Жизненный цикл клетки. Митотический цикл. Регуляция митотического цикла. Понятие об апоптозе презентация
- Жизненный цикл клетки. Митотический цикл. Регуляция митотического цикла. Понятие об апоптозе

Содержание
- 2. План лекции Понятие о жизненном митотическом цикле клетки Характеристика периодов митотического цикла Митоз и его биологическое
- 3. Жизненный цикл клетки ( (митотический цикЛ) Это-совокупность всех процессов, происходящих от образования клетки до ее гибели.
- 4. Митотический цикл Длительность его различна для различных организмов: для бактериальных клеток цикл может занимать 20-30 минут;
- 5. митотический цикл В типичном митотическом цикле эукариотической клетки выделяют интерфазу и Митоз
- 6. интерфаза В интерфазе выделяют 3 периода: 1.пресинтетический G1- (2n2c) 2.период синтеза ДНК, синтетический S - (2n4c)
- 7. Характеристика жизненного цикла клетки
- 8. Пресинтетический период G1 Пресинтетический период G1 - следует непосредственно за делением (за митозом) и характеризуется ростом
- 9. S-период(сл1) В S-периоде, который длится обычно 6-10 часов, происходит главный процесс - репликация (удвоение) ДНК, т.е
- 10. и продолжается синтез РНК и белков, начавшийся еще в G1-периоде: 1.происходит интенсивный синтез белков гистонов в
- 11. G2 -период (пост-синтетический) После полного удвоения хромосом наступает постсинтетический (иногда его называют премитотическим) G2 -период. В
- 12. Митоз и его биологическое значение Митоз – это непрямое деление клетки, при котором происходит точное распределение
- 13. Фазы митоза В кариокинезе различают 4 основных фазы: профазу, метафазу, анафазу и телофазу. Они непосредственно следуют
- 14. Профаза Во время профазы митоза происходят следующие процессы: 1.распад ядерной оболочки, на мелкие мембранные пузырьки (за
- 15. Профаза. (2слайд) Но главное: Хроматин начинает спирализоваться ( упаковываться) вследствие чего формируются хромосомы. При этом каждая
- 16. Метафаза.(1 сл) -Различают раннюю метафазу или прометафазу: -происходит присоединение хромосом к полюсным микротрубочкам веретена деления с
- 17. Метафаза. В животных клетках хромосомы располагаются так, что центромерные участки обращены к центру веретена, а плечи
- 18. Анафаза В анафазе выделяют два этапа А и В. Во время анафазы А движение хроматид осуществляется
- 19. Анафаза) А в анафазе В за счет удлинения полюсных и астральных микротрубочек полюса деления клетки отодвигаются
- 20. Телофаза. Она начинается с остановки хромосом и кончается реконструкцией нового интерфазного ядра. Хромосомы деконденсируются, увеличиваются в
- 21. Цитокинез За телофазой обычно следует цитокинез. Если он не происходит, то образуются многоядерные клетки (эндосперм растений,
- 22. Таким образом, в результате митотического деления происходит точное воспроизводство генетического материала и его равномерное распределение между
- 23. В то же время митоз закрепляет и отрицательные качества. Такая консервативность препятствует эволюционным изменениям. Митоз обуславливает
- 24. Клетки многоклеточного организма могут находиться в одном из трех возможных состояний: В цикле; В стадии покоя
- 25. Категории клеточных комплексов выделяют несколько различных категорий клеточных комплексов в многоклеточном организме, которые отличаются по своей
- 26. В организме существуют также клетки, которые не размножаются в обычных условиях, но вновь приобретают это свойство
- 27. 2) Растущие клеточные комплексы большинство клеток в них находятся «вне цикла» в G0 периоде - в
- 28. 3) Стабильные клеточные комплексы – нейроны и кардиомиоциты – для них характерна высокая дифференцировка и утрата
- 29. Митотический индекс Для характеристики митотической активности в тканях определяют митотический индекс – это количество делящихся клеток
- 30. Регуляция митотического цикла Процесс пролиферации(деления) клеток жестко регулируется как самой клеткой (регуляция клеточного цикла, прекращение или
- 31. Факторы регулирующие митотическую активность клеток
- 32. Регуляторные факторы, контролирующие размножение клеток Экзогенные факторы находятся в микроокружении клетки и взаимодействуют с поверхностью клетки.
- 33. Экзогенные факторы Ритмический - зависит от внешних факторов (ритма активности, света температуры) и внутренних (нейрогуморальная регуляция)
- 34. эндогенные факторы Эндокринный – саматотропин и гормоны щитовидной железы Продукты - распада тканей стимулируют митозы и
- 35. В 1989 году была присуждена Нобелевская премия за открытие двух типов генов, управляющих размножением клеток: Протоонкогены
- 36. Протоонкогены акселераторы Кодируют семейство белков называемых циклинзависимыми киназами (ЦЗК 1,2,3,4,5,6 и циклинов А, В, С, Д,
- 37. Протоонкогены супрессоры Кодируют другую группу белков-ферментов подавляющих процессы деления клеток :Р13, Р15, Р16, Р53 и убиквитин
- 38. точки рестрикции, (чикет поинт точки) Четкую смену фаз митоза определяют несколько так называемых «точек рестрикций», (чикет
- 39. Для того чтобы клетка вступила МЦ, она должна получить на мембрану митогенный сигнал, который должен дойти
- 40. Основную функцию, связанную с инициацией пролиферации, берет на себя плазматическая мембрана клетки, которая с помощью молекул-рецепторов
- 41. Этапы регуляции митотического цикла
- 42. 1.Точка рестрикции – R1 – это самый ответственный момент ( период), так как решается «судьба клетки»
- 43. Фактором стимулирующим клетку к делению является белок RUS, который выделяется соседними клетками
- 44. Полученный от белка RUS митогенный сигнал влючает первый Прото -окоген, который кодирует образование комплекса ЦЗК 2
- 45. S-периода(механизмы регуляции) Начало S-периода знаменуется появление еще одного комплекса ЦЗК 2 + ЦикЕ, который запускает работу
- 46. G2 фаза(механизмы регуляции) G2 фаза – это этап клеточного цикла, который начинается после завершения синтеза ДНК,
- 47. Повреждения ДНК или наличие нереплицированных участков предотвращает переход в М фазу. Запускается митоз распадом комплекса В-CDK1
- 49. Скачать презентацию

