- Вирусология. История вирусологии

Содержание
- 3. История вирусологии 1892 Д.И.Ивановский представил работу, в которой показал, что экстракт из зараженного растения табака после
- 4. Ивановский Д.И.
- 5. История вирусологии 1898г. Ф. Лефлер и П.Фрош был открыт вирус ящера, первый вирус животного 1915г. ФредерикТуорт
- 6. История вирусологии 1935 Уэндел Стенли получил очищенный препарат ВТМ, предположив, что он является глобулярным белком 1937
- 7. История вирусологии 1940г Бернет вводит экспериментальную модель - куриный эмбрион. 1949г. Эндерс, Роббинс – разработали модель
- 8. Вирусы Вирусы – неклеточная форма существования живой материи, обладающая собственным геномом, которые являются абсолютными паразитами на
- 9. Вирусы 1. Отсутствие клеточного строения 2. Отсутствие собственной белок синтезирующей системы 3. Один тип нуклеиновой кислоты
- 10. Формы существования 1. Внутри клетки - вегетативная 2. Вне клетки - форма ВИРИОНА
- 11. Строение вириона 1. ПРОСТООРГАНИЗОВАНЫЕ: а. нуклеиновая кислота б. капсид 2. СЛОЖНООРГАНИЗОВАННЫЕ а. нуклеиновая кислота б. капсид
- 14. Капсид 1.Устойчивость к протеолитическим ферментам (особенность укладки) 2.Защита от нуклеаз 3. Способность к самосборке В состав
- 15. Оболочка (суперкапсид) структура 1. Двойной фосфолипидный слой, являющейся производным ядерной или цитоплазматической мембраны клетки 2. Вируспецифические
- 17. Сложноорганизованный вирион
- 18. Функции гликопротеидов 1. белки прикрепления к клеточным рецепторам 2. ферментативная активность (лецитиназа, нейраминидаза) принимают участие в
- 19. Резистентность вирусов Степень устойчивости вирусов к факторам окружающей среды зависит от типа вирионов Наименее устойчивы сложноорганизованные
- 20. Резистентность вирусов 1. Хорошо переносят низкие температуры. Сохраняются в замороженном состоянии 2. Большинство вирусов чувствительны к
- 21. Культивирование вирусов
- 22. Классификация вирусов Вирусы классифицируются в соответстствии строения их генома и структурой вириона Царство Vira Семейство viridae
- 23. Взаимодействие вируса и клетки 1. Интегративная инфекция 2. Продуктивная инфекция 3. Абортивная инфекция
- 24. Интегративная инфекция 1. Вирусы , обладающие двуцепочечной ДНК кольцевой формы (Papillomaviridae, Polyomaviridae ) 2. Вирусы, обладающие
- 25. Интегративная инфекция
- 26. Интегративная инфекция
- 27. Интегративная инфекция 1. Вирус переходит в состояние провируса 2. Новое поколение вирионов не формируется 3. Происходит
- 28. Продуктивная инфекция 1. Прикрепление и пенетрация 2. Раздевание 3. Реализация вирусного генома: а. образование иРНК б.
- 29. Продуктивная инфекция Проникновение вируса в клетку является энергозависимым процессом, поэтому вирусы проникают только в метаболически активные
- 30. Продуктивная инфекция ПРОНИКНОВЕНИЕ: 1. Эндоцитоз с образованием внутриклеточной вакуоли 2. Слияние липидной оболочки вируса с цитоплазматической
- 31. Продуктивная инфекция Раздевание вируса (декапсидация): 1. Удаление вирусного капсида 2. Освобождение вирусного генома Происходит непосредственно в
- 32. Продуктивная инфекция После проникновения в клетку вирусу необходимо: 1. синтезировать иРНК, которые смогут транслироваться с образованием
- 33. Продуктивная инфекция 3. Обеспечить упаковку вирусного генома в вирион и выход из клетки дочерних вирусных частиц
- 34. Стратегия репликации ДНК-содержащих вирусов ДНК - содержащие вирусы ( за исключением вируса натуральной оспы)размножаются в ядре,
- 36. Двуцепочечная ДНК ( ds) ДНК иРНК белок ДНК-зависимая РНК полимераза
- 37. Двуцепочечная ДНК ( ds) Образовавшиеся иРНК транспортируется в цитоплазму и транслируется с образованием ранних и вирусных
- 38. Одноцепочечная ДНК (ss) 1. ДНК(+) ДНК(-) Синтез комплементарной нити 2. ДНК(-) иРНК белок ДНК-зависимая РНК полимераза
- 39. Однонитевые ДНК Репликация происходит в ядре Вначале происходит синтез комплементарной нити ДНК и образование двунитевой комплементарной
- 40. РНК-содержащие вирусы Не используют аппарат клетки для репликации генома В состав вириона входят их собственные РНК-зависимая
- 41. РНК (+) РНК(+) белок РНК-зависимая РНК полимераза РНК(-) Репликативная форма +/- РНК РНК(+)
- 42. РНК(-) РНК(-) РНК(+) белок вирусная РНК транскриптаза (матрица для образования дочерних геномных РНК(-) ) Вирусы с
- 43. Двуцепочечные РНК (ds) Двуцепочечная РНК (ds) не может функционировать как иРНК, поэтому нуждается в упаковке в
- 44. Двуцепочечные РНК (ds) (ds) РНК иРНК белок РНК транскриптаза вируса Эти вирусы (Reoviridae) имеют сегментированный геном,
- 45. d+(диплоидные)РНК 1. РНК(+) ДНК ревертаза ДНК встраивается в геном провирус 2. ДНК иРНК белок ДНК-зависимая РНК
- 46. Ретроидные вирусы ДНК +/- (ДНК-полимераза с ревертазной активностью) ДНК(-) прегеномная РНК + субгеномные РНК ДНК-зависимая РНК
- 47. Сборка вирионов Участок сборки вириона зависит от места репликации вирусного генома и способа выхода из клетки
- 48. Выход вируса из клетки Простоорганизованные вирусы выходят из клетки путем лизиса Вирусы, покрытые липидной оболочкой, покидают
- 49. ЦПД в культуре клеток
- 51. Бактериофаги
- 53. Бактериофаги
- 56. Взаимодействие фага и бактерии
- 57. Фаговая конверсия Приобретение бактериями новых свойств в результате инфекции умеренным фагом
- 59. Применение бактериофагов
- 61. Скачать презентацию


