- Зрение. Оптические и жидкостные среды глаза

Содержание
- 2. Оптические и жидкостные среды глаза Прежде чем световая волна достигнет рецепторных клеток (палочек и колбочек), расположенных
- 3. Рефракция Преломление происходит в связи с тем, что луч света с различной скоростью про-ходит через данные
- 4. Слезная жидкость Слезная жидкость улучшает оптические свойства роговицы, защищая ее от пыли и пересыхания. Стекая в
- 5. Водянистая влага глаза Водянистая влага глаза секрети-руется клетками цилиарного тела в заднюю камеру глаза. Через зрачок
- 6. Фильтрация светового потока Оптические среды глаза не только фокусируют лучи на рецепторных клетках сетчатки, но и
- 7. Аккомодация Для ясного видения предмета необходимо, чтобы лучи от отдельных точек его были сфокусированы на сетчатке.
- 8. Механизм аккомодации: Аккомодация обеспечивается хрусталиком, кривизна которого может меняться (рис.) в диапазоне от 15 D до
- 9. Регуляция аккомодации Цилиарная мышца иннервируется парасимпатическими волокнами глазодвигательного нерва, при возбуждении которых глаз начинает ясно видеть
- 10. Оптические несовершенства глаза Сферическая аберрация: центр больше преломляет чем периферия (поэтому вечером предметы нечеткие). Хроматическая аберрация:
- 11. Несовершенства глазного яблока и патология аккомодации 1 - норма, 2 – миопия (исправляется двояковогнутой линзой), 3
- 12. Зрачок Зрачок регулирует количество света, поступающего к сетчатке (т.е. участвует в адаптации зрения). Зрачковый рефлекс регулируется
- 13. Сетчатка Схема расположения клеточных элементов сетчатки: слева - на периферии, справа - в центре.
- 14. Схема палочки и колбочки
- 15. Зрительные пигменты Светочувствительный сегмент фоторецепторов содержит зрительные пигменты, а на противоположном конце их имеется синапс с
- 16. Плотность размещения палочек и колбочек в различных отделах сетчатки. Кривая остроты зрения (справа) полностью совпадает с
- 17. Расположение родопсина на мембране палочки
- 18. Превращение молекулы родопсина при поступлении кванта света. Трансретиналь восстанавливается до вит. А. Поэтому для восприятия следующего
- 19. Витамин А и восстановление родопсина При нехватке в пище витамина А родопсин восстанавливается плохо – ночная
- 20. Медиатор и действие света В темноте Na+-каналы отрыты (-25 мВ) – деполяризация (РП) выделение медиатора. На
- 21. Включение вторых посредников При изменении зрительного пигмента запускаются вторые посредники (цГМФ), которые и регулируют проницаемость мембраны
- 22. Различие адаптации палочек и колбочек Рецепторный потенциал в палочках развивается медленнее, чем в колбочках. Обусловлено это
- 23. Но основное значение в адаптации принадлежит изменению количества зрительных пигментов: на свету они разрушаются, а в
- 24. Нейроны сетчатки – первый нервный центр В нейронах сетчатки при передаче сигналов широко происходят процессы схождения
- 25. Медиаторы нейронов Медиаторы фоторецепторов и большинства нейронов сетчатки (глютамат) оказывают тормозное влияние на постсинаптическую мембрану, вызывая
- 26. Биполярные клетки В сетчатке имеется два типа биполярных клеток – депо-ляризующиеся и гиперполяризующиеся. Эти клетки обладают
- 27. Ганглиозные поля сетчатки Ганглиозная клетка связана с многими рецепторными клетками сетчатки – это ее рецептивное поле
- 28. Ганглиозные поля палочек Рецептивные поля ганглиозных нейронов организованы с антагонистической характеристикой центра и периферии. Если свет
- 29. Ганглиозная клетка с on-центром ПД в ганглиозной клетке при освещении периферии не появляется. Это РП с
- 30. Схема исследования остроты зрения Ганглиозные поля обеспечивают возможность видеть отдельные детали предмета – ОСТРОТУ ЗРЕНИЯ, так
- 31. Ганглиозные поля колбочкового аппарата В основе формирования пар колбочек лежат 4 основных цвета: красный-зеленый, желтый-синий. Имеется
- 32. Аддитивное смешение трех основных цветов Откуда же взялся желтый цвет? Четвертый (желтый) цвет появляется при смешение
- 33. Состояние спектральной чувствительности трех типов колбочек в зависимости от длины волны Максимумы чувствительности при: Родопсин -
- 34. Зрительный тракт На уровне нейронов подкорковых ядер также можно обнаружить наличие рецептивных полей, обеспечивающих их связь
- 35. Верхние бугры четверохолмия Импульсы, поступающие в верхние бугорки чрезвычайно важны для определения движущегося объекта и регуляции
- 36. Запись движения глаз при рассматривании неподвижного предмета в течение 2-х минут Так как зрительный пигмент каждые
- 37. Мышцы, осуществляющие движения глаз
- 38. Схема путей, управляющих движениями глаз Для сознательных движений глаза центры находятся в лобной доле коры .
- 39. Латеральное коленчатое тело В латеральном коленчатом теле три слоя нейронов связаны с сетчаткой этой же стороны,
- 40. Расположение нейронов в коре от двух глаз К затылочным полям коры импульсы поступают от обеих глаз
- 41. Схема опознания образа с помощью различного типа нейронов (1-3) 1 – образ, 2 – врожденные характеристики
- 43. Скачать презентацию

