- Введение генов в клетки растений - основные способы

Содержание
- 2. .ГЛАВА: ВВЕДЕНИЕ ГЕНОВ В КЛЕТКИ РАСТЕНИЙ - ОСНОВНЫЕ СПОСОБЫ ВВЕСТИ ЧУЖЕРОДНУЮ ДНК В РАСТЕНИЯ МОЖНО РАЗЛИЧНЫМИ
- 3. После проведения тем или иным способом трансформации растительной ткани ее помещают in vitro на специальную среду
- 4. Было установлено также, что обычно применяемые процедуры трансформации вовсе не безразличны и для хозяйского генома. Во-первых,
- 5. Проблема замолкания генов имеет большое практическое значение, так как у генетически трансформированных сельскохозяйственных культур трансгены должны
- 6. Стабильно введенный ген должен передаваться потомству при семенном размножении, поэтому важно определить этот параметр на практике.
- 7. Как другой биохимический маркер при анализе соматических гибридов может использоваться рибулозо - 1, 5 - бифосфат
- 8. Клеточная стенка растения повреждается вследствие выделения бактериями пектолитических ферментов, что обеспечивает плотный контакт бактерий с плазмалеммой
- 9. РИСУНОК: Генетическая колонизация растения A. tumefaciens: 1- агробактерии существуют в ризосфере; 2 - строение A. tumefaciens;
- 10. Наиболее подробно изучены опухоли — корончатые галлы, индуцируемые Agrobacterium tumefaciens. Они представляют собой истинно злокачественные опухоли,
- 11. Ткани корончатых галлов содержат более высокие уровни ауксина и цитокининов. Выявлено еще одно наследуемое изменение в
- 12. Распространение Agrobacterium и других фитопатогенных бактерий по межклетникам и ксилеме является хорошо доказанным фактом. Агробактерии могут
- 13. Невозможность заражения в природе обуславливается отсутствием соответствующих рецепторов, необходимых для взаимодействия с бактериями. Другим фактором, препятствующим
- 14. Клеточная стенка растения повреждается вследствие выделения бактериями пектолитических ферментов, что обеспечивает плотный контакт бактерий с плазмалеммой
- 15. После прохождения всех необходимых тестов на токсичность, аллергенность, мутагенность и т.д. первые трансгенные продукты появились в
- 16. Нынешний этап развития генетической инженерии растений получил название "метаболическая инженерия". При этом ставится задача не столько
- 17. Позднее ген фазеолина был передан клеткам табака: в растениях-регенерантах ген экспрессировался во всех тканях, хотя и
- 18. 1.Изучение биологии клетки, существующей вне организма, обуславливает ведущую роль клеточных культур в фундаментальных исследованиях по генетике
- 19. 1. Получение биологически активных веществ растительного происхождения: традиционных продуктов вторичного метаболизма (токсинов, гербицидов, регуляторов роста, алкалоидов,
- 21. Скачать презентацию

.ГЛАВА: ВВЕДЕНИЕ ГЕНОВ В КЛЕТКИ РАСТЕНИЙ - ОСНОВНЫЕ СПОСОБЫ
ВВЕСТИ ЧУЖЕРОДНУЮ ДНК
.ГЛАВА: ВВЕДЕНИЕ ГЕНОВ В КЛЕТКИ РАСТЕНИЙ - ОСНОВНЫЕ СПОСОБЫ ВВЕСТИ ЧУЖЕРОДНУЮ ДНК

После проведения тем или иным способом трансформации растительной ткани ее помещают
После проведения тем или иным способом трансформации растительной ткани ее помещают

Было установлено также, что обычно применяемые процедуры трансформации вовсе не безразличны
Было установлено также, что обычно применяемые процедуры трансформации вовсе не безразличны

Проблема замолкания генов имеет большое практическое значение, так как у генетически
Проблема замолкания генов имеет большое практическое значение, так как у генетически

Стабильно введенный ген должен передаваться потомству при семенном размножении, поэтому важно
Стабильно введенный ген должен передаваться потомству при семенном размножении, поэтому важно

Как другой биохимический маркер при анализе соматических гибридов может использоваться рибулозо
Как другой биохимический маркер при анализе соматических гибридов может использоваться рибулозо

Клеточная стенка растения повреждается вследствие выделения бактериями пектолитических ферментов, что обеспечивает
Клеточная стенка растения повреждается вследствие выделения бактериями пектолитических ферментов, что обеспечивает
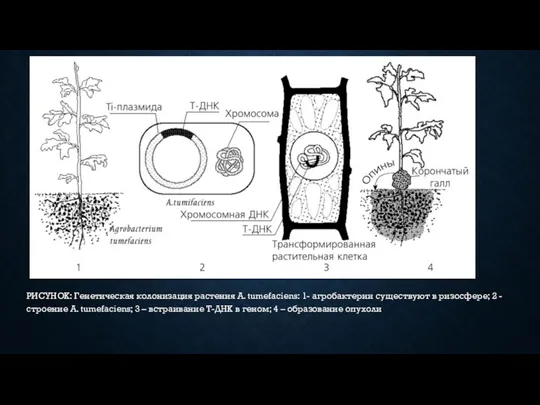
РИСУНОК: Генетическая колонизация растения A. tumefaciens: 1- агробактерии существуют в ризосфере;
РИСУНОК: Генетическая колонизация растения A. tumefaciens: 1- агробактерии существуют в ризосфере;