План лекции
Понятие о жизненном митотическом цикле клетки
Характеристика периодов митотического
План лекции
Понятие о жизненном митотическом цикле клетки
Характеристика периодов митотического
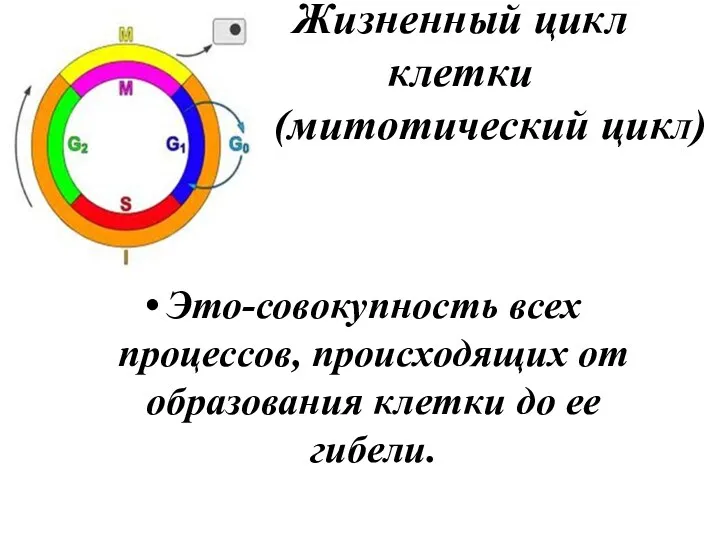
Жизненный цикл
клетки
( (митотический цикЛ)
Это-совокупность всех процессов, происходящих от образования
Жизненный цикл
клетки
( (митотический цикЛ)
Это-совокупность всех процессов, происходящих от образования

