биологии
Практика по общей биологии Простейшие – эукариотические одноклеточные микроорганизмы
Простейшие – эукариотические одноклеточные микроорганизмы Главные направления эволюции органического мира
Главные направления эволюции органического мира Желудочная секреция
Желудочная секреция Строение, химический состав и жизнедеятельность растительной клетки
Строение, химический состав и жизнедеятельность растительной клетки презентация по биологии Экологическая задача Ч.Дарвина
презентация по биологии Экологическая задача Ч.Дарвина Nervové mechanismy regulace dýchání
Nervové mechanismy regulace dýchání Минеральные вещества
Минеральные вещества Что такое хвоинки
Что такое хвоинки Глюконеогенез. Регуляция гликолиза и глюконеогенеза. Цикл кори. Пентозофосфатный путь превращения глюкозы. Спиртовое брожение
Глюконеогенез. Регуляция гликолиза и глюконеогенеза. Цикл кори. Пентозофосфатный путь превращения глюкозы. Спиртовое брожение