- Внеклеточные образования. Клеточная стенка

Содержание
- 2. Клеточная стенка у растений - это структурное образование, располагающееся по периферии клетки, за пределами плазматической мембраны,
- 3. Основу клеточной стенки растений составляют молекулы целлюлозы, гемицеллюлозы и пектины.
- 4. Клеточные стенки есть также у бактерий
- 5. Клеточные стенки бактерий состоят преимущественно из пептидогликана (муреина)
- 6. Внеклеточный матрикс — внеклеточные структуры соединительной ткани, обеспечивающие механическую поддержку клеток и транспорт химических веществ.
- 7. Главный компонент внеклеточного матрикса — белки. Выделяют 3 группы белков: - протеогликаны (ПГ); - фибриллярные структурные
- 9. Ткань хряща
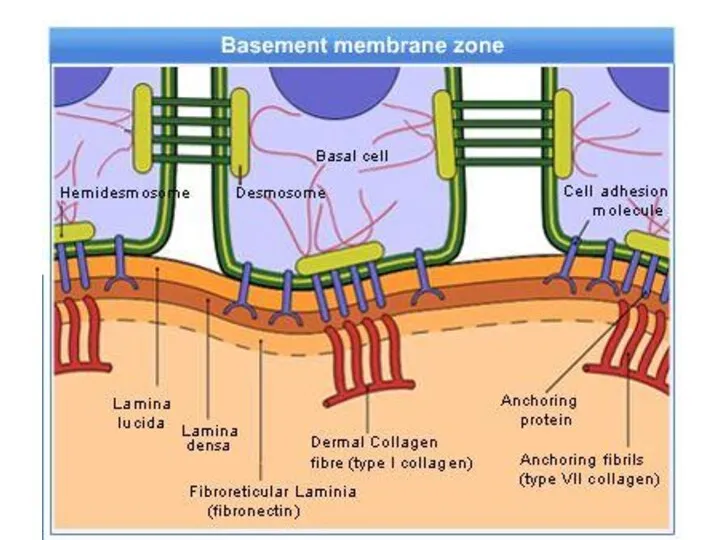
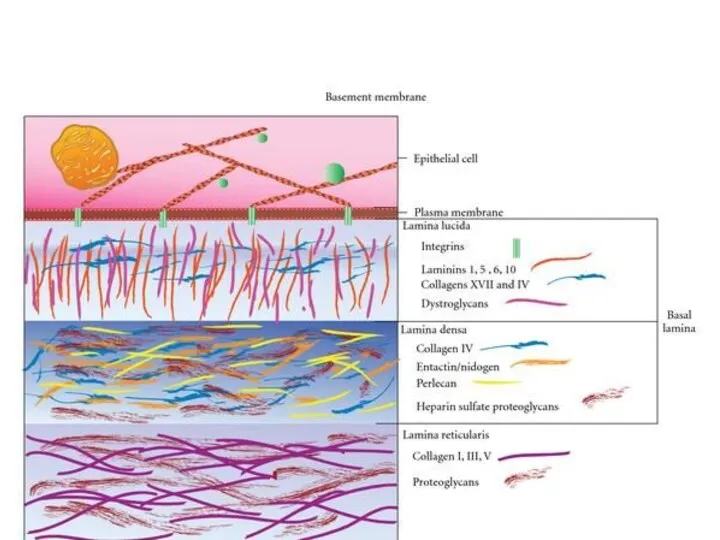
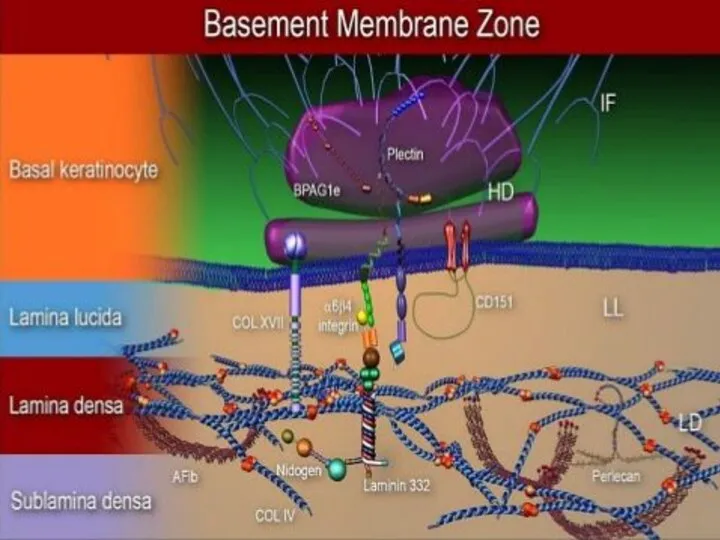
- 12. Базальная мембрана - особый вид внеклеточного матрикса, имеет вид листа или пластинки толщиной 20-200 нм и
- 16. Синтез молекул внеклеточного матрикса Фибробласты синтезируют коллаген (проколлаген), эластин, фибронектин, гликозаминогликаны, протеогликаны и другие компоненты внеклеточного
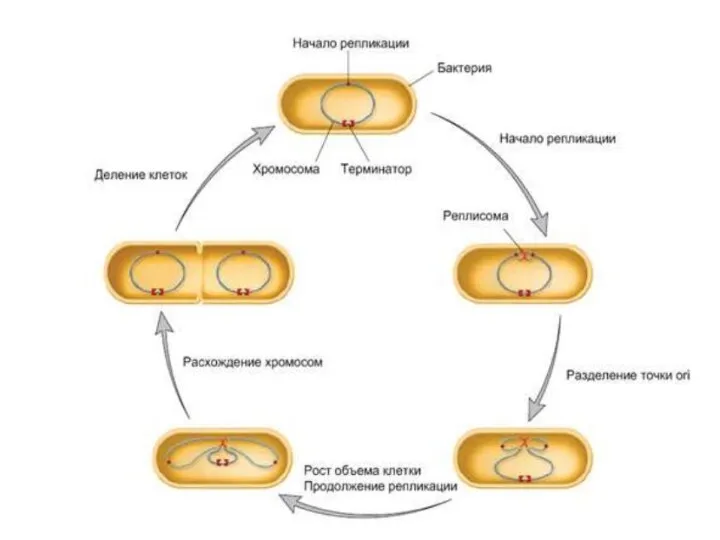
- 17. Клеточный цикл прокариот Процесс клеточного деления у прокариот включает следующие события в определенной очередности: 1) накопление
- 22. Клеточный цикл эукариот
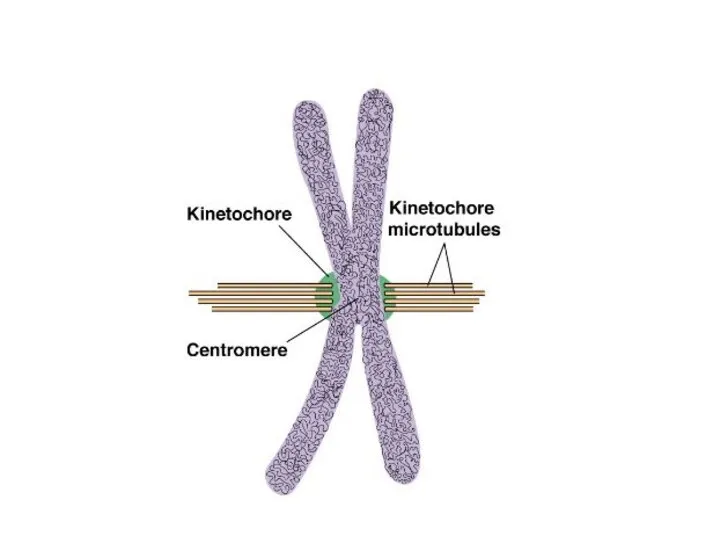
- 25. Митоз
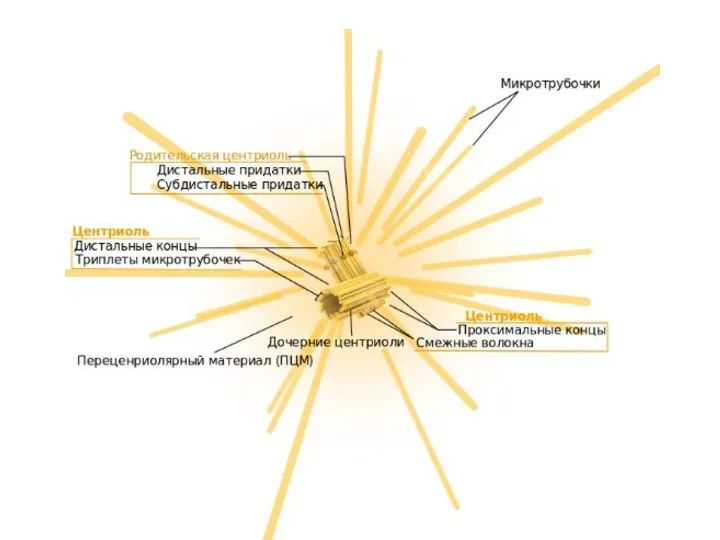
- 26. Клеточный центр (центросома) Немембранный органоид в клетках эукариот, состоит из двух центриолей и перицентриолярного материала. Является
- 28. Профаза конденсация хромосом; исчезновение ядрышек; ядерная оболочка распадается на фрагменты и мелкие мембранные пузырьки; к противоположным
- 29. Начало профазы
- 30. Конец профазы
- 31. Метафаза Перемещение хромосом в экваториальную плоскость и образование метафазной пластинки.
- 34. Анафаза включает процесс расхождения хромосом к полюсам делящейся клетки. Механизм движения хромосом объясняется гипотезой скользящих нитей,
- 35. Телофаза — конечная стадия митоза, в течение которой на полюсах веретена реконструируются дочерние ядра. Перестройка телофазных
- 38. G1-фаза — первая из четырёх фаз клеточного цикла эукариотических клеток. На этом этапе интерфазы клетка увеличивается
- 39. G1-фаза и другие фазы клеточного цикла могут зависеть от факторов, таких как питательные вещества, температура и
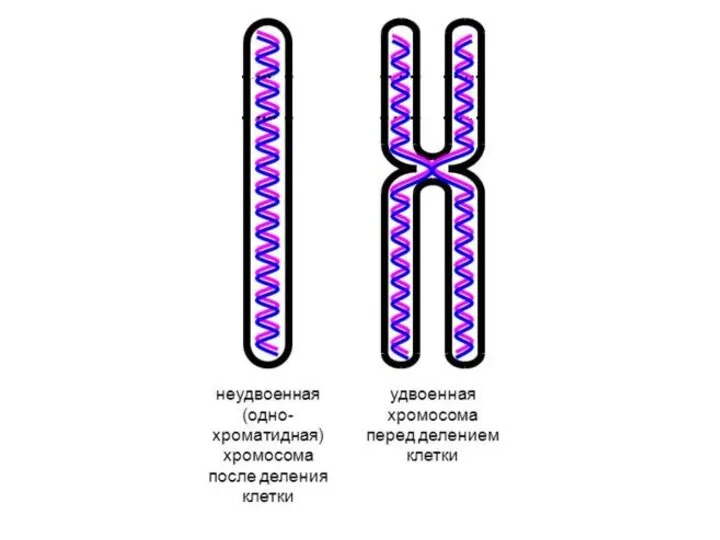
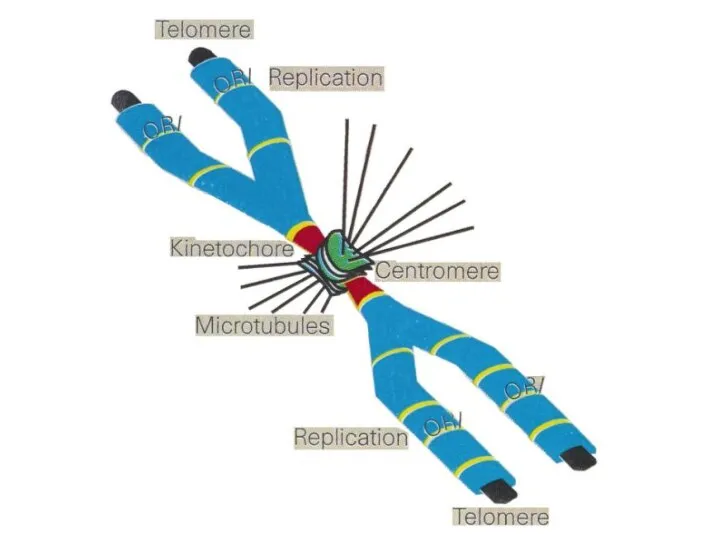
- 40. S-фаза Репликация ДНК. Теперь каждая хромосома будет состоять из 2 хроматид; Удвоение центриолей; Интенсивно синтезируются РНК
- 41. G2 G2-фаза - постсинтетическая, или премитотическая. Во время этой фазы происходят интенсивные процессы биосинтеза, деление митохондрий
- 42. Закономерности репликации хромосом Репликация ДНК и ее закономерности изучаются с помощью бромдезоксиуридина - синтетического нуклеозида, аналога
- 43. A – начало S-фазы B – середина C - конец
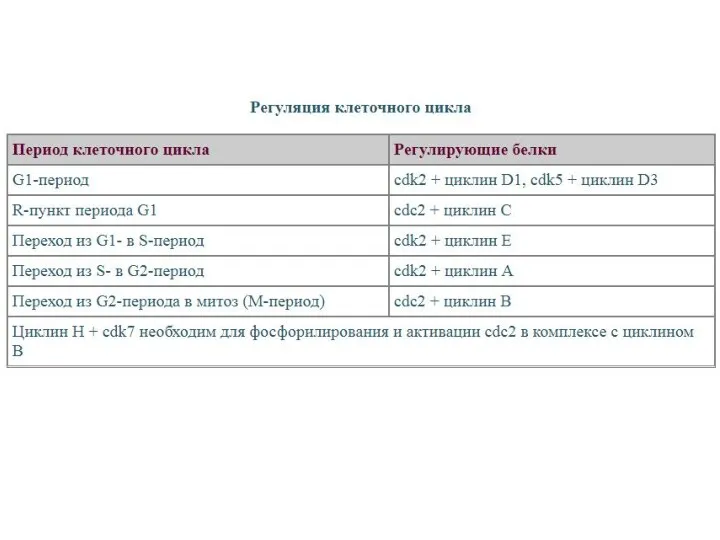
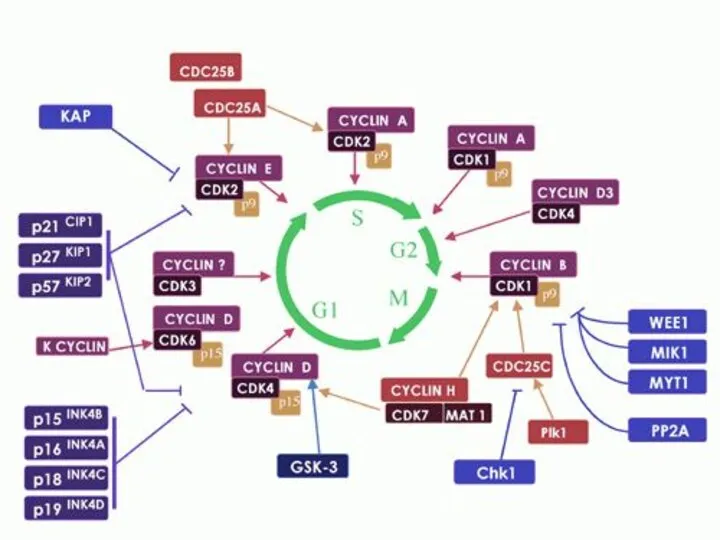
- 44. Прохождение клетки по всем периодам клеточного цикла строго контролируется специальными регуляторными молекулами, которые обеспечивают: 1) прохождение
- 47. Клеточный центр способен удваиваться — каждая из центриолей достраивает возле себя дочернюю.
- 48. Деление митохондрий у красных водорослей
- 49. При исследований деления митохондрий красных водорослей был обнаружен белок FtsZ, принадлежащий к семейству бактериальных белков, участвующих
- 50. Ген FtsZ был потерян в ходе эволюции, его нет уже у ряда простейших, а также у
- 52. Деление пластид
- 53. Взаимодействие клеток
- 54. Клеточные контакты
- 55. Сигнальная трансдукция
- 56. Клеточный ответ на действие гормонов и цитокинов Гормоны — биологически активные вещества органической природы, вырабатывающиеся в
- 57. Действие адреналина
- 58. Цитокины — большая и разнообразная группа небольших по размерам (молекулярная масса от 8 до 80 кДа)
- 59. Действие интерферона
- 60. Апоптоз Апоптоз - регулируемый процесс программируемой клеточной гибели, в результате которого клетка распадается на отдельные апоптотические
- 63. Особенности опухолевых клеток
- 64. Предел Хайфлека – около 50-70 делений
- 65. Особенности метаболизма опухолевой клетки 1. Возрастает активность рибонуклеотидредук-тазы и снижается катаболизм пиримидинов и пуринов, увеличивается синтез
- 66. Появление эмбриональных белков и ферментов Клетки синтезируют, а иногда и секретируют в кровь эмбриональные белки и
- 68. Профаза I мейоза
- 69. Лептотена - конденсация ДНК с образованием хромосом в виде тонких нитей.
- 70. Зиготена - происходит конъюгация — соединение гомологичных хромосом с образованием структур, состоящих из двух соединённых хромосом,
- 71. Пахитена — в некоторых местах гомологичные хромосомы плотно соединяются, образуя хиазмы. В них происходит кроссинговер —
- 72. Кроссинговер
- 73. Механизм кроссинговера
- 74. Диплотена, или диплонема — происходит частичная деконденсация хромосом, при этом часть генома может работать, происходят процессы
- 75. Диакинез — ДНК снова максимально конденсируется, синтетические процессы прекращаются, распадается ядерная оболочка; центриоли расходятся к полюсам;
- 76. Метафаза I — бивалентные хромосомы выстраиваются вдоль экватора клетки. Анафаза I — микротрубочки сокращаются, биваленты делятся,
- 77. Синаптонемный комплекс Синаптонемный ко́мплекс (СК) — белковый комплекс, формирующийся между гомологичными хромосомами (двумя парами сестринских хроматид)
- 80. Длительность: 73-75 дней
- 82. Выход ооцита из фолликула
- 84. Хромосомные перестройки - Делеции - Инсерции - Дупликации - Инверсии - Транслокации
- 86. Изменения структуры хромосом в предшественниках гамет сопровождаются нарушением процесса конъюгации гомологов в мейозе и их последующего
- 88. Конъюгация и последующее расхождение структур, образованных измененными хромосомами, приводит к появлению новых хромосомных перестроек. В результате
- 90. Скачать презентацию