Оптические и жидкостные среды глаза
Прежде чем световая волна достигнет рецепторных
Оптические и жидкостные среды глаза
Прежде чем световая волна достигнет рецепторных
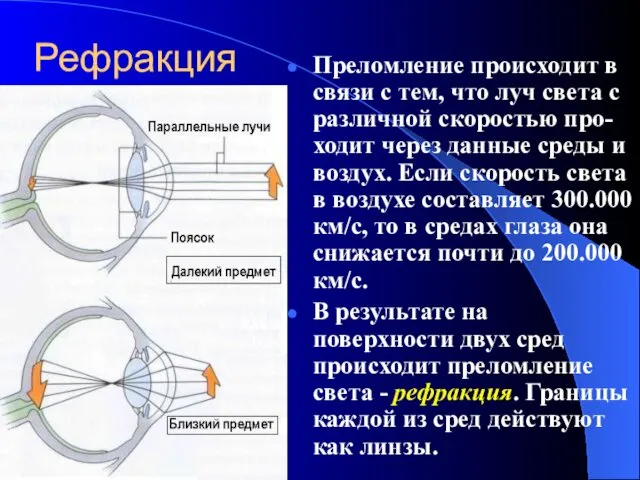
Рефракция
Преломление происходит в связи с тем, что луч света с различной
Рефракция
Преломление происходит в связи с тем, что луч света с различной
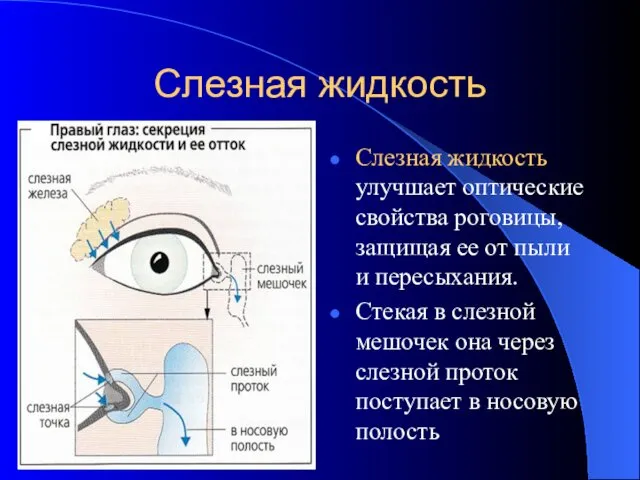
Слезная жидкость
Слезная жидкость улучшает оптические свойства роговицы, защищая ее от пыли
Слезная жидкость
Слезная жидкость улучшает оптические свойства роговицы, защищая ее от пыли

Водянистая влага глаза
Водянистая влага глаза секрети-руется клетками цилиарного тела в заднюю
Водянистая влага глаза
Водянистая влага глаза секрети-руется клетками цилиарного тела в заднюю

Фильтрация светового потока
Оптические среды глаза не только фокусируют лучи на рецепторных
Фильтрация светового потока
Оптические среды глаза не только фокусируют лучи на рецепторных

Аккомодация
Для ясного видения предмета необходимо, чтобы лучи от отдельных точек его
Аккомодация
Для ясного видения предмета необходимо, чтобы лучи от отдельных точек его

Механизм аккомодации:
Аккомодация обеспечивается хрусталиком, кривизна которого может меняться (рис.) в
Механизм аккомодации:
Аккомодация обеспечивается хрусталиком, кривизна которого может меняться (рис.) в

Регуляция аккомодации
Цилиарная мышца иннервируется парасимпатическими волокнами глазодвигательного нерва, при возбуждении которых
Регуляция аккомодации
Цилиарная мышца иннервируется парасимпатическими волокнами глазодвигательного нерва, при возбуждении которых