История вирусологии
1892 Д.И.Ивановский представил работу, в которой показал, что экстракт из
История вирусологии
1892 Д.И.Ивановский представил работу, в которой показал, что экстракт из

Ивановский Д.И.
Ивановский Д.И.

История вирусологии
1898г. Ф. Лефлер и П.Фрош был открыт вирус ящера, первый
История вирусологии
1898г. Ф. Лефлер и П.Фрош был открыт вирус ящера, первый

История вирусологии
1935 Уэндел Стенли получил очищенный препарат ВТМ, предположив, что он
История вирусологии
1935 Уэндел Стенли получил очищенный препарат ВТМ, предположив, что он

История вирусологии
1940г Бернет вводит экспериментальную модель - куриный эмбрион.
1949г. Эндерс, Роббинс
История вирусологии
1940г Бернет вводит экспериментальную модель - куриный эмбрион.
1949г. Эндерс, Роббинс

Вирусы
Вирусы – неклеточная форма существования живой материи, обладающая собственным геномом, которые
Вирусы
Вирусы – неклеточная форма существования живой материи, обладающая собственным геномом, которые

Вирусы
1. Отсутствие клеточного строения
2. Отсутствие собственной белок синтезирующей системы
3. Один тип
Вирусы
1. Отсутствие клеточного строения
2. Отсутствие собственной белок синтезирующей системы
3. Один тип

Формы существования
1. Внутри клетки - вегетативная
2. Вне клетки - форма ВИРИОНА
Формы существования
1. Внутри клетки - вегетативная
2. Вне клетки - форма ВИРИОНА

Строение вириона
1. ПРОСТООРГАНИЗОВАНЫЕ:
а. нуклеиновая кислота
б. капсид
2. СЛОЖНООРГАНИЗОВАННЫЕ
а. нуклеиновая
Строение вириона
1. ПРОСТООРГАНИЗОВАНЫЕ:
а. нуклеиновая кислота
б. капсид
2. СЛОЖНООРГАНИЗОВАННЫЕ
а. нуклеиновая
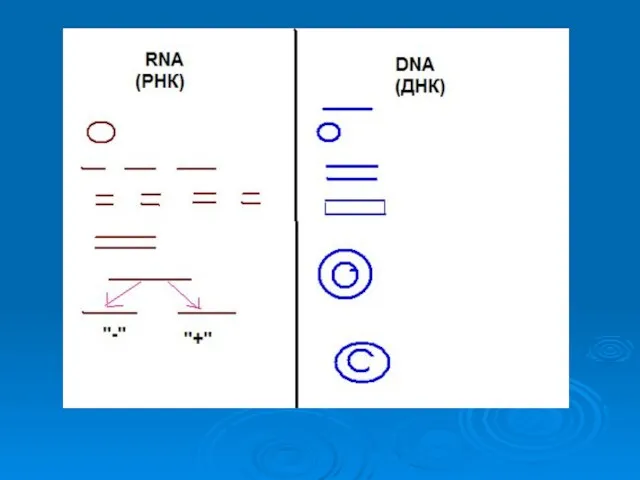
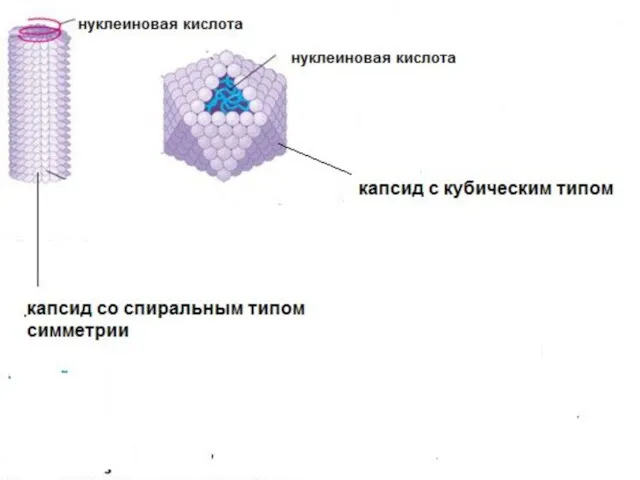