Наиболее подробно изучены опухоли — корончатые галлы, индуцируемые Agrobacterium tumefaciens. Они
Наиболее подробно изучены опухоли — корончатые галлы, индуцируемые Agrobacterium tumefaciens. Они

Ткани корончатых галлов содержат более высокие уровни ауксина и цитокининов. Выявлено
Ткани корончатых галлов содержат более высокие уровни ауксина и цитокининов. Выявлено

Распространение Agrobacterium и других фитопатогенных бактерий по межклетникам и ксилеме является
Распространение Agrobacterium и других фитопатогенных бактерий по межклетникам и ксилеме является

Невозможность заражения в природе обуславливается отсутствием соответствующих рецепторов, необходимых для взаимодействия
Невозможность заражения в природе обуславливается отсутствием соответствующих рецепторов, необходимых для взаимодействия

Клеточная стенка растения повреждается вследствие выделения бактериями пектолитических ферментов, что обеспечивает
Клеточная стенка растения повреждается вследствие выделения бактериями пектолитических ферментов, что обеспечивает

После прохождения всех необходимых тестов на токсичность, аллергенность, мутагенность и т.д.
После прохождения всех необходимых тестов на токсичность, аллергенность, мутагенность и т.д.

Нынешний этап развития генетической инженерии растений получил название "метаболическая инженерия". При
Нынешний этап развития генетической инженерии растений получил название "метаболическая инженерия". При

Позднее ген фазеолина был передан клеткам табака: в растениях-регенерантах ген экспрессировался
Позднее ген фазеолина был передан клеткам табака: в растениях-регенерантах ген экспрессировался

1.Изучение биологии клетки, существующей вне организма, обуславливает ведущую роль клеточных культур
1.Изучение биологии клетки, существующей вне организма, обуславливает ведущую роль клеточных культур

1. Получение биологически активных веществ растительного происхождения:
традиционных продуктов вторичного метаболизма (токсинов,
1. Получение биологически активных веществ растительного происхождения: традиционных продуктов вторичного метаболизма (токсинов,
 Членистоногие. Общая характеристика. Классы Ракообразных и Паукообразных
Членистоногие. Общая характеристика. Классы Ракообразных и Паукообразных Мінливість у людини як властивість життя і генетичне явище
Мінливість у людини як властивість життя і генетичне явище Культивирование растительных и животных клеток
Культивирование растительных и животных клеток Скелет человека
Скелет человека Мамонттар
Мамонттар Circulatory system
Circulatory system обмен веществ!!!
обмен веществ!!! Нейрон. Нервная ткань. Урок 5
Нейрон. Нервная ткань. Урок 5 Основы информационного обмена клетки с окружающей средой. Гемопоэз. (Лекция 2)
Основы информационного обмена клетки с окружающей средой. Гемопоэз. (Лекция 2) 20231109_sobaki
20231109_sobaki Protein Structure and Function
Protein Structure and Function Экология. Экосистемы
Экология. Экосистемы Пищеварение в ротовой полости и желудке
Пищеварение в ротовой полости и желудке Теоретическая, функциональная и возрастная анатомия костной системы
Теоретическая, функциональная и возрастная анатомия костной системы Біліктер мен жазықтар туралы түсініктер. Қанқадан жалпы шолу. Тұлға сүйектерінің құрылысы: омыртқалар, төс, қабырғалар
Біліктер мен жазықтар туралы түсініктер. Қанқадан жалпы шолу. Тұлға сүйектерінің құрылысы: омыртқалар, төс, қабырғалар История развития генетики. 10 класс
История развития генетики. 10 класс Хвостатая семейка. Интересные факты и истории из жизни
Хвостатая семейка. Интересные факты и истории из жизни Бурый медведь
Бурый медведь Тип Хордовые
Тип Хордовые Лисица
Лисица Биология и распространение сырдарьинского фазана
Биология и распространение сырдарьинского фазана Интегрированная система защиты чечевицы от болезней, вредителей и сорняков
Интегрированная система защиты чечевицы от болезней, вредителей и сорняков Вегетативные органы растений: корень
Вегетативные органы растений: корень Интересные факты о птицах
Интересные факты о птицах Пастер Луи
Пастер Луи Генетика людини
Генетика людини Что значит вода для человека
Что значит вода для человека Эмбриональное развитие человека
Эмбриональное развитие человека