Клеточная стенка у растений - это структурное образование, располагающееся по периферии
Клеточная стенка у растений - это структурное образование, располагающееся по периферии
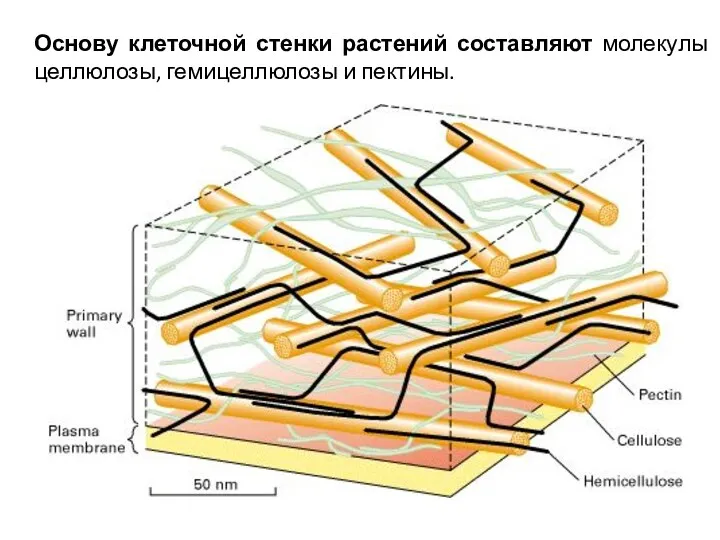
Основу клеточной стенки растений составляют молекулы целлюлозы, гемицеллюлозы и пектины.
Основу клеточной стенки растений составляют молекулы целлюлозы, гемицеллюлозы и пектины.
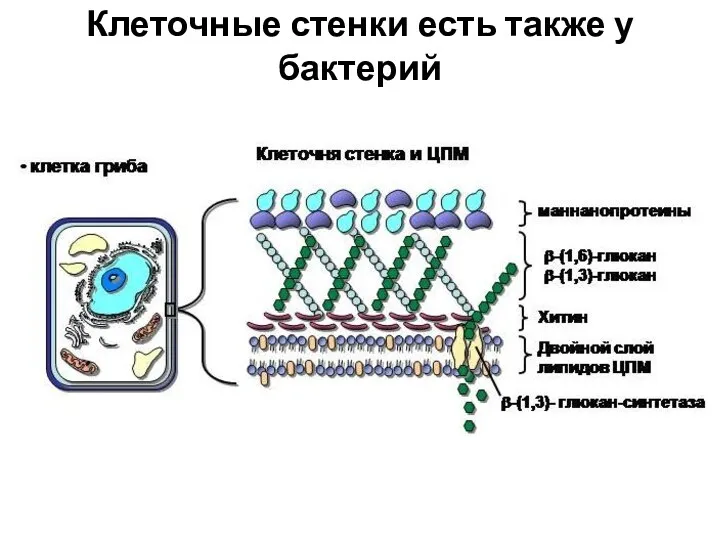
Клеточные стенки есть также у бактерий
Клеточные стенки есть также у бактерий
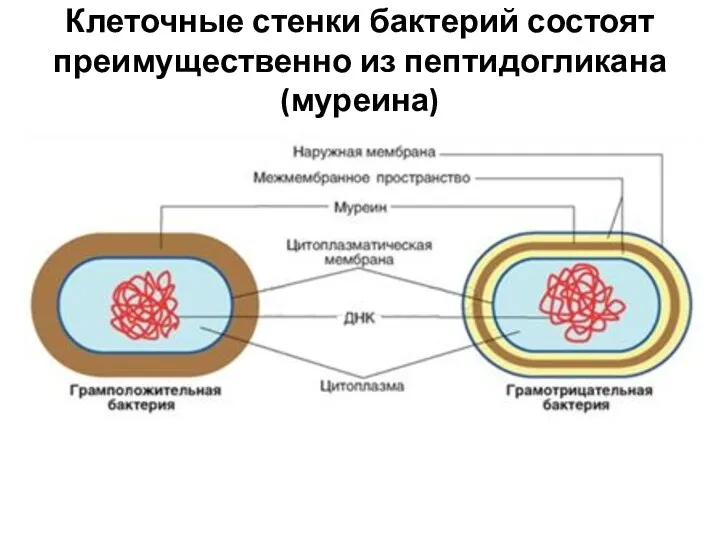
Клеточные стенки бактерий состоят преимущественно из пептидогликана (муреина)
Клеточные стенки бактерий состоят преимущественно из пептидогликана (муреина)
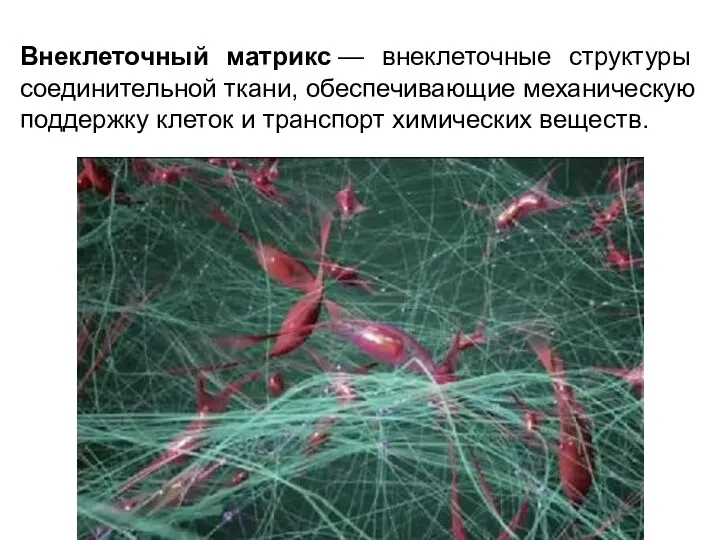
Внеклеточный матрикс — внеклеточные структуры соединительной ткани, обеспечивающие механическую поддержку клеток и
Внеклеточный матрикс — внеклеточные структуры соединительной ткани, обеспечивающие механическую поддержку клеток и
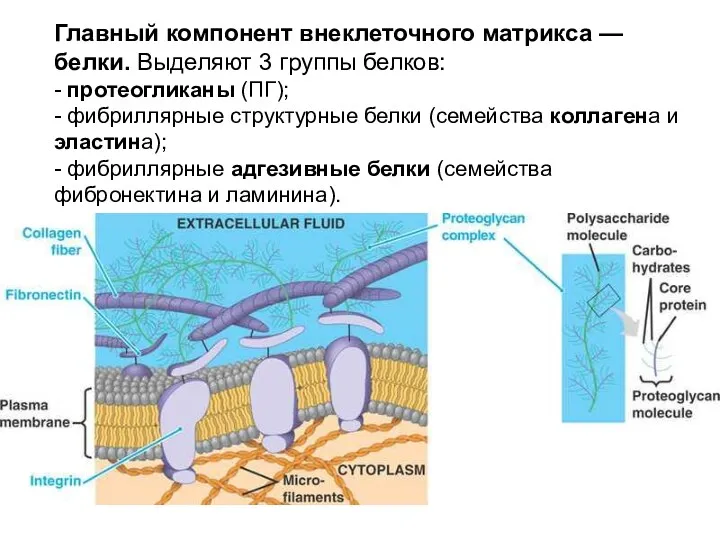
Главный компонент внеклеточного матрикса — белки. Выделяют 3 группы белков:
- протеогликаны
Главный компонент внеклеточного матрикса — белки. Выделяют 3 группы белков: - протеогликаны
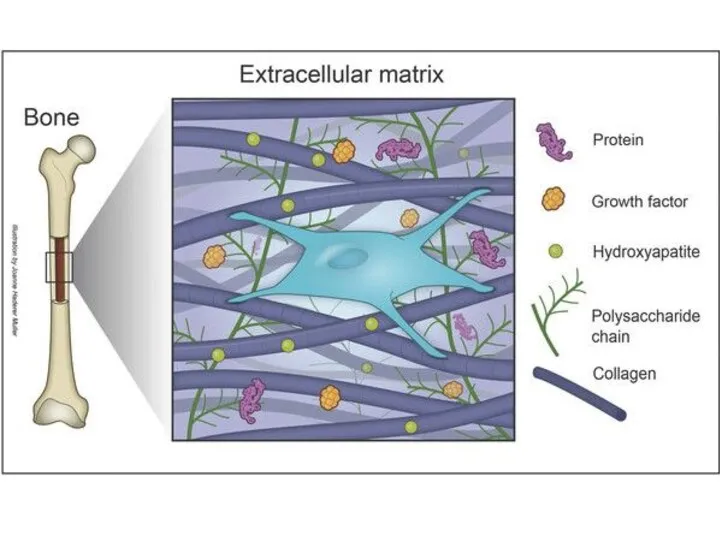