Капсид
1.Устойчивость к протеолитическим ферментам (особенность укладки)
2.Защита от нуклеаз
3. Способность к самосборке
В
Капсид
1.Устойчивость к протеолитическим ферментам (особенность укладки)
2.Защита от нуклеаз
3. Способность к самосборке
В

Оболочка (суперкапсид)
структура
1. Двойной фосфолипидный слой, являющейся производным ядерной или цитоплазматической мембраны
Оболочка (суперкапсид)
структура
1. Двойной фосфолипидный слой, являющейся производным ядерной или цитоплазматической мембраны
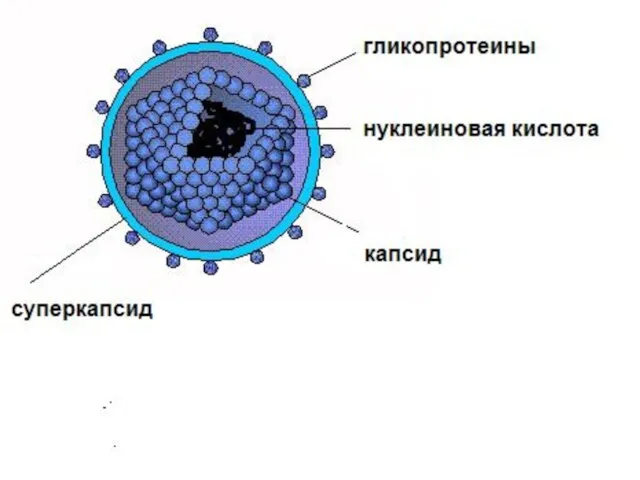
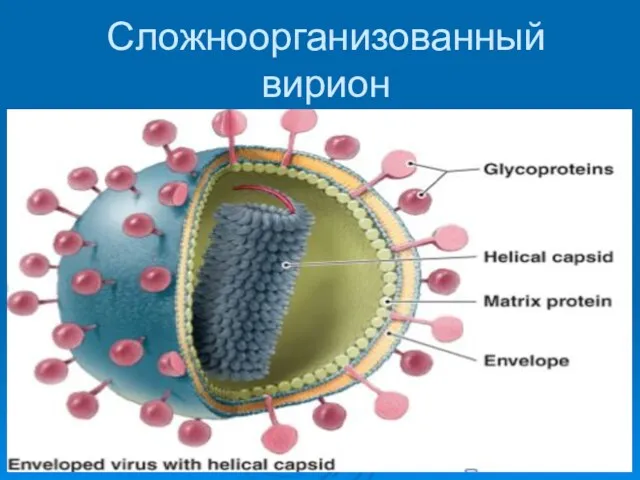
Сложноорганизованный вирион
Сложноорганизованный вирион

Функции гликопротеидов
1. белки прикрепления к клеточным рецепторам
2. ферментативная активность (лецитиназа, нейраминидаза)
Функции гликопротеидов
1. белки прикрепления к клеточным рецепторам
2. ферментативная активность (лецитиназа, нейраминидаза)

Резистентность вирусов
Степень устойчивости вирусов к факторам окружающей среды зависит от типа
Резистентность вирусов
Степень устойчивости вирусов к факторам окружающей среды зависит от типа