Митотический цикл
Длительность его различна для различных организмов: для бактериальных клеток
Митотический цикл
Длительность его различна для различных организмов: для бактериальных клеток
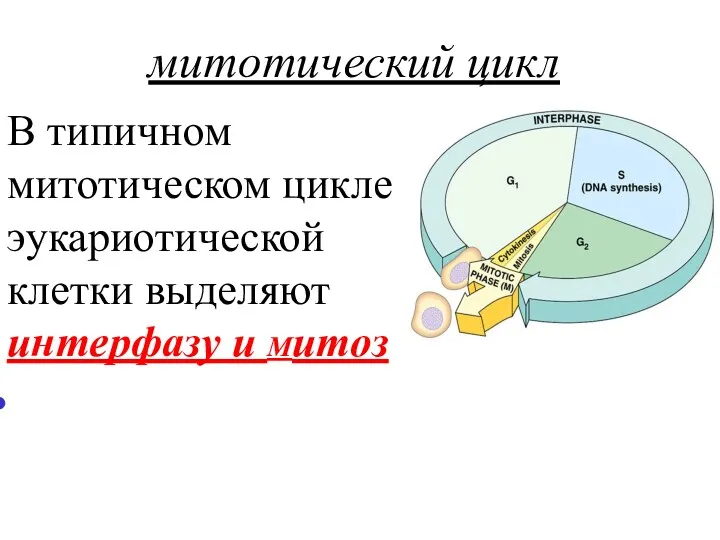
митотический цикл
В типичном митотическом цикле эукариотической клетки выделяют интерфазу и
митотический цикл
В типичном митотическом цикле эукариотической клетки выделяют интерфазу и
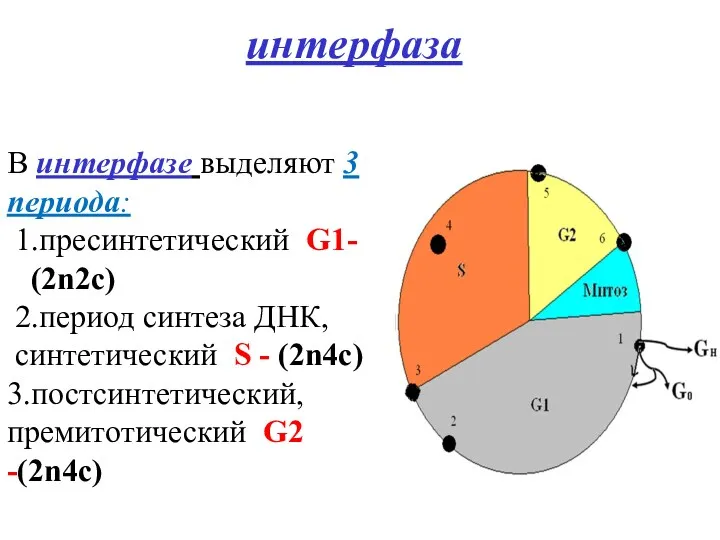
интерфаза
В интерфазе выделяют 3 периода:
1.пресинтетический G1-
(2n2c)
2.период синтеза ДНК,
интерфаза
В интерфазе выделяют 3 периода:
1.пресинтетический G1-
(2n2c)
2.период синтеза ДНК,
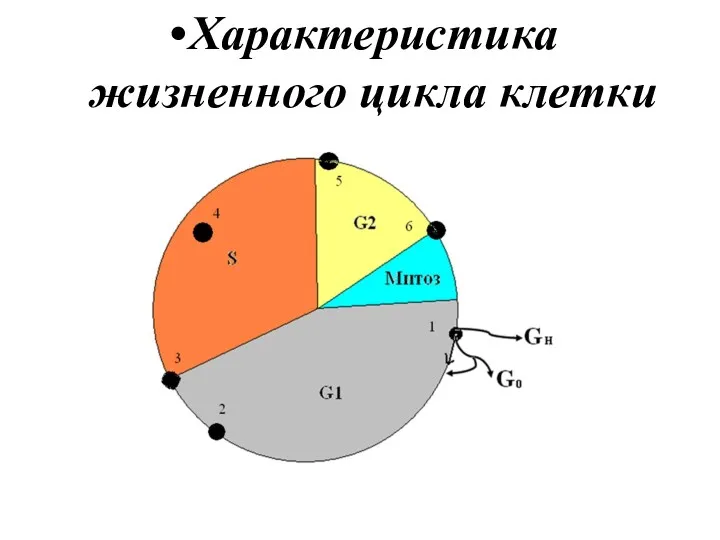
Характеристика жизненного цикла клетки
Характеристика жизненного цикла клетки