Ткань хряща
Ткань хряща
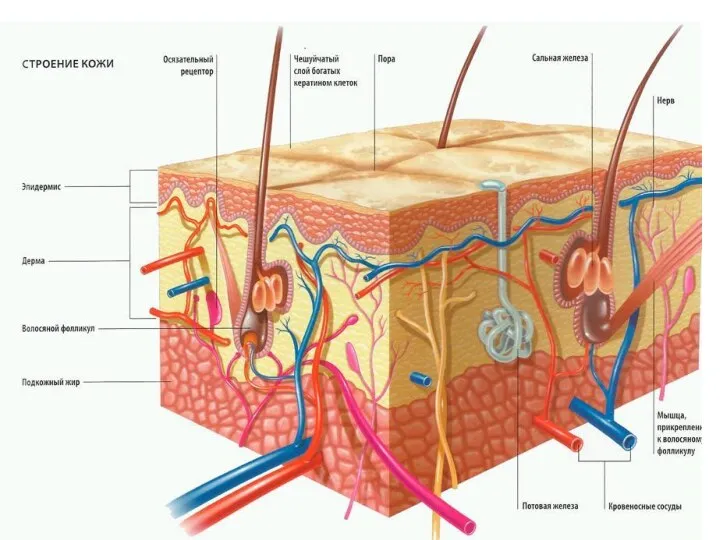
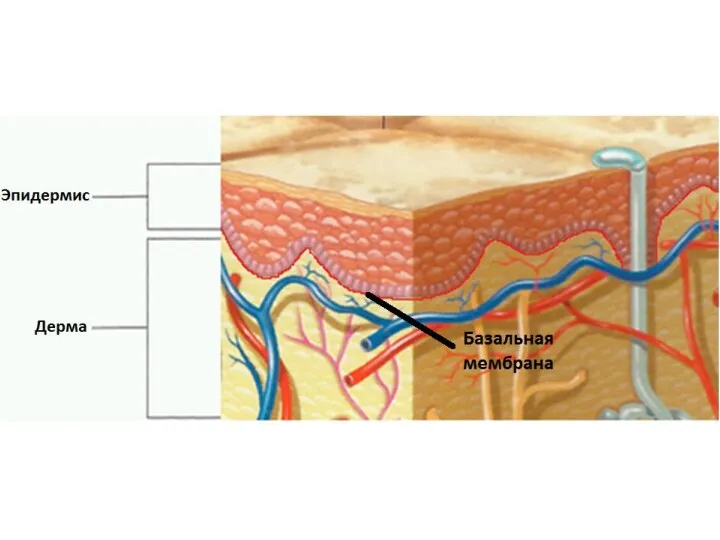