Резистентность вирусов
1. Хорошо переносят низкие температуры. Сохраняются в замороженном состоянии
2. Большинство
Резистентность вирусов
1. Хорошо переносят низкие температуры. Сохраняются в замороженном состоянии
2. Большинство
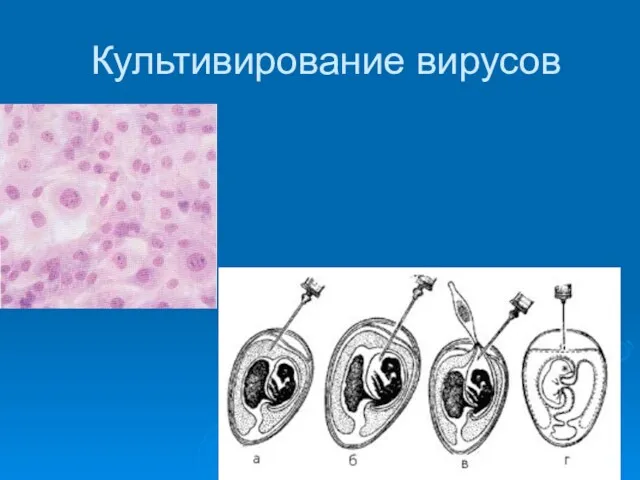
Культивирование вирусов
Культивирование вирусов

Классификация вирусов
Вирусы классифицируются в соответстствии строения их генома и структурой вириона
Царство
Классификация вирусов
Вирусы классифицируются в соответстствии строения их генома и структурой вириона
Царство

Взаимодействие вируса и клетки
1. Интегративная инфекция
2. Продуктивная инфекция
3. Абортивная инфекция
Взаимодействие вируса и клетки
1. Интегративная инфекция
2. Продуктивная инфекция
3. Абортивная инфекция

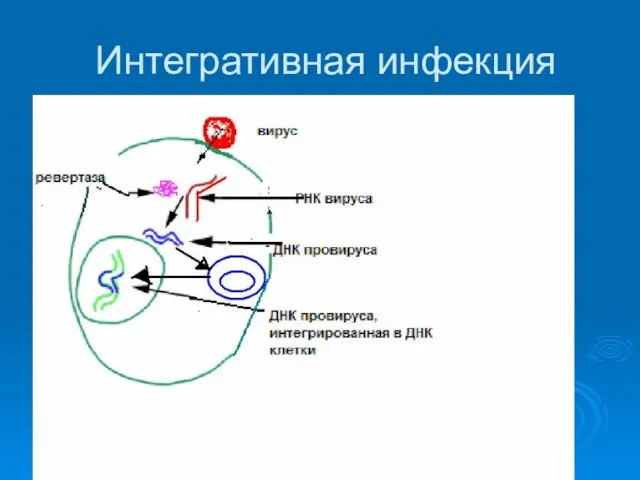
Интегративная инфекция
1. Вирусы , обладающие двуцепочечной ДНК кольцевой формы (Papillomaviridae, Polyomaviridae
Интегративная инфекция
1. Вирусы , обладающие двуцепочечной ДНК кольцевой формы (Papillomaviridae, Polyomaviridae
Интегративная инфекция
Интегративная инфекция
Интегративная инфекция
Интегративная инфекция

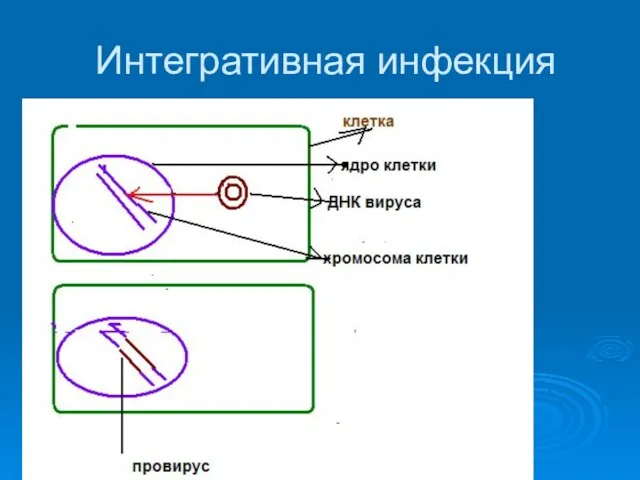
Интегративная инфекция
1. Вирус переходит в состояние провируса
2. Новое поколение вирионов не
Интегративная инфекция
1. Вирус переходит в состояние провируса
2. Новое поколение вирионов не

Продуктивная инфекция
1. Прикрепление и пенетрация
2. Раздевание
3. Реализация вирусного генома:
а. образование
Продуктивная инфекция
1. Прикрепление и пенетрация
2. Раздевание
3. Реализация вирусного генома:
а. образование

Продуктивная инфекция
Проникновение вируса в клетку является энергозависимым процессом, поэтому вирусы проникают
Продуктивная инфекция
Проникновение вируса в клетку является энергозависимым процессом, поэтому вирусы проникают