Оптические несовершенства глаза
Сферическая аберрация: центр больше преломляет чем периферия (поэтому вечером
Оптические несовершенства глаза
Сферическая аберрация: центр больше преломляет чем периферия (поэтому вечером
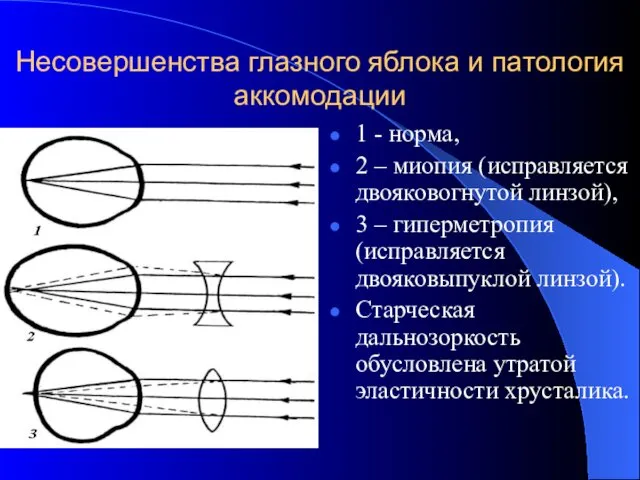
Несовершенства глазного яблока и патология аккомодации
1 - норма,
2 – миопия
Несовершенства глазного яблока и патология аккомодации
1 - норма,
2 – миопия
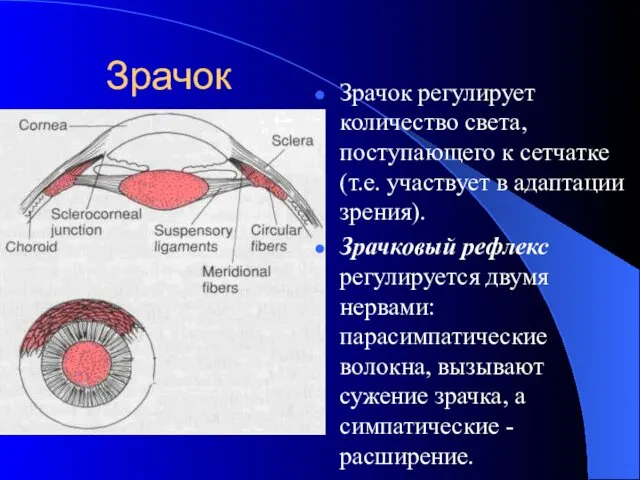
Зрачок
Зрачок регулирует количество света, поступающего к сетчатке (т.е. участвует в адаптации
Зрачок
Зрачок регулирует количество света, поступающего к сетчатке (т.е. участвует в адаптации
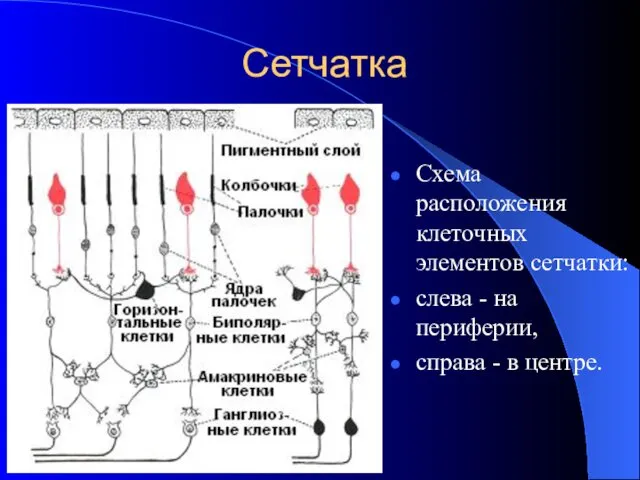
Сетчатка
Схема расположения клеточных элементов сетчатки:
слева - на периферии,
справа -
Сетчатка
Схема расположения клеточных элементов сетчатки:
слева - на периферии,
справа -
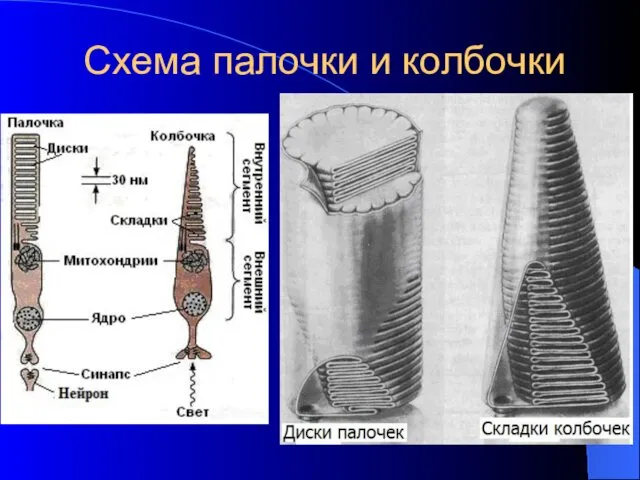
Схема палочки и колбочки
Схема палочки и колбочки