Пресинтетический период G1
Пресинтетический период G1 -
следует непосредственно за делением
Пресинтетический период G1
Пресинтетический период G1 -
следует непосредственно за делением

S-период(сл1)
В S-периоде, который длится обычно 6-10 часов, происходит главный процесс
S-период(сл1)
В S-периоде, который длится обычно 6-10 часов, происходит главный процесс

и продолжается синтез РНК и белков, начавшийся еще в G1-периоде:
1.происходит
и продолжается синтез РНК и белков, начавшийся еще в G1-периоде:
1.происходит

G2 -период (пост-синтетический)
После полного удвоения хромосом наступает постсинтетический (иногда его называют
G2 -период (пост-синтетический)
После полного удвоения хромосом наступает постсинтетический (иногда его называют
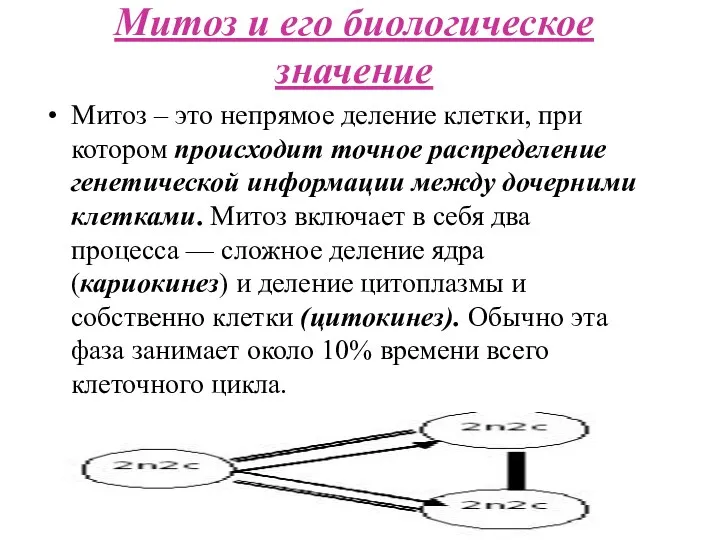
Митоз и его биологическое значение
Митоз – это непрямое деление клетки, при
Митоз и его биологическое значение
Митоз – это непрямое деление клетки, при
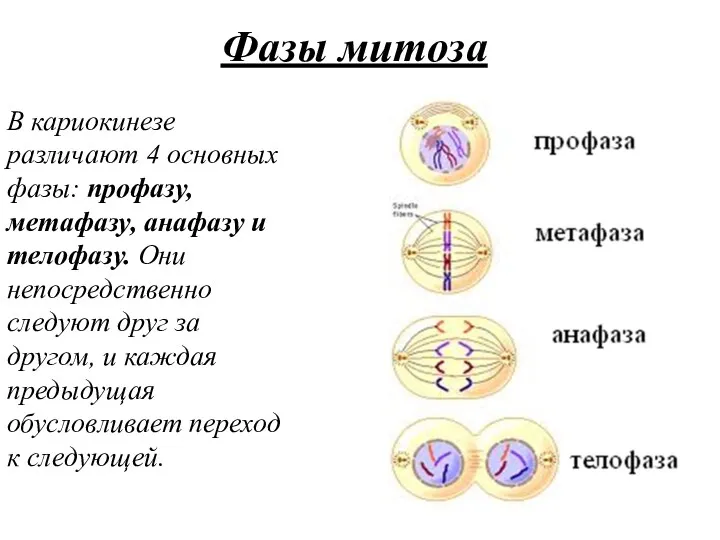
Фазы митоза
В кариокинезе различают 4 основных фазы: профазу, метафазу, анафазу и
Фазы митоза
В кариокинезе различают 4 основных фазы: профазу, метафазу, анафазу и