Продуктивная инфекция
ПРОНИКНОВЕНИЕ:
1. Эндоцитоз с образованием внутриклеточной вакуоли
2. Слияние липидной оболочки вируса
Продуктивная инфекция
ПРОНИКНОВЕНИЕ:
1. Эндоцитоз с образованием внутриклеточной вакуоли
2. Слияние липидной оболочки вируса

Продуктивная инфекция
Раздевание вируса (декапсидация):
1. Удаление вирусного капсида
2. Освобождение вирусного генома
Происходит
Продуктивная инфекция
Раздевание вируса (декапсидация):
1. Удаление вирусного капсида
2. Освобождение вирусного генома
Происходит

Продуктивная инфекция
После проникновения в клетку вирусу необходимо:
1. синтезировать иРНК, которые смогут
Продуктивная инфекция
После проникновения в клетку вирусу необходимо:
1. синтезировать иРНК, которые смогут

Продуктивная инфекция
3. Обеспечить упаковку вирусного генома в вирион и выход из
Продуктивная инфекция
3. Обеспечить упаковку вирусного генома в вирион и выход из

Стратегия репликации ДНК-содержащих вирусов
ДНК - содержащие вирусы ( за исключением вируса
Стратегия репликации ДНК-содержащих вирусов
ДНК - содержащие вирусы ( за исключением вируса
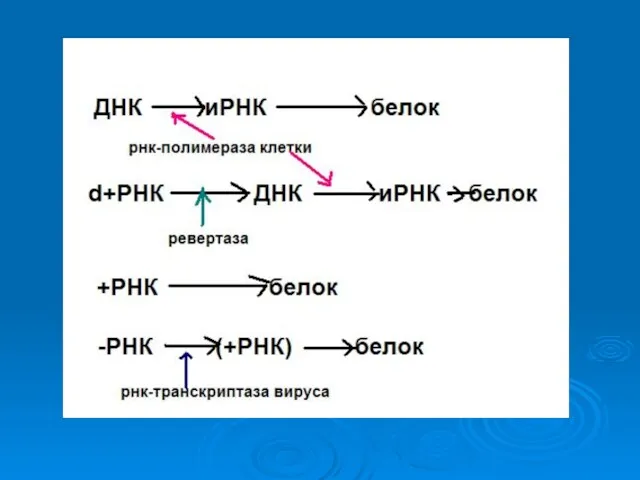
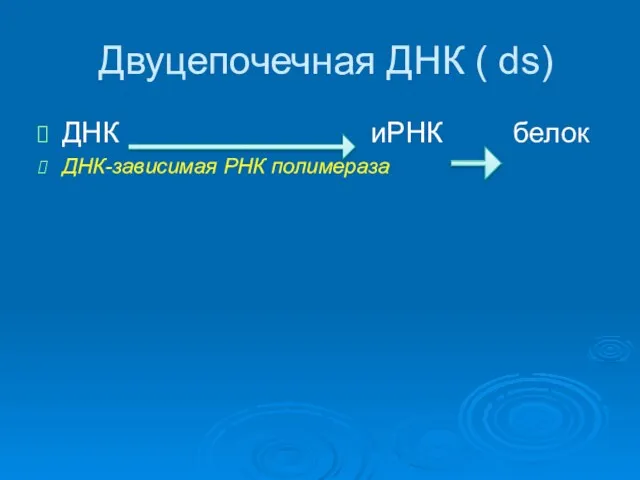
Двуцепочечная ДНК ( ds)
ДНК иРНК белок
ДНК-зависимая РНК полимераза
Двуцепочечная ДНК ( ds)
ДНК иРНК белок
ДНК-зависимая РНК полимераза

Двуцепочечная ДНК ( ds)
Образовавшиеся иРНК транспортируется в цитоплазму и транслируется с
Двуцепочечная ДНК ( ds)
Образовавшиеся иРНК транспортируется в цитоплазму и транслируется с
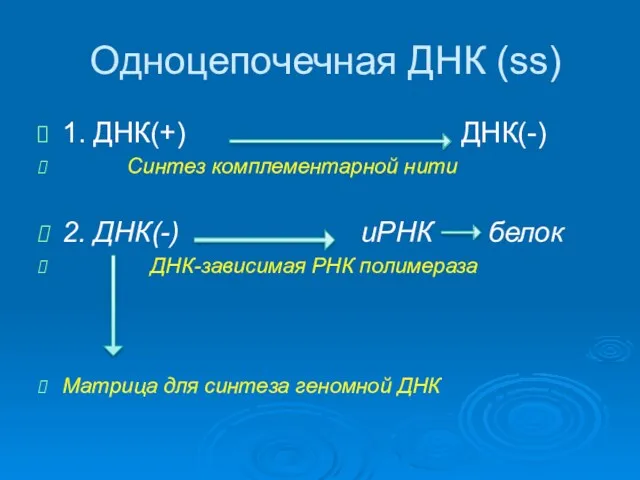
Одноцепочечная ДНК (ss)
1. ДНК(+) ДНК(-)
Синтез комплементарной нити
2. ДНК(-) иРНК белок
Одноцепочечная ДНК (ss)
1. ДНК(+) ДНК(-)
Синтез комплементарной нити
2. ДНК(-) иРНК белок