Базальная мембрана - особый вид внеклеточного матрикса, имеет вид листа или
Базальная мембрана - особый вид внеклеточного матрикса, имеет вид листа или

Синтез молекул внеклеточного матрикса
Фибробласты синтезируют коллаген (проколлаген), эластин, фибронектин, гликозаминогликаны, протеогликаны
Синтез молекул внеклеточного матрикса
Фибробласты синтезируют коллаген (проколлаген), эластин, фибронектин, гликозаминогликаны, протеогликаны

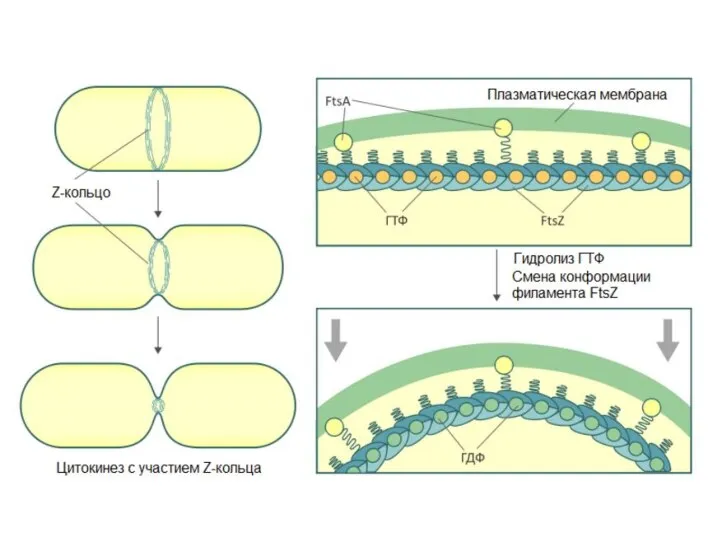
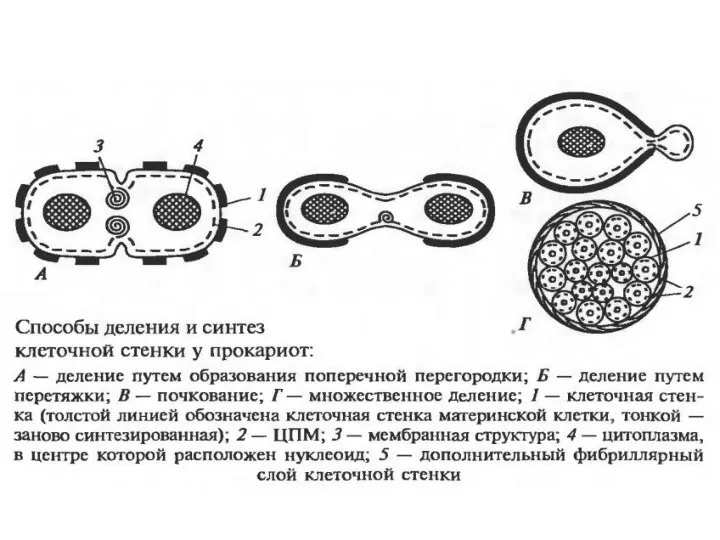
Клеточный цикл прокариот
Процесс клеточного деления у прокариот включает следующие события в
Клеточный цикл прокариот
Процесс клеточного деления у прокариот включает следующие события в
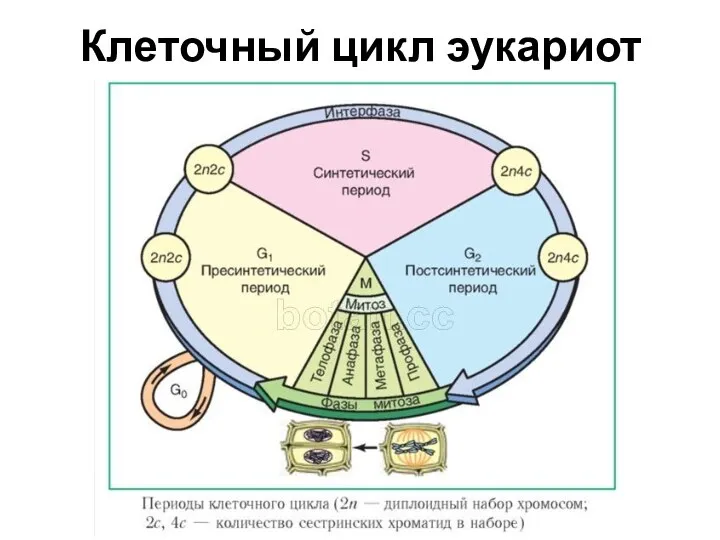
Клеточный цикл эукариот
Клеточный цикл эукариот
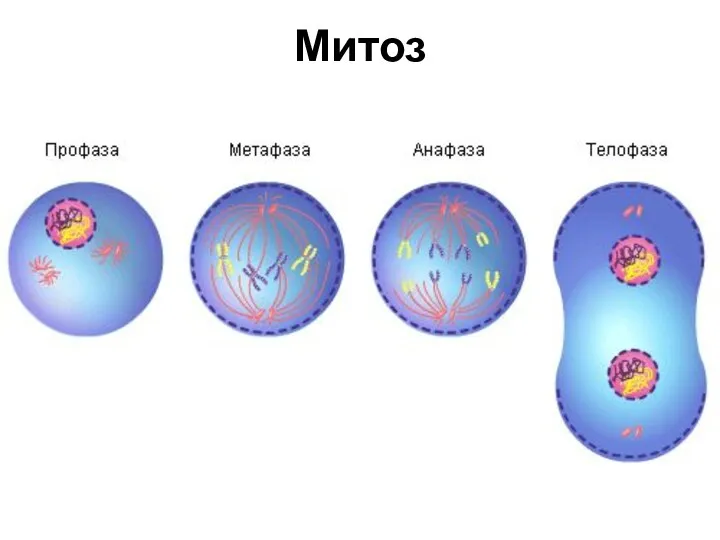
Митоз
Митоз

Клеточный центр (центросома)
Немембранный органоид в клетках эукариот, состоит из двух центриолей и перицентриолярного материала.
Клеточный центр (центросома)
Немембранный органоид в клетках эукариот, состоит из двух центриолей и перицентриолярного материала.

Профаза
конденсация хромосом;
исчезновение ядрышек;
ядерная оболочка распадается на фрагменты и
Профаза
конденсация хромосом;
исчезновение ядрышек;
ядерная оболочка распадается на фрагменты и
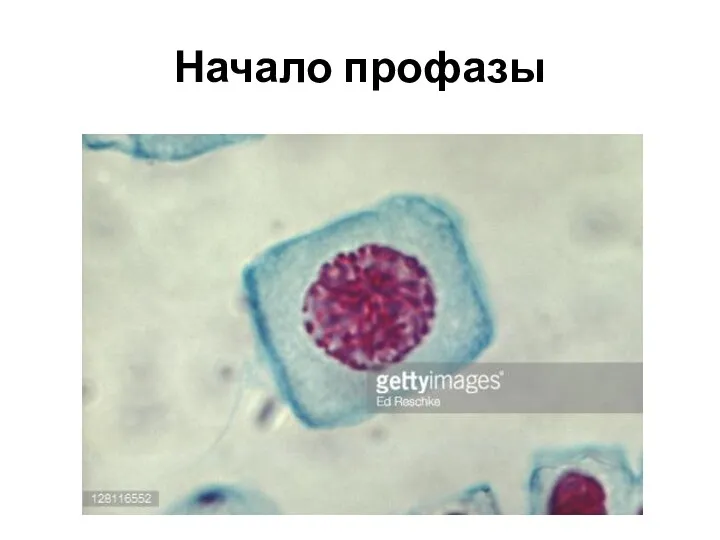
Начало профазы
Начало профазы
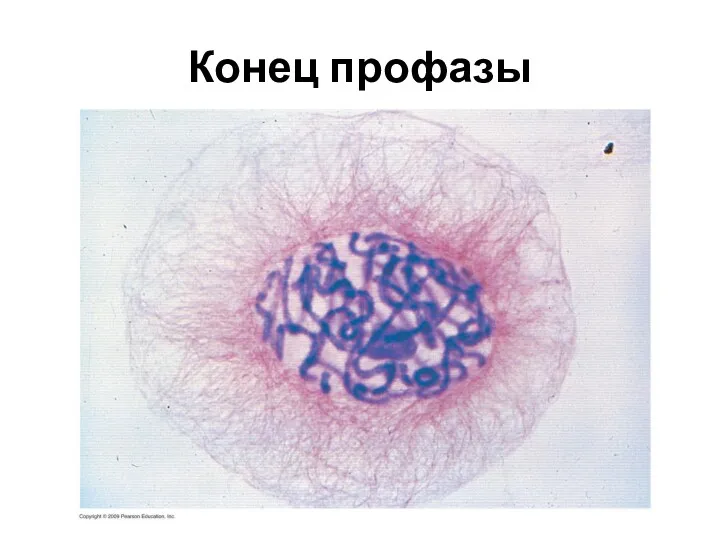
Конец профазы
Конец профазы
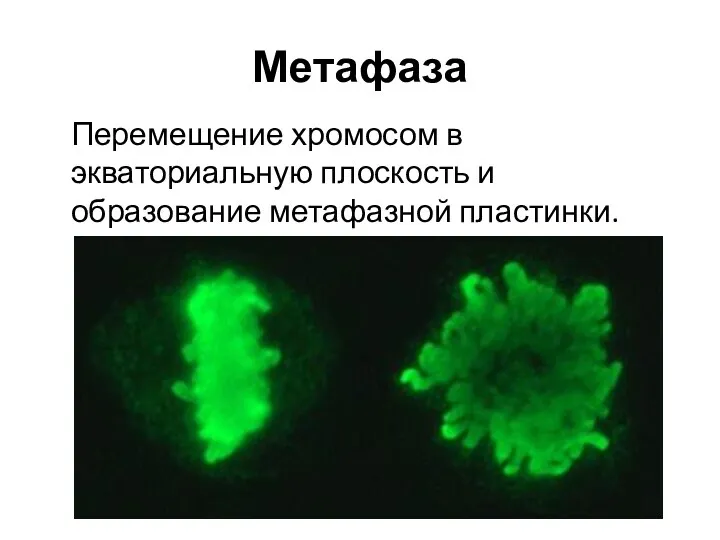
Метафаза
Перемещение хромосом в экваториальную плоскость и образование метафазной пластинки.
Метафаза
Перемещение хромосом в экваториальную плоскость и образование метафазной пластинки.
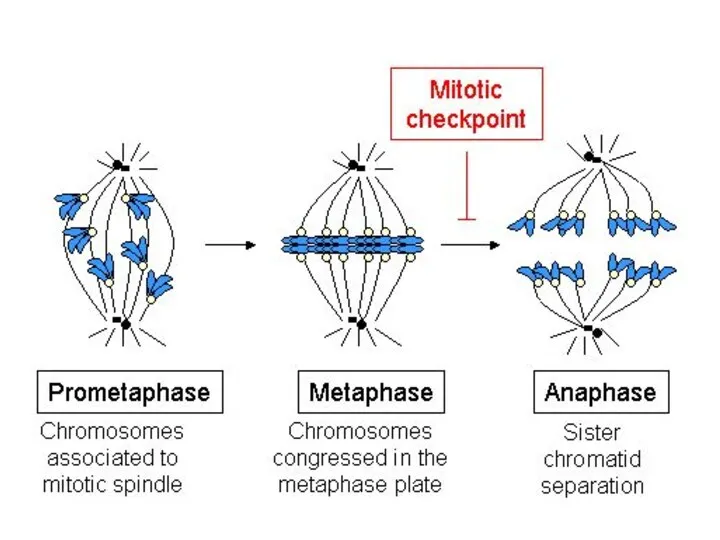
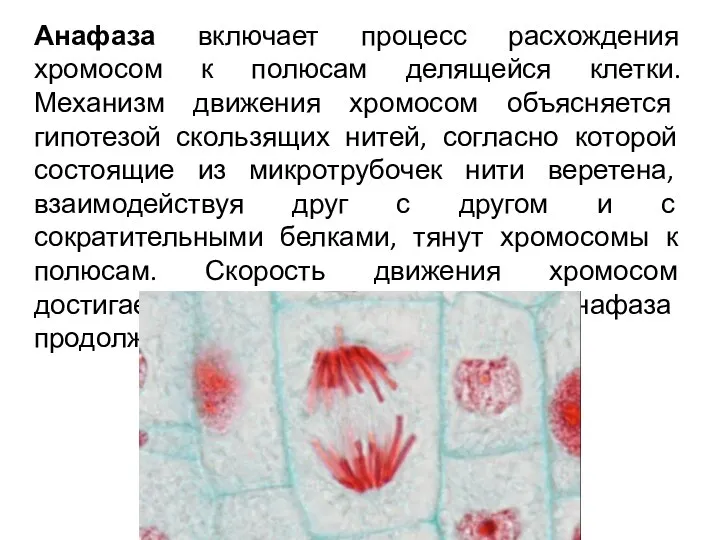
Анафаза включает процесс расхождения хромосом к полюсам делящейся клетки. Механизм движения
Анафаза включает процесс расхождения хромосом к полюсам делящейся клетки. Механизм движения