Профаза
Во время профазы митоза происходят следующие процессы:
1.распад ядерной оболочки, на
Профаза
Во время профазы митоза происходят следующие процессы:
1.распад ядерной оболочки, на

Профаза. (2слайд)
Но главное:
Хроматин начинает спирализоваться
( упаковываться) вследствие чего формируются
Профаза. (2слайд)
Но главное:
Хроматин начинает спирализоваться
( упаковываться) вследствие чего формируются

Метафаза.(1 сл)
-Различают раннюю метафазу или прометафазу:
-происходит присоединение хромосом
Метафаза.(1 сл)
-Различают раннюю метафазу или прометафазу:
-происходит присоединение хромосом

Метафаза.
В животных клетках хромосомы располагаются так, что центромерные участки обращены
Метафаза.
В животных клетках хромосомы располагаются так, что центромерные участки обращены
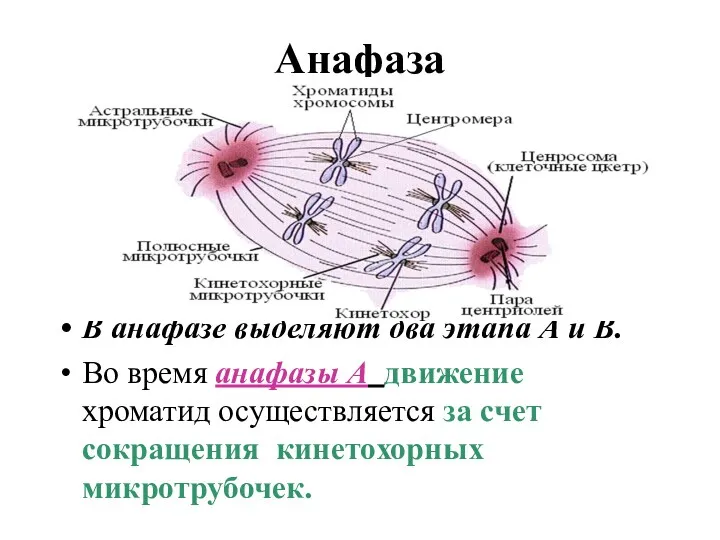
Анафаза
В анафазе выделяют два этапа А и В.
Во время анафазы А
Анафаза
В анафазе выделяют два этапа А и В.
Во время анафазы А
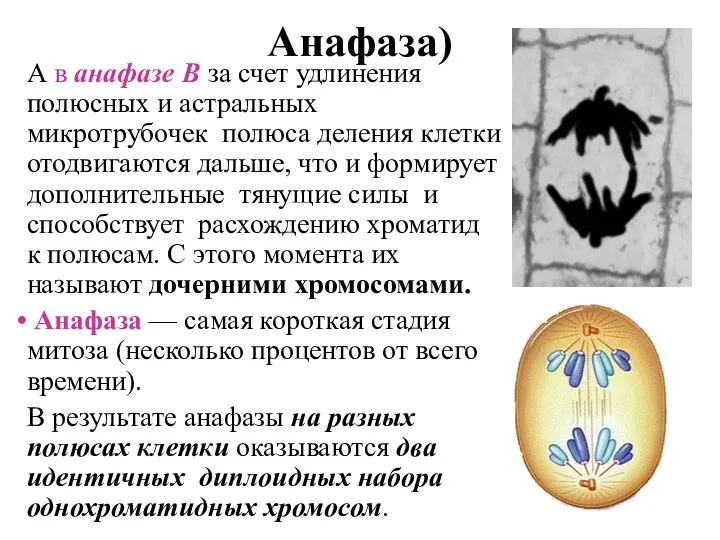
Анафаза)
А в анафазе В за счет удлинения полюсных и астральных
Анафаза)
А в анафазе В за счет удлинения полюсных и астральных
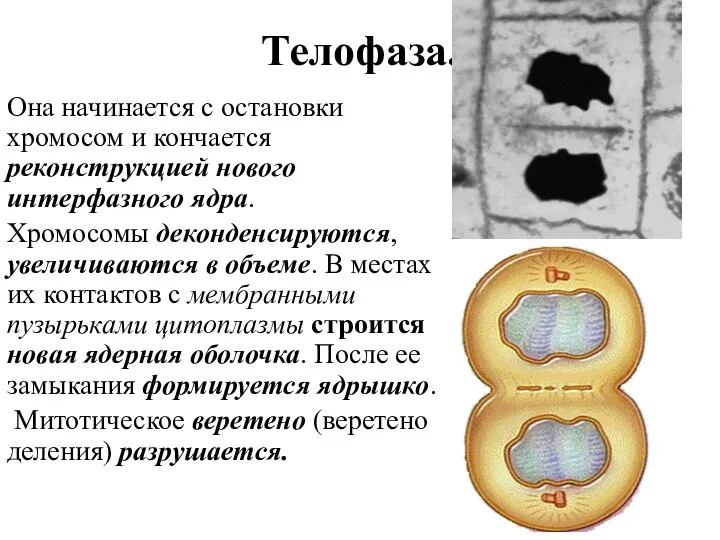
Телофаза.
Она начинается с остановки хромосом и кончается реконструкцией нового интерфазного
Телофаза.
Она начинается с остановки хромосом и кончается реконструкцией нового интерфазного