Однонитевые ДНК
Репликация происходит в ядре
Вначале происходит синтез комплементарной нити ДНК и
Однонитевые ДНК
Репликация происходит в ядре
Вначале происходит синтез комплементарной нити ДНК и

РНК-содержащие вирусы
Не используют аппарат клетки для репликации генома
В состав вириона входят
РНК-содержащие вирусы
Не используют аппарат клетки для репликации генома
В состав вириона входят
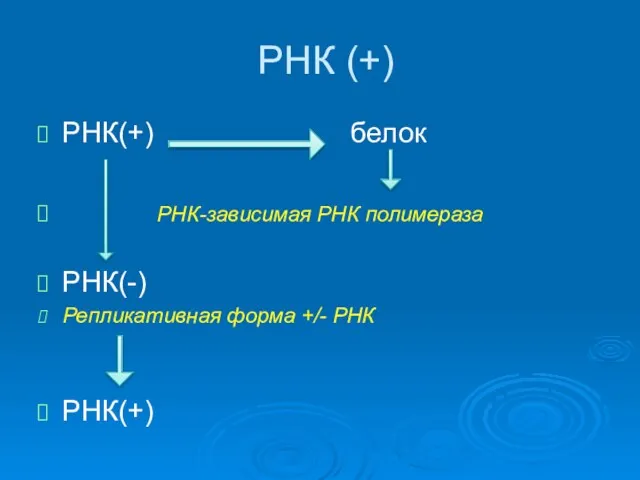
РНК (+)
РНК(+) белок
РНК-зависимая РНК полимераза
РНК(-)
Репликативная форма +/- РНК
РНК(+)
РНК (+)
РНК(+) белок
РНК-зависимая РНК полимераза
РНК(-)
Репликативная форма +/- РНК
РНК(+)
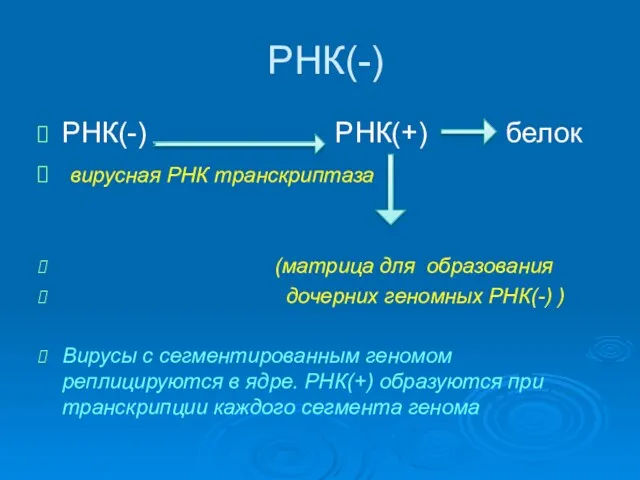
РНК(-)
РНК(-) РНК(+) белок
вирусная РНК транскриптаза
(матрица для образования
дочерних геномных РНК(-)
РНК(-)
РНК(-) РНК(+) белок
вирусная РНК транскриптаза
(матрица для образования
дочерних геномных РНК(-)

Двуцепочечные РНК (ds)
Двуцепочечная РНК (ds) не может функционировать как иРНК, поэтому
Двуцепочечные РНК (ds)
Двуцепочечная РНК (ds) не может функционировать как иРНК, поэтому

Двуцепочечные РНК (ds)
(ds) РНК иРНК белок
РНК транскриптаза вируса
Эти вирусы (Reoviridae)
Двуцепочечные РНК (ds)
(ds) РНК иРНК белок
РНК транскриптаза вируса
Эти вирусы (Reoviridae)