Зрительные пигменты
Светочувствительный сегмент фоторецепторов содержит зрительные пигменты, а на противоположном конце
Зрительные пигменты
Светочувствительный сегмент фоторецепторов содержит зрительные пигменты, а на противоположном конце
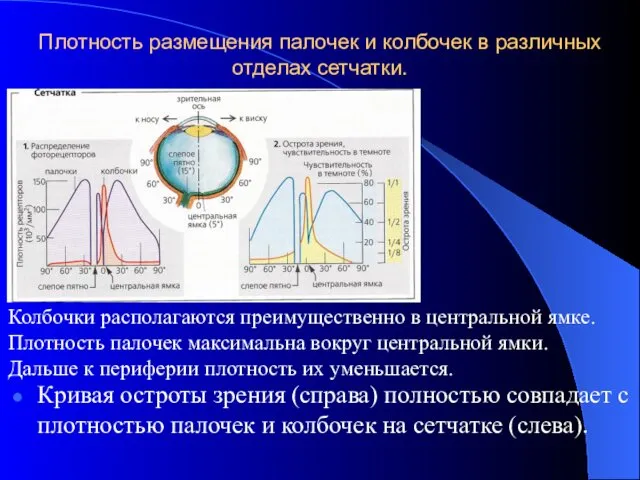
Плотность размещения палочек и колбочек в различных отделах сетчатки.
Кривая остроты зрения
Плотность размещения палочек и колбочек в различных отделах сетчатки.
Кривая остроты зрения
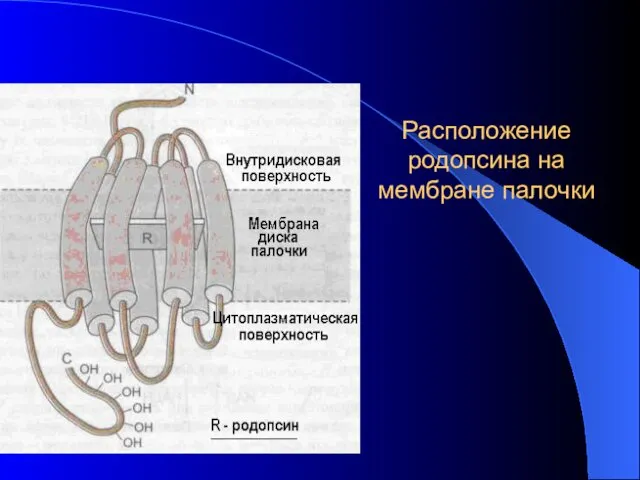
Расположение родопсина на мембране палочки
Расположение родопсина на мембране палочки
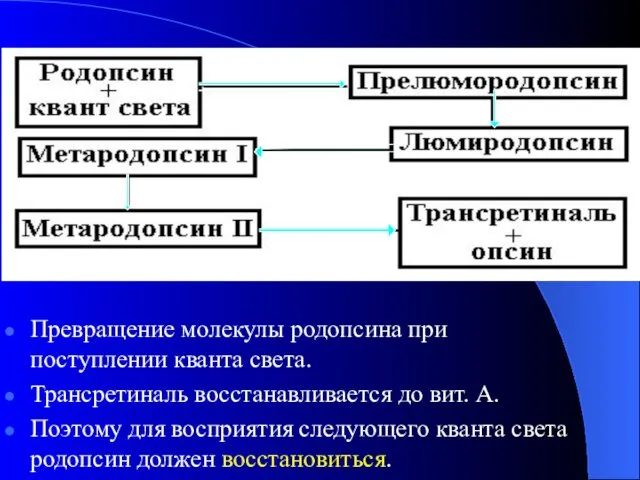
Превращение молекулы родопсина при поступлении кванта света.
Трансретиналь восстанавливается до вит. А.
Поэтому
Превращение молекулы родопсина при поступлении кванта света.
Трансретиналь восстанавливается до вит. А.
Поэтому