Цитокинез
За телофазой обычно следует цитокинез. Если он не происходит, то
Цитокинез
За телофазой обычно следует цитокинез. Если он не происходит, то

Таким образом, в результате митотического деления происходит точное воспроизводство генетического материала
Таким образом, в результате митотического деления происходит точное воспроизводство генетического материала

В то же время митоз закрепляет и отрицательные качества. Такая консервативность
В то же время митоз закрепляет и отрицательные качества. Такая консервативность

Клетки многоклеточного организма могут находиться в одном из трех возможных состояний:
В
Клетки многоклеточного организма могут находиться в одном из трех возможных состояний:
В

Категории клеточных комплексов
выделяют несколько различных категорий клеточных комплексов в многоклеточном организме,
Категории клеточных комплексов
выделяют несколько различных категорий клеточных комплексов в многоклеточном организме,

В организме существуют также клетки, которые не размножаются в обычных
В организме существуют также клетки, которые не размножаются в обычных

2) Растущие клеточные комплексы большинство клеток в них находятся «вне цикла»
2) Растущие клеточные комплексы большинство клеток в них находятся «вне цикла»

3) Стабильные клеточные комплексы – нейроны и кардиомиоциты – для них
3) Стабильные клеточные комплексы – нейроны и кардиомиоциты – для них

Митотический индекс
Для характеристики митотической активности в тканях определяют митотический
Митотический индекс
Для характеристики митотической активности в тканях определяют митотический

Регуляция митотического цикла
Процесс пролиферации(деления) клеток жестко регулируется как самой клеткой
Регуляция митотического цикла
Процесс пролиферации(деления) клеток жестко регулируется как самой клеткой

Факторы регулирующие митотическую активность клеток
Факторы регулирующие митотическую активность клеток

Регуляторные факторы, контролирующие размножение клеток
Экзогенные факторы находятся в микроокружении клетки
Регуляторные факторы, контролирующие размножение клеток
Экзогенные факторы находятся в микроокружении клетки

Экзогенные факторы
Ритмический - зависит от внешних факторов (ритма активности, света температуры)
Экзогенные факторы
Ритмический - зависит от внешних факторов (ритма активности, света температуры)

эндогенные факторы
Эндокринный – саматотропин и гормоны щитовидной железы
Продукты - распада
эндогенные факторы
Эндокринный – саматотропин и гормоны щитовидной железы
Продукты - распада

В 1989 году была присуждена Нобелевская премия за открытие двух типов
В 1989 году была присуждена Нобелевская премия за открытие двух типов

Протоонкогены акселераторы
Кодируют семейство белков называемых циклинзависимыми киназами (ЦЗК 1,2,3,4,5,6 и
Протоонкогены акселераторы
Кодируют семейство белков называемых циклинзависимыми киназами (ЦЗК 1,2,3,4,5,6 и

Протоонкогены супрессоры
Кодируют другую группу белков-ферментов подавляющих процессы деления клеток :Р13,
Протоонкогены супрессоры
Кодируют другую группу белков-ферментов подавляющих процессы деления клеток :Р13,