d+(диплоидные)РНК
1. РНК(+) ДНК
ревертаза
ДНК встраивается в геном провирус
2. ДНК иРНК
d+(диплоидные)РНК
1. РНК(+) ДНК
ревертаза
ДНК встраивается в геном провирус
2. ДНК иРНК
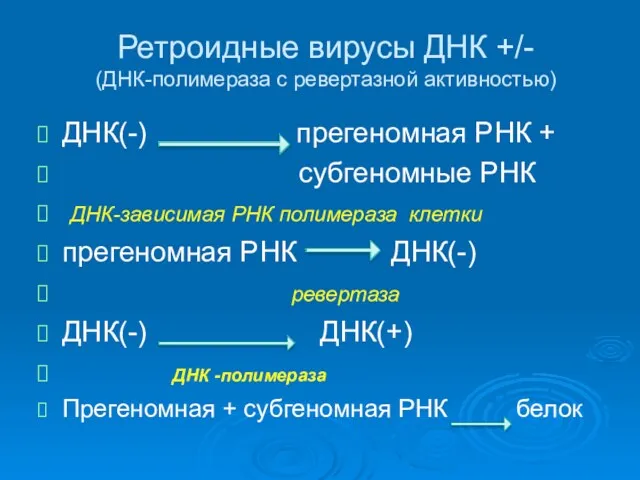
Ретроидные вирусы ДНК +/-
(ДНК-полимераза с ревертазной активностью)
ДНК(-) прегеномная РНК +
Ретроидные вирусы ДНК +/-
(ДНК-полимераза с ревертазной активностью)
ДНК(-) прегеномная РНК +

Сборка вирионов
Участок сборки вириона зависит от места репликации вирусного генома и
Сборка вирионов
Участок сборки вириона зависит от места репликации вирусного генома и

Выход вируса из клетки
Простоорганизованные вирусы выходят из клетки путем лизиса
Вирусы, покрытые
Выход вируса из клетки
Простоорганизованные вирусы выходят из клетки путем лизиса
Вирусы, покрытые
ЦПД в культуре клеток
ЦПД в культуре клеток

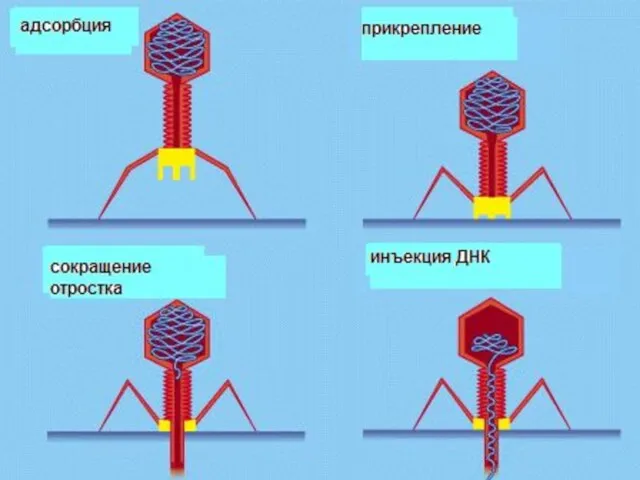
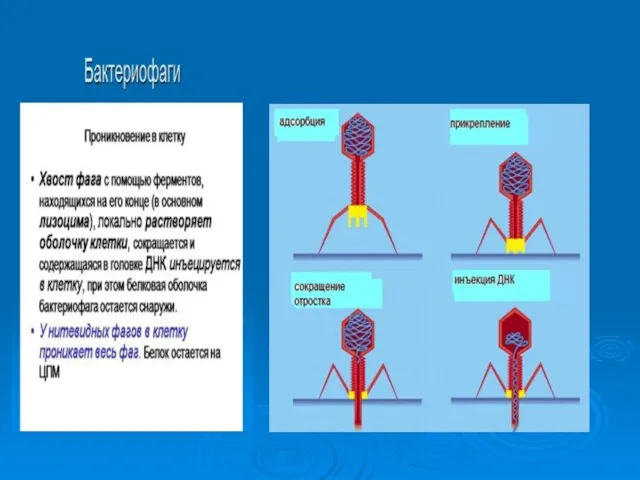
Бактериофаги
Бактериофаги
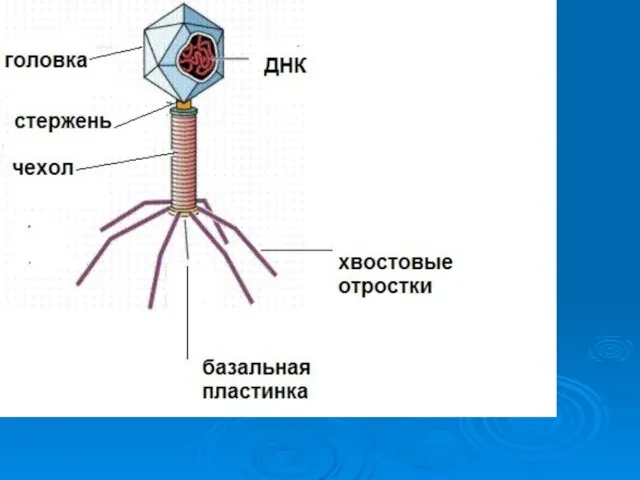