Витамин А и восстановление родопсина
При нехватке в пище витамина А родопсин
Витамин А и восстановление родопсина
При нехватке в пище витамина А родопсин
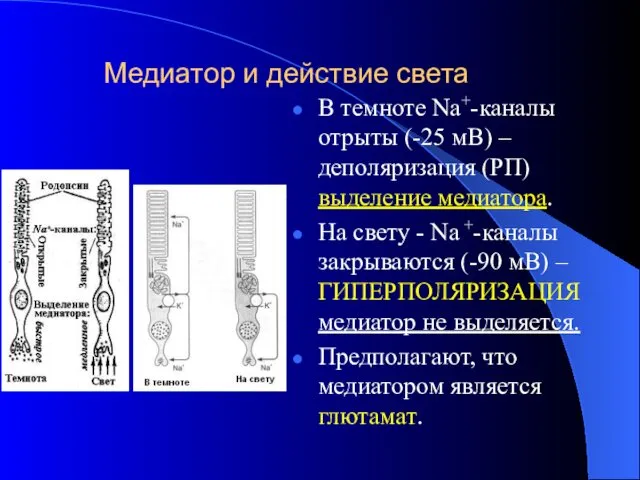
Медиатор и действие света
В темноте Na+-каналы отрыты (-25 мВ) –
Медиатор и действие света
В темноте Na+-каналы отрыты (-25 мВ) –
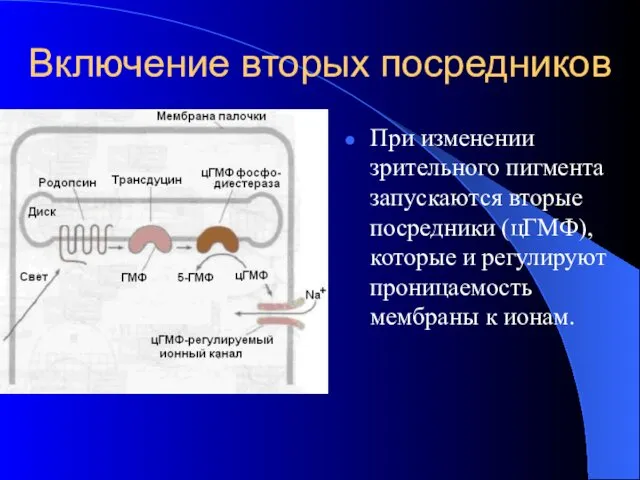
Включение вторых посредников
При изменении зрительного пигмента запускаются вторые посредники (цГМФ), которые
Включение вторых посредников
При изменении зрительного пигмента запускаются вторые посредники (цГМФ), которые

Различие адаптации палочек и колбочек
Рецепторный потенциал в палочках развивается медленнее, чем
Различие адаптации палочек и колбочек
Рецепторный потенциал в палочках развивается медленнее, чем

Но основное значение в адаптации принадлежит изменению количества зрительных пигментов:
на свету
Но основное значение в адаптации принадлежит изменению количества зрительных пигментов:
на свету
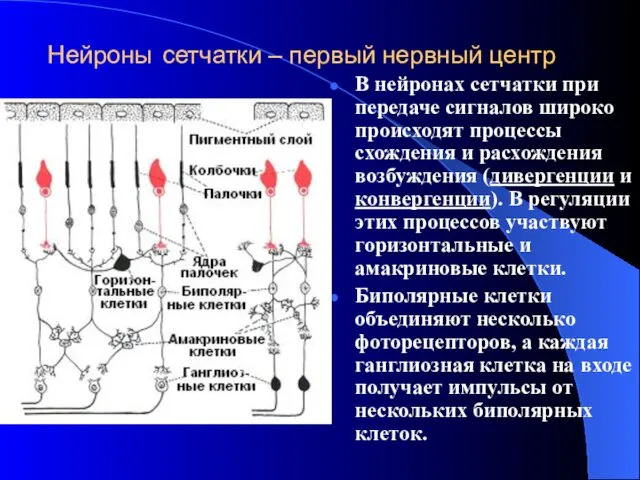
Нейроны сетчатки – первый нервный центр
В нейронах сетчатки при передаче сигналов
Нейроны сетчатки – первый нервный центр
В нейронах сетчатки при передаче сигналов

Медиаторы нейронов
Медиаторы фоторецепторов и большинства нейронов сетчатки (глютамат) оказывают тормозное влияние
Медиаторы нейронов
Медиаторы фоторецепторов и большинства нейронов сетчатки (глютамат) оказывают тормозное влияние

Биполярные клетки
В сетчатке имеется два типа биполярных клеток – депо-ляризующиеся и
Биполярные клетки
В сетчатке имеется два типа биполярных клеток – депо-ляризующиеся и
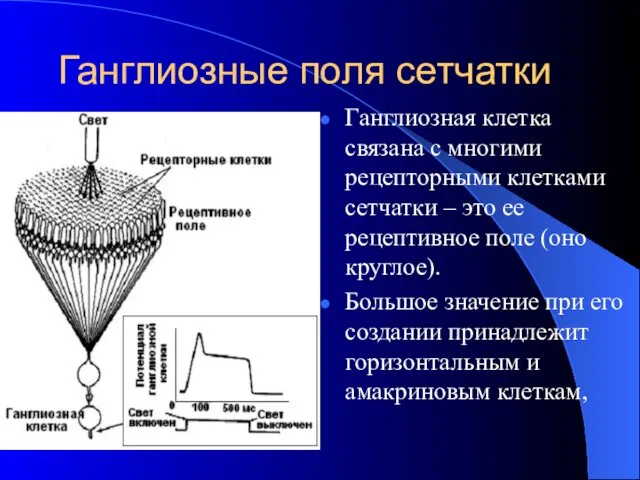
Ганглиозные поля сетчатки
Ганглиозная клетка связана с многими рецепторными клетками сетчатки –
Ганглиозные поля сетчатки
Ганглиозная клетка связана с многими рецепторными клетками сетчатки –

Ганглиозные поля палочек
Рецептивные поля ганглиозных нейронов организованы с антагонистической характеристикой центра
Ганглиозные поля палочек
Рецептивные поля ганглиозных нейронов организованы с антагонистической характеристикой центра
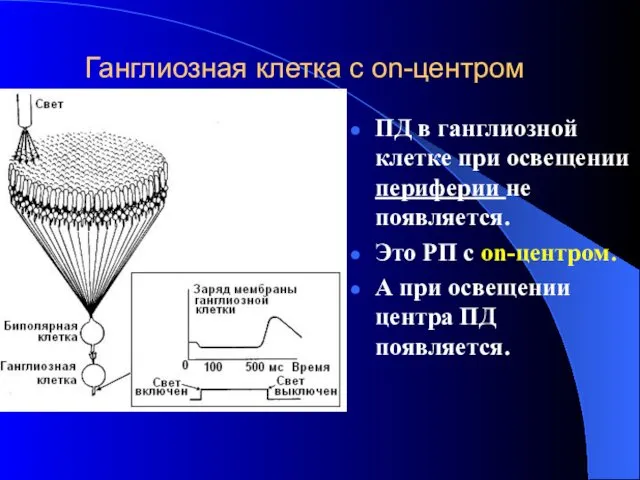
Ганглиозная клетка с on-центром
ПД в ганглиозной клетке при освещении периферии не
Ганглиозная клетка с on-центром
ПД в ганглиозной клетке при освещении периферии не

Схема исследования остроты зрения
Ганглиозные поля обеспечивают возможность видеть отдельные детали
Схема исследования остроты зрения
Ганглиозные поля обеспечивают возможность видеть отдельные детали

Ганглиозные поля колбочкового аппарата
В основе формирования пар колбочек лежат 4 основных
Ганглиозные поля колбочкового аппарата
В основе формирования пар колбочек лежат 4 основных

Аддитивное смешение трех основных цветов
Откуда же взялся желтый цвет?
Четвертый (желтый) цвет
Аддитивное смешение трех основных цветов
Откуда же взялся желтый цвет?
Четвертый (желтый) цвет
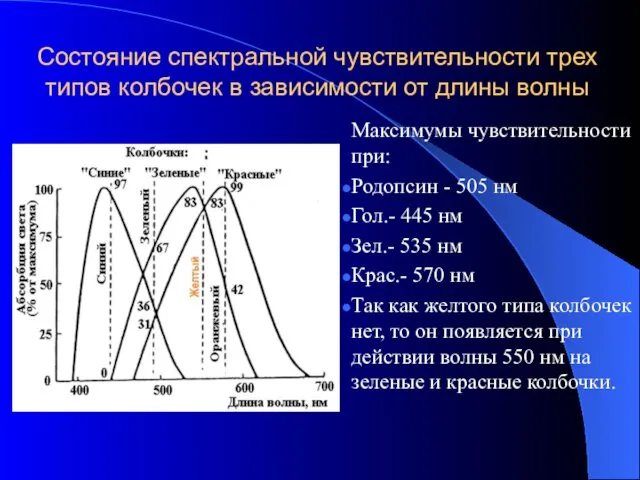
Состояние спектральной чувствительности трех типов колбочек в зависимости от длины волны
Максимумы
Состояние спектральной чувствительности трех типов колбочек в зависимости от длины волны
Максимумы
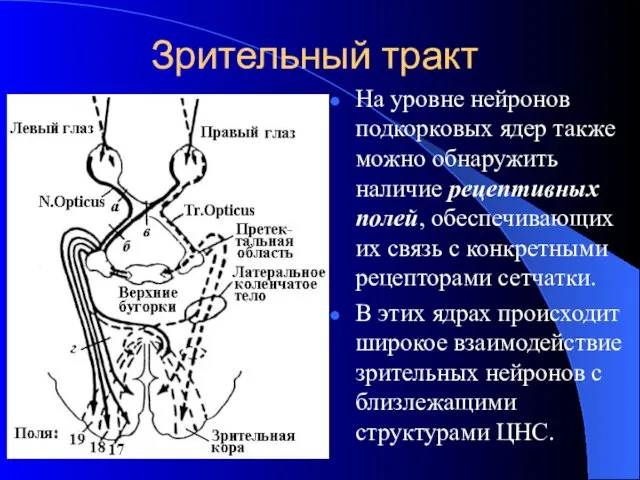
Зрительный тракт
На уровне нейронов подкорковых ядер также можно обнаружить наличие рецептивных
Зрительный тракт
На уровне нейронов подкорковых ядер также можно обнаружить наличие рецептивных

Верхние бугры четверохолмия
Импульсы, поступающие в верхние бугорки чрезвычайно важны для определения
Верхние бугры четверохолмия
Импульсы, поступающие в верхние бугорки чрезвычайно важны для определения
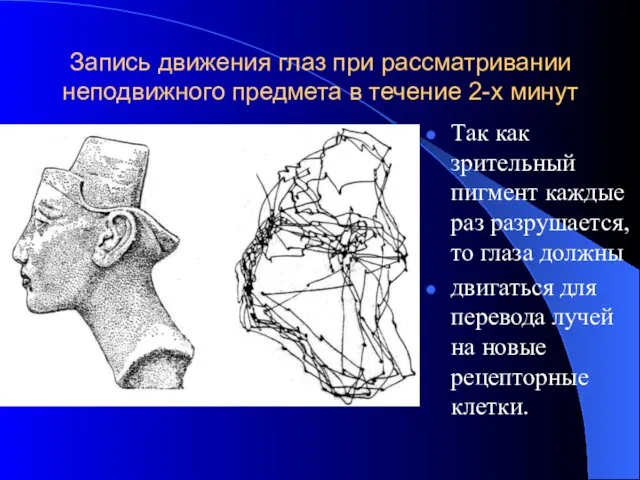
Запись движения глаз при рассматривании неподвижного предмета в течение 2-х минут
Запись движения глаз при рассматривании неподвижного предмета в течение 2-х минут
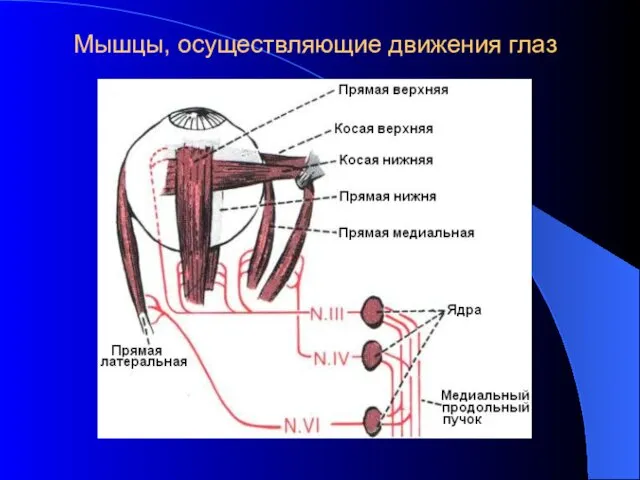
Мышцы, осуществляющие движения глаз
Мышцы, осуществляющие движения глаз
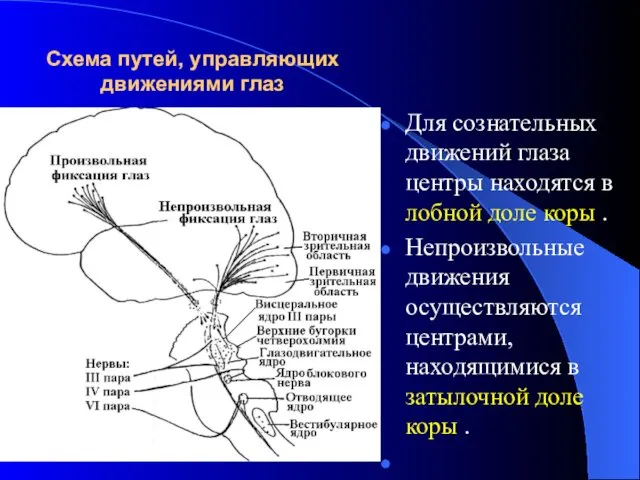
Схема путей, управляющих движениями глаз
Для сознательных движений глаза центры находятся
Схема путей, управляющих движениями глаз
Для сознательных движений глаза центры находятся

Латеральное коленчатое тело
В латеральном коленчатом теле три слоя нейронов связаны с
Латеральное коленчатое тело
В латеральном коленчатом теле три слоя нейронов связаны с
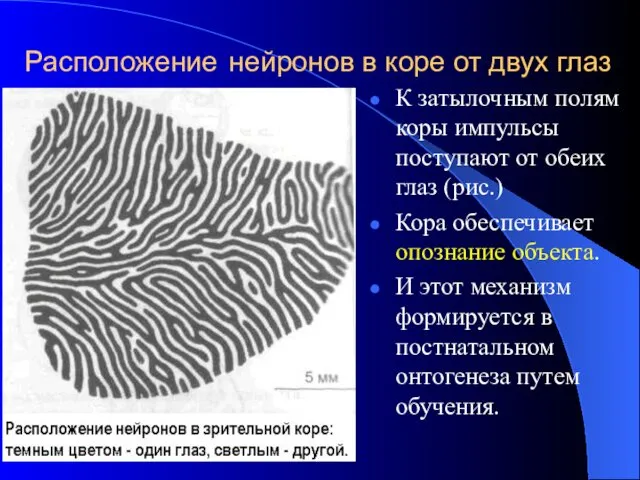
Расположение нейронов в коре от двух глаз
К затылочным полям коры импульсы
Расположение нейронов в коре от двух глаз
К затылочным полям коры импульсы
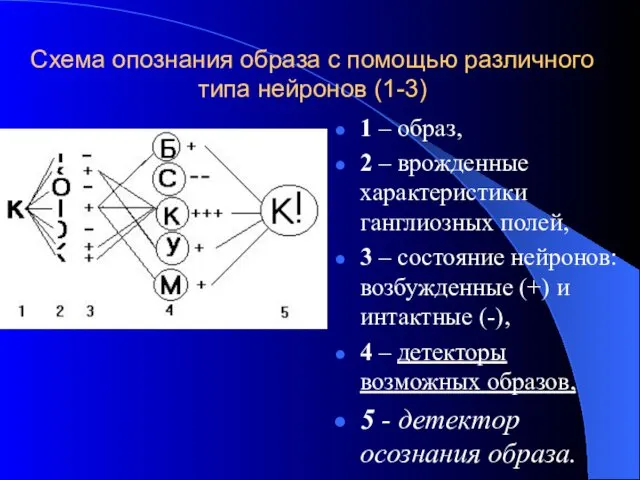
Схема опознания образа с помощью различного типа нейронов (1-3)
1 – образ,
Схема опознания образа с помощью различного типа нейронов (1-3)
1 – образ,
 Побег. Строение и значение
Побег. Строение и значение Класс млекопитающие. Отряд хищные
Класс млекопитающие. Отряд хищные Физиологиялық қызметтердің гормональдық реттелуі. Гормондардың жалпы қасиеттері және қызметтері
Физиологиялық қызметтердің гормональдық реттелуі. Гормондардың жалпы қасиеттері және қызметтері Ощущение и восприятие. Общая и экспериментальная психология
Ощущение и восприятие. Общая и экспериментальная психология Презентация Экологический календарь
Презентация Экологический календарь Общая характеристика низших грибов
Общая характеристика низших грибов Хемосинтез. Автотрофное питание
Хемосинтез. Автотрофное питание Международный день полярного медведя
Международный день полярного медведя Характеристика ферментів мікроорганізмів, які використовують у генній інженерії як інструмент
Характеристика ферментів мікроорганізмів, які використовують у генній інженерії як інструмент Мышечная система
Мышечная система Строение и функции отделов центральной нервной системы. Спинной мозг
Строение и функции отделов центральной нервной системы. Спинной мозг Зеленые водоросли
Зеленые водоросли Внеклеточные образования. Клеточная стенка
Внеклеточные образования. Клеточная стенка Эволюционная теория Ч. Дарвина
Эволюционная теория Ч. Дарвина Общая характеристика мезенхимных тканей. Соединительная ткань. Костная ткань
Общая характеристика мезенхимных тканей. Соединительная ткань. Костная ткань Презентация Человек как житель биосферы
Презентация Человек как житель биосферы Адаптивная роль психики в эволюции животных
Адаптивная роль психики в эволюции животных Adaptation
Adaptation Презентация к уроку Круглые черви 7 класс
Презентация к уроку Круглые черви 7 класс Классификация растений. Водоросли
Классификация растений. Водоросли Введение в физиологию человека. Часть 1
Введение в физиологию человека. Часть 1 Генетика человека и медицинская генетика
Генетика человека и медицинская генетика Основы медицинской электроники. Биоэнергетика
Основы медицинской электроники. Биоэнергетика презентация Самые высокие горы
презентация Самые высокие горы Строение растительной клетки
Строение растительной клетки Группы крови. Резус-фактор
Группы крови. Резус-фактор Наследственность и изменчивость организмов
Наследственность и изменчивость организмов Химический состав клетки
Химический состав клетки