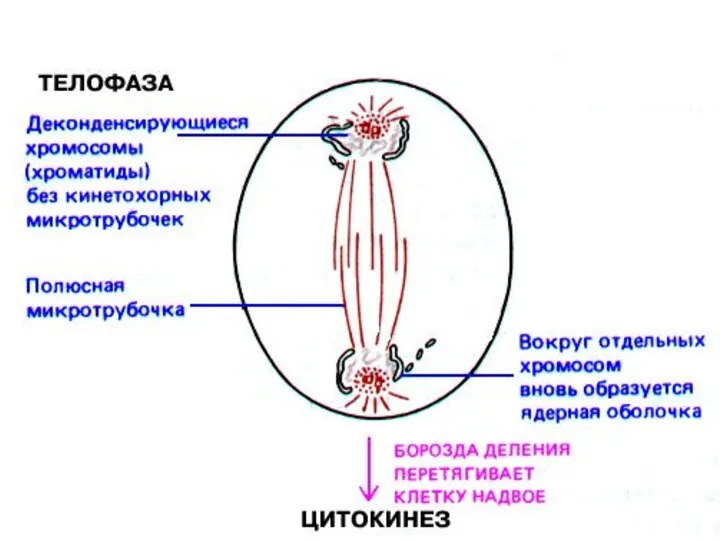
Телофаза — конечная стадия митоза, в течение которой на полюсах веретена
Телофаза — конечная стадия митоза, в течение которой на полюсах веретена
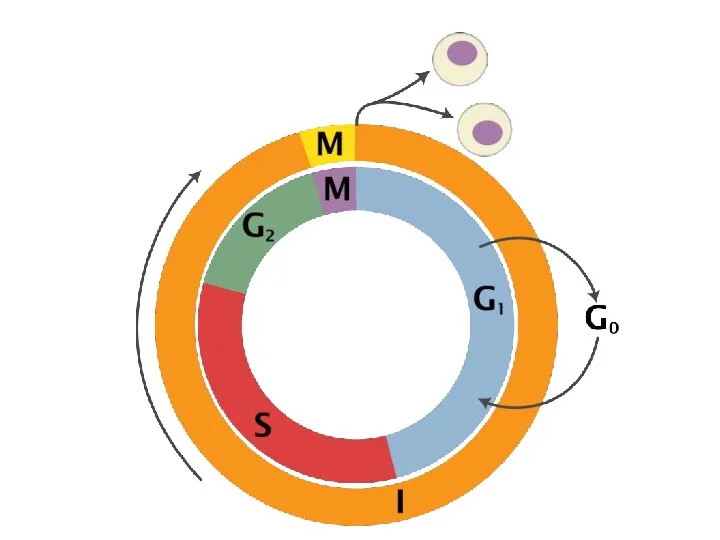

G1-фаза — первая из четырёх фаз клеточного цикла эукариотических клеток. На этом этапе интерфазы клетка увеличивается
G1-фаза — первая из четырёх фаз клеточного цикла эукариотических клеток. На этом этапе интерфазы клетка увеличивается

G1-фаза и другие фазы клеточного цикла могут зависеть от факторов, таких как питательные
G1-фаза и другие фазы клеточного цикла могут зависеть от факторов, таких как питательные

S-фаза
Репликация ДНК. Теперь каждая хромосома будет состоять из 2 хроматид;
S-фаза
Репликация ДНК. Теперь каждая хромосома будет состоять из 2 хроматид;

G2
G2-фаза - постсинтетическая, или премитотическая. Во время этой фазы происходят интенсивные процессы
G2
G2-фаза - постсинтетическая, или премитотическая. Во время этой фазы происходят интенсивные процессы

Закономерности репликации хромосом
Репликация ДНК и ее закономерности изучаются с помощью бромдезоксиуридина -
Закономерности репликации хромосом
Репликация ДНК и ее закономерности изучаются с помощью бромдезоксиуридина -
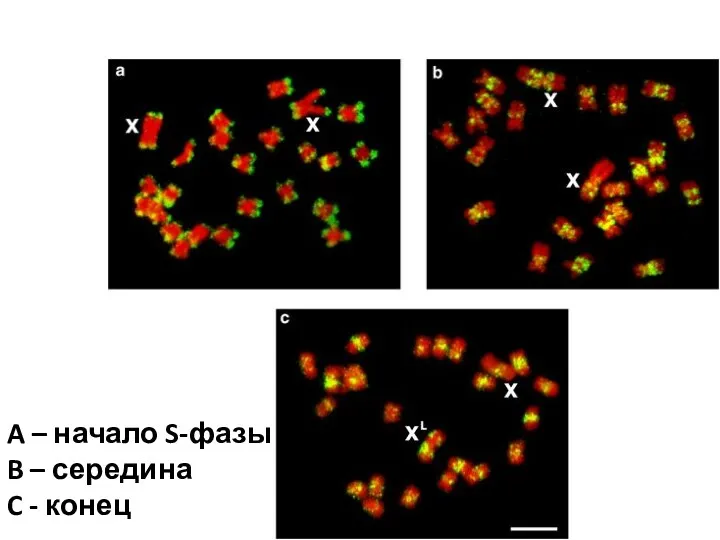
A – начало S-фазы
B – середина
C - конец
A – начало S-фазы
B – середина
C - конец