Бактериофаги
Бактериофаги
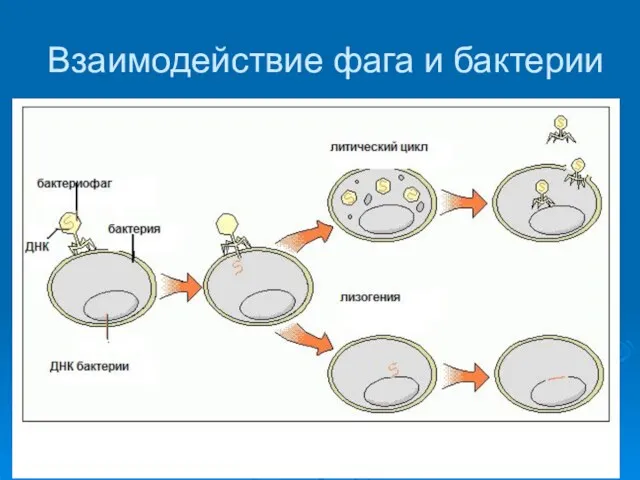
Взаимодействие фага и бактерии
Взаимодействие фага и бактерии
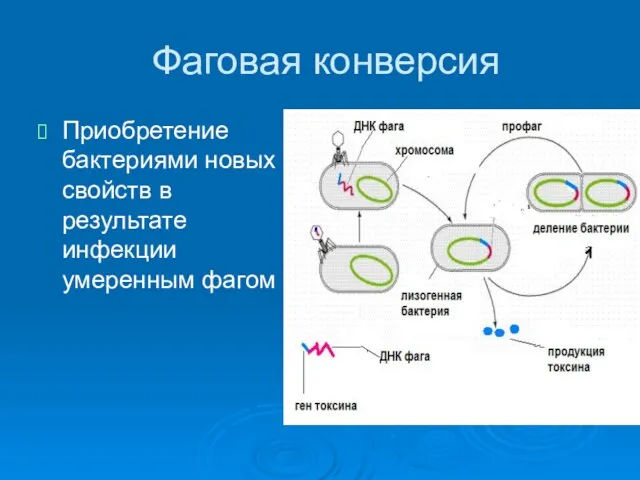
Фаговая конверсия
Приобретение бактериями новых свойств в результате инфекции умеренным фагом
Фаговая конверсия
Приобретение бактериями новых свойств в результате инфекции умеренным фагом


Применение бактериофагов
Применение бактериофагов
 Биологическое действие радиации
Биологическое действие радиации Индивидуальное развитие организмов
Индивидуальное развитие организмов Waterflood Design and Operational Best Practices
Waterflood Design and Operational Best Practices Нейроны. Функции, строение, классификация
Нейроны. Функции, строение, классификация 20. Біохімія мязів і сполучної тканини
20. Біохімія мязів і сполучної тканини Красная книга Воронежской области
Красная книга Воронежской области Прецептрон. Практична реалізація
Прецептрон. Практична реалізація Генетическая инженерия растений
Генетическая инженерия растений Салонные процедуры
Салонные процедуры Плеснивые грибы и грибы паразиты
Плеснивые грибы и грибы паразиты Эволюция. Теория Ламарка
Эволюция. Теория Ламарка Музей московского зоопарка
Музей московского зоопарка Презентация по теме Лишайники
Презентация по теме Лишайники Анатомия ЦНС. Анатомия и физиология спинного мозга
Анатомия ЦНС. Анатомия и физиология спинного мозга Сердце и кровообращение.
Сердце и кровообращение. Грибы
Грибы Интерактивный кроссворд Насекомые
Интерактивный кроссворд Насекомые презентация Пушистый доктор
презентация Пушистый доктор Дикие и домашние животные
Дикие и домашние животные Семя. Внешнее и внутреннее строение семени
Семя. Внешнее и внутреннее строение семени Генетический контроль метаболизма азота, фосфора и ретроградная регуляция
Генетический контроль метаболизма азота, фосфора и ретроградная регуляция Растительная клетка
Растительная клетка Заповедник Оренбургский
Заповедник Оренбургский Пингвины (1)
Пингвины (1) Сторінка дослідника. Твій город на підвіконні. Урок №76. Я досліджую світ
Сторінка дослідника. Твій город на підвіконні. Урок №76. Я досліджую світ Механическая обработка молока
Механическая обработка молока Презентация Загрязнение воздуха
Презентация Загрязнение воздуха Звери-млекопитающие (урок окружающего мира 1 класс)
Звери-млекопитающие (урок окружающего мира 1 класс)