точки рестрикции, (чикет поинт точки)
Четкую смену фаз митоза определяют несколько
точки рестрикции, (чикет поинт точки)
Четкую смену фаз митоза определяют несколько

Для того чтобы клетка вступила МЦ, она должна получить на мембрану
Для того чтобы клетка вступила МЦ, она должна получить на мембрану

Основную функцию, связанную с инициацией пролиферации, берет на себя плазматическая мембрана
Основную функцию, связанную с инициацией пролиферации, берет на себя плазматическая мембрана

Этапы регуляции митотического цикла
Этапы регуляции митотического цикла

1.Точка рестрикции – R1 – это самый ответственный момент ( период),
1.Точка рестрикции – R1 – это самый ответственный момент ( период),

Фактором стимулирующим клетку к делению является белок RUS, который выделяется соседними
Фактором стимулирующим клетку к делению является белок RUS, который выделяется соседними

Полученный от белка RUS митогенный сигнал
влючает первый Прото -окоген, который
кодирует
Полученный от белка RUS митогенный сигнал
влючает первый Прото -окоген, который
кодирует

S-периода(механизмы регуляции)
Начало S-периода знаменуется появление еще одного комплекса ЦЗК 2 +
S-периода(механизмы регуляции)
Начало S-периода знаменуется появление еще одного комплекса ЦЗК 2 +

G2 фаза(механизмы регуляции)
G2 фаза – это этап клеточного цикла, который начинается
G2 фаза(механизмы регуляции)
G2 фаза – это этап клеточного цикла, который начинается

Повреждения ДНК или наличие нереплицированных участков предотвращает переход в М
Повреждения ДНК или наличие нереплицированных участков предотвращает переход в М
 Теоретическая, функциональная и возрастная анатомия костной системы
Теоретическая, функциональная и возрастная анатомия костной системы Общие свойства живого
Общие свойства живого Водоросли. 6 класс
Водоросли. 6 класс Конечный мозг (thelencephalon, cerebrum)
Конечный мозг (thelencephalon, cerebrum) Тайская кошка
Тайская кошка Красная книга
Красная книга Введение в биомеханику. Общая, дифференциальная и частная биомеханика
Введение в биомеханику. Общая, дифференциальная и частная биомеханика Ноосфера
Ноосфера Сон и его значение
Сон и его значение Микрофлора организма человека. Дисбактериоз. Экология микроорганизмов
Микрофлора организма человека. Дисбактериоз. Экология микроорганизмов Основы селекции организмов
Основы селекции организмов Зимующие птицы
Зимующие птицы Биологические ресурсы
Биологические ресурсы Презентация по биологии на тему Абиотические факторы среды. Влажность для 11 класса
Презентация по биологии на тему Абиотические факторы среды. Влажность для 11 класса Авторская дидактическая игра Угадай, кто спрятался?
Авторская дидактическая игра Угадай, кто спрятался? Биологические ритмы
Биологические ритмы Закон гомологических рядов
Закон гомологических рядов Семейство кошачьих
Семейство кошачьих Размножение - свойство живых организмов
Размножение - свойство живых организмов Введение в биоинформатику. Биологические базы данных. Лекция 1
Введение в биоинформатику. Биологические базы данных. Лекция 1 Организация генома. Прокариот и эукариот
Организация генома. Прокариот и эукариот Многолетние растения: флоксы, пионы, розы
Многолетние растения: флоксы, пионы, розы Готовимся к ЕГЭ по биологии
Готовимся к ЕГЭ по биологии Презентация к выступлению на научной конференции Совершенствование естественнонаучного школьного образования
Презентация к выступлению на научной конференции Совершенствование естественнонаучного школьного образования Пептиды. Белки. (Лекция 16)
Пептиды. Белки. (Лекция 16) Классификация царства грибы. Низшие и высшие грибы
Классификация царства грибы. Низшие и высшие грибы Редкие животные нашего округа
Редкие животные нашего округа Интегративная деятельность организма. Рефлексы
Интегративная деятельность организма. Рефлексы