Прохождение клетки по всем периодам клеточного цикла строго контролируется специальными регуляторными
Прохождение клетки по всем периодам клеточного цикла строго контролируется специальными регуляторными
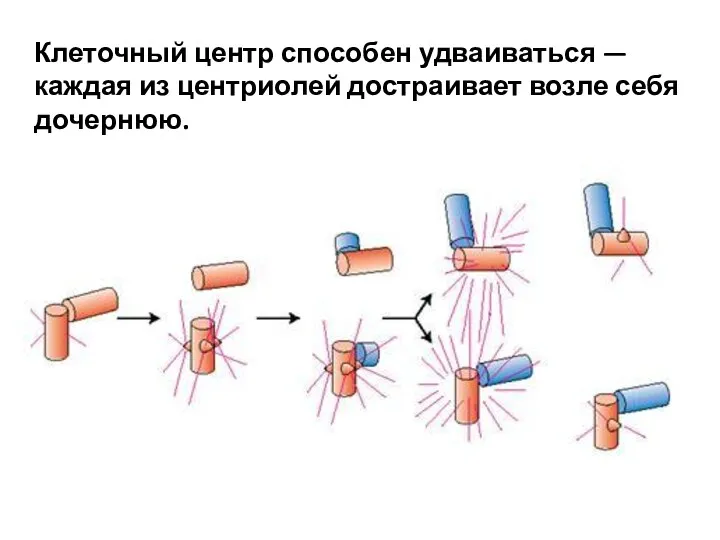
Клеточный центр способен удваиваться — каждая из центриолей достраивает возле себя
Клеточный центр способен удваиваться — каждая из центриолей достраивает возле себя
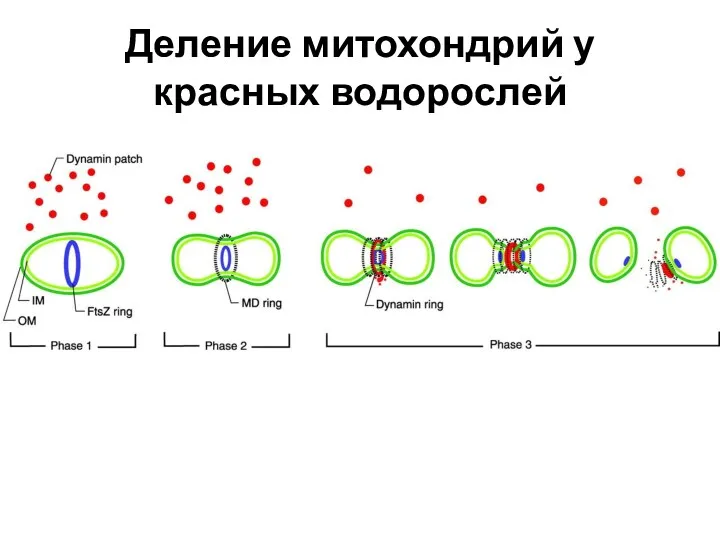
Деление митохондрий у красных водорослей
Деление митохондрий у красных водорослей

При исследований деления митохондрий красных водорослей был обнаружен белок FtsZ, принадлежащий к
При исследований деления митохондрий красных водорослей был обнаружен белок FtsZ, принадлежащий к
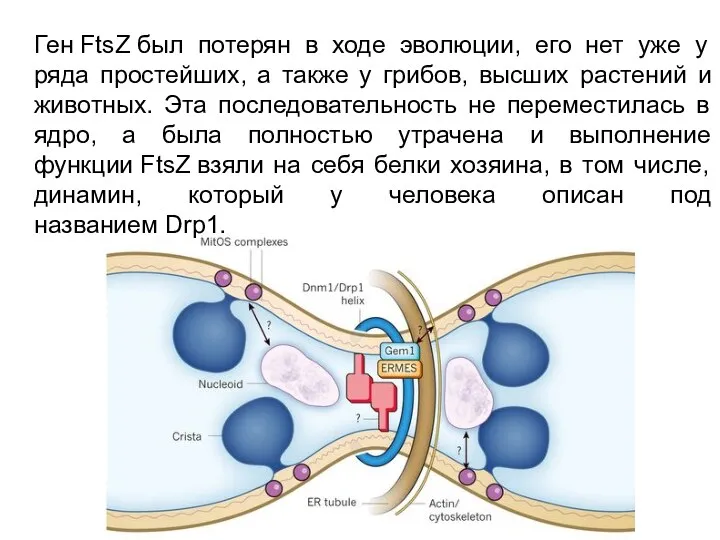
Ген FtsZ был потерян в ходе эволюции, его нет уже у ряда простейших,
Ген FtsZ был потерян в ходе эволюции, его нет уже у ряда простейших,
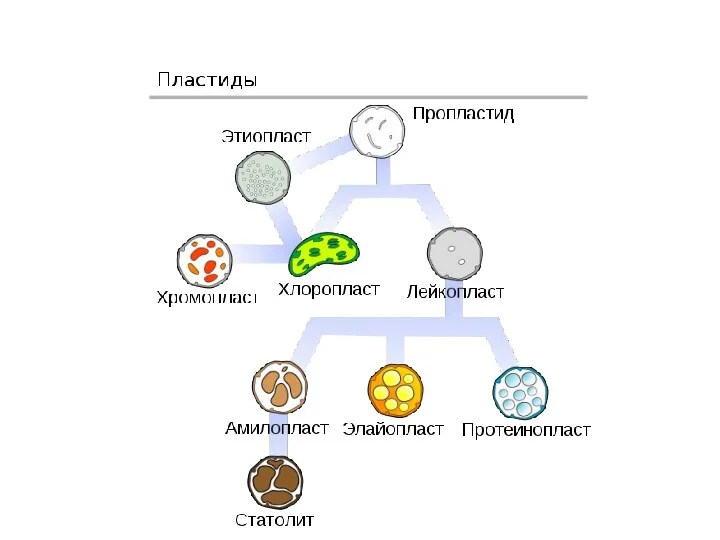
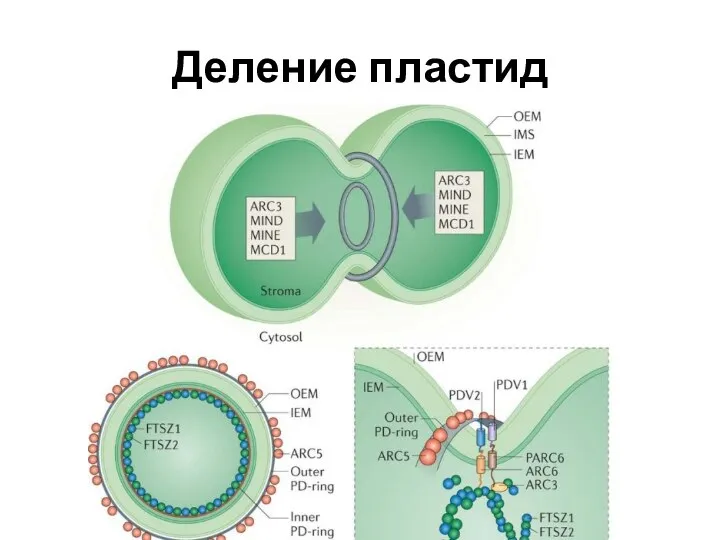
Деление пластид
Деление пластид
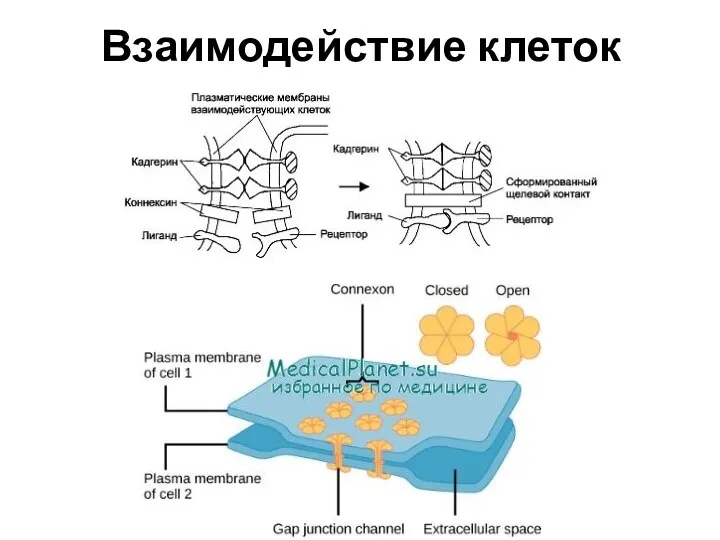
Взаимодействие клеток
Взаимодействие клеток
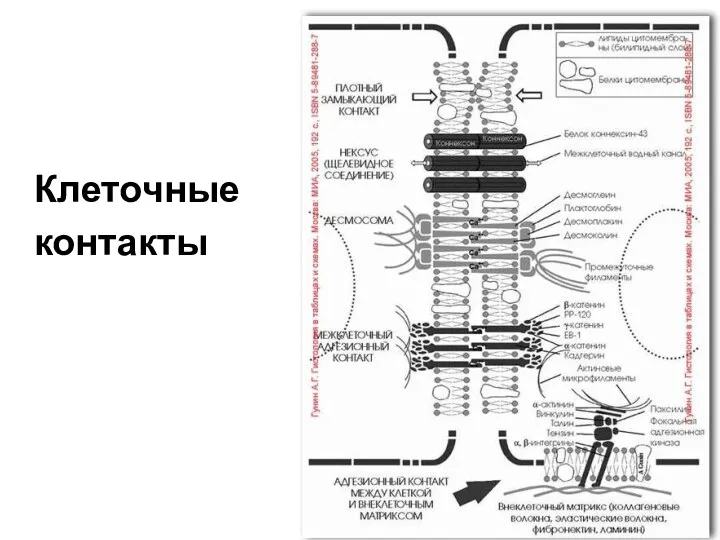
Клеточные
контакты
Клеточные
контакты
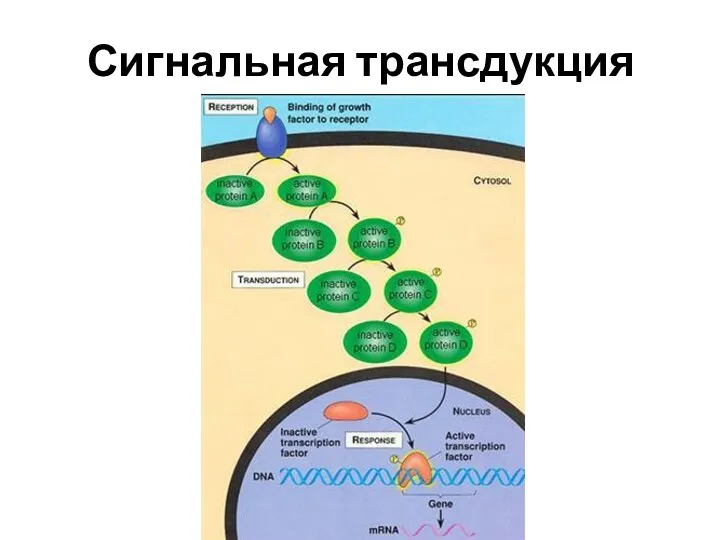
Сигнальная трансдукция
Сигнальная трансдукция

Клеточный ответ на действие гормонов и цитокинов
Гормоны — биологически активные вещества органической природы, вырабатывающиеся
Клеточный ответ на действие гормонов и цитокинов
Гормоны — биологически активные вещества органической природы, вырабатывающиеся
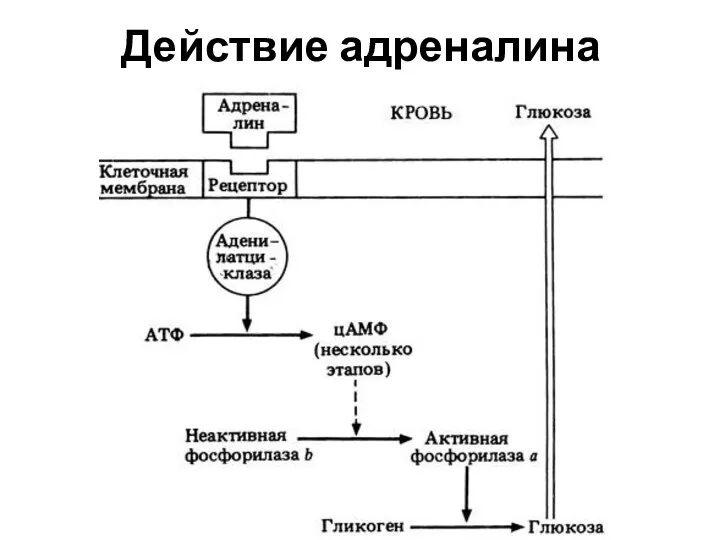
Действие адреналина
Действие адреналина

Цитокины — большая и разнообразная группа небольших по размерам (молекулярная масса
Цитокины — большая и разнообразная группа небольших по размерам (молекулярная масса
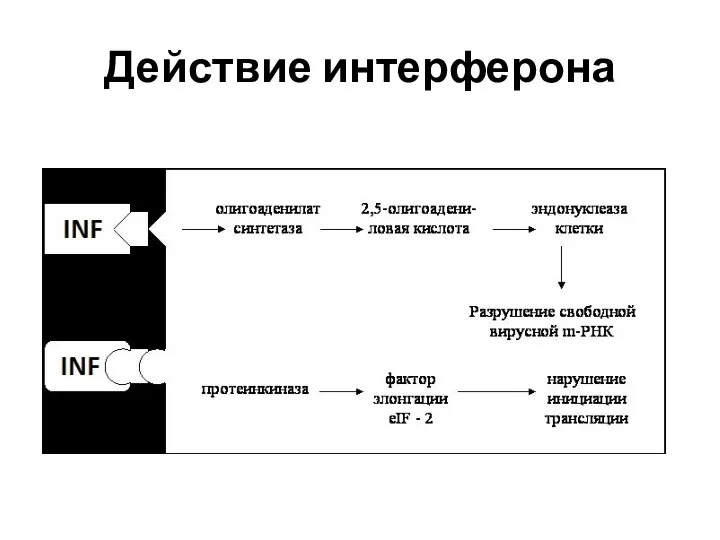
Действие интерферона
Действие интерферона

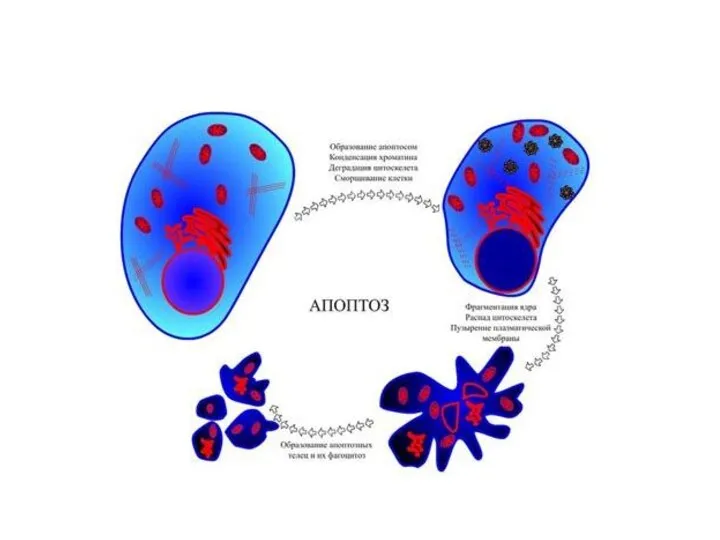
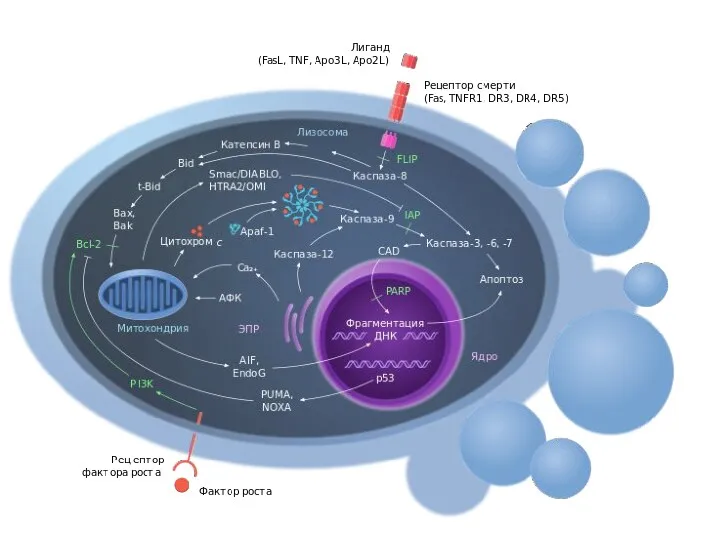
Апоптоз
Апоптоз - регулируемый процесс программируемой клеточной гибели, в результате которого клетка распадается
Апоптоз
Апоптоз - регулируемый процесс программируемой клеточной гибели, в результате которого клетка распадается
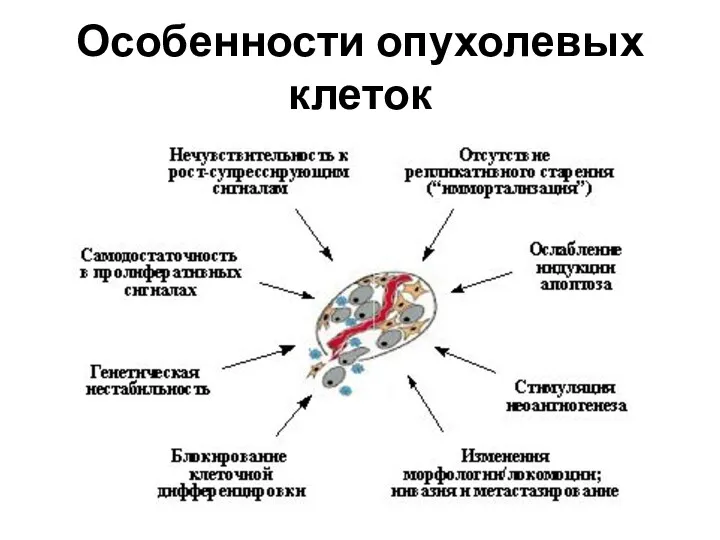
Особенности опухолевых клеток
Особенности опухолевых клеток
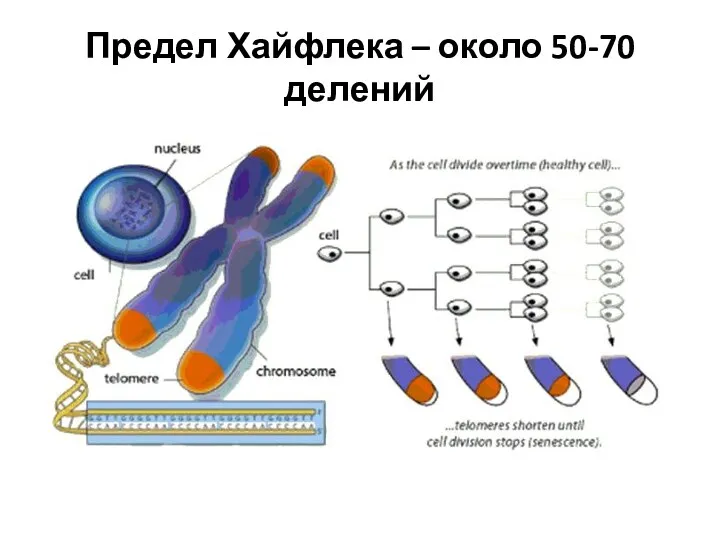
Предел Хайфлека – около 50-70 делений
Предел Хайфлека – около 50-70 делений

Особенности метаболизма опухолевой клетки
1. Возрастает активность рибонуклеотидредук-тазы и снижается катаболизм пиримидинов
Особенности метаболизма опухолевой клетки
1. Возрастает активность рибонуклеотидредук-тазы и снижается катаболизм пиримидинов

Появление эмбриональных белков и ферментов
Клетки синтезируют, а иногда и секретируют в
Появление эмбриональных белков и ферментов
Клетки синтезируют, а иногда и секретируют в
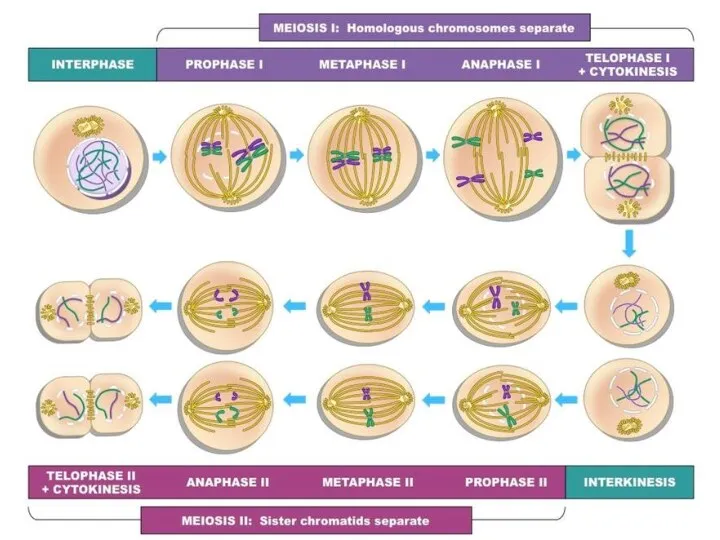
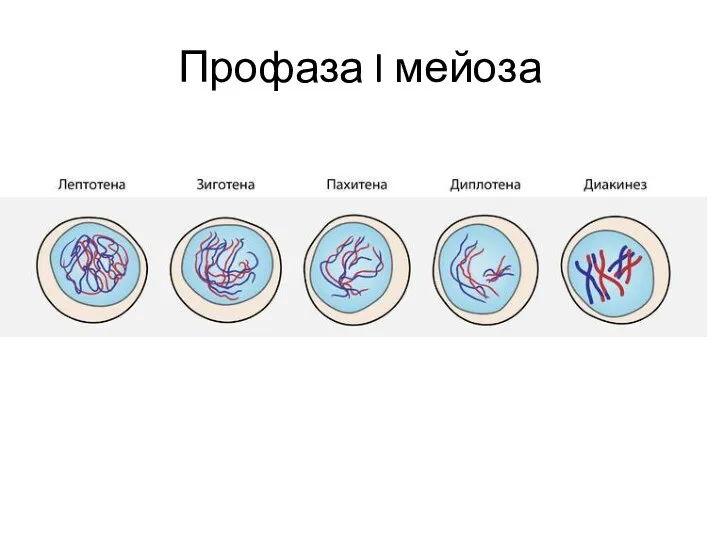
Профаза I мейоза
Профаза I мейоза
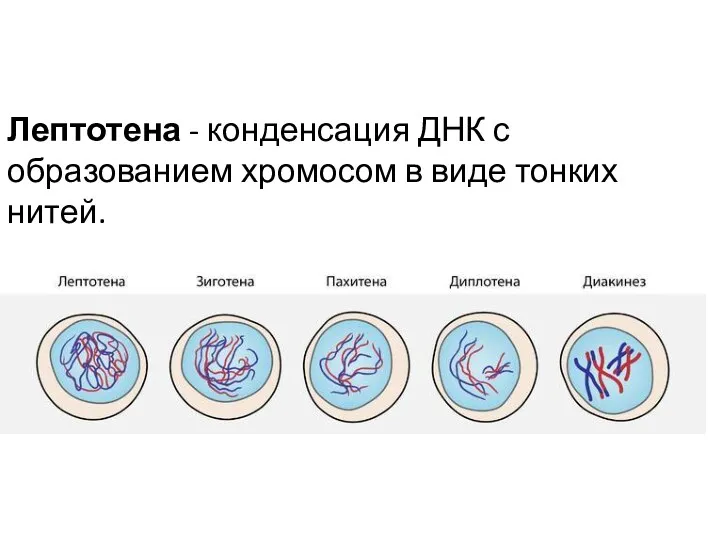
Лептотена - конденсация ДНК с образованием хромосом в виде тонких нитей.
Лептотена - конденсация ДНК с образованием хромосом в виде тонких нитей.
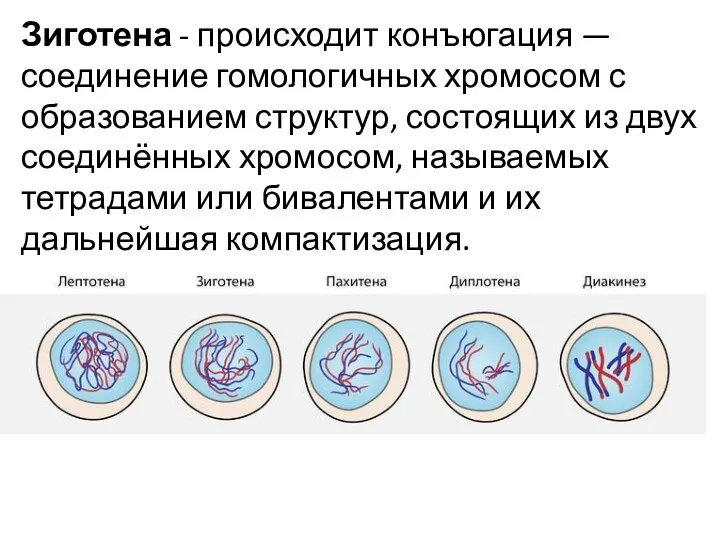
Зиготена - происходит конъюгация — соединение гомологичных хромосом с образованием структур, состоящих из
Зиготена - происходит конъюгация — соединение гомологичных хромосом с образованием структур, состоящих из
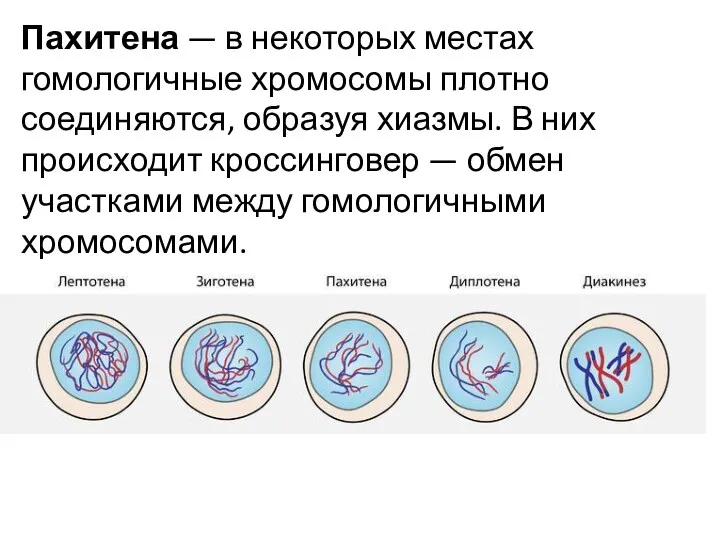
Пахитена — в некоторых местах гомологичные хромосомы плотно соединяются, образуя хиазмы. В них
Пахитена — в некоторых местах гомологичные хромосомы плотно соединяются, образуя хиазмы. В них
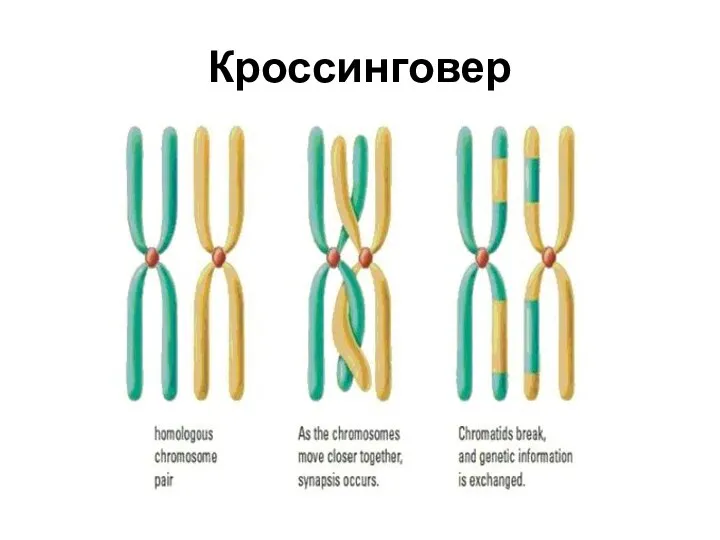
Кроссинговер
Кроссинговер
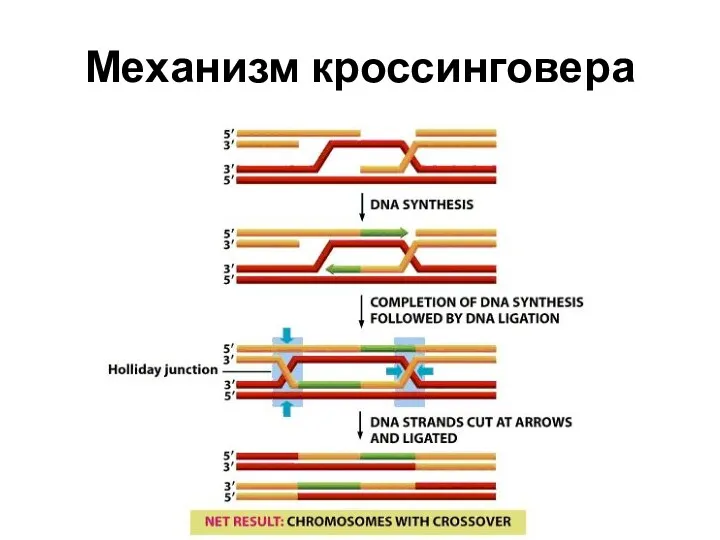
Механизм кроссинговера
Механизм кроссинговера
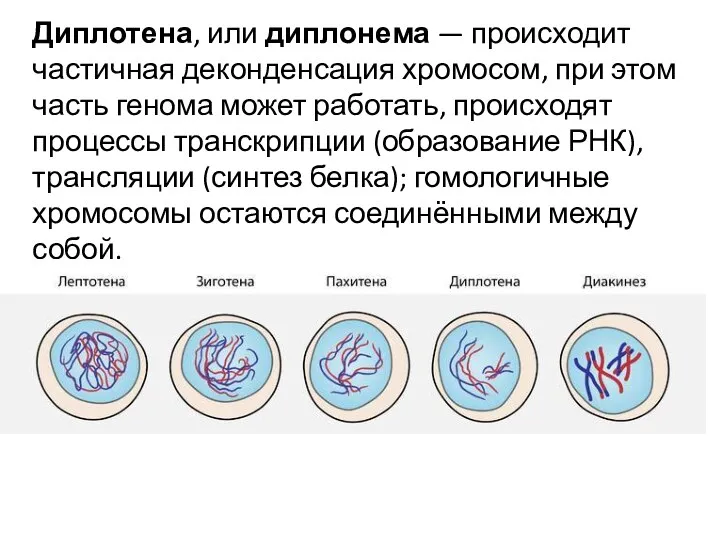
Диплотена, или диплонема — происходит частичная деконденсация хромосом, при этом часть генома может
Диплотена, или диплонема — происходит частичная деконденсация хромосом, при этом часть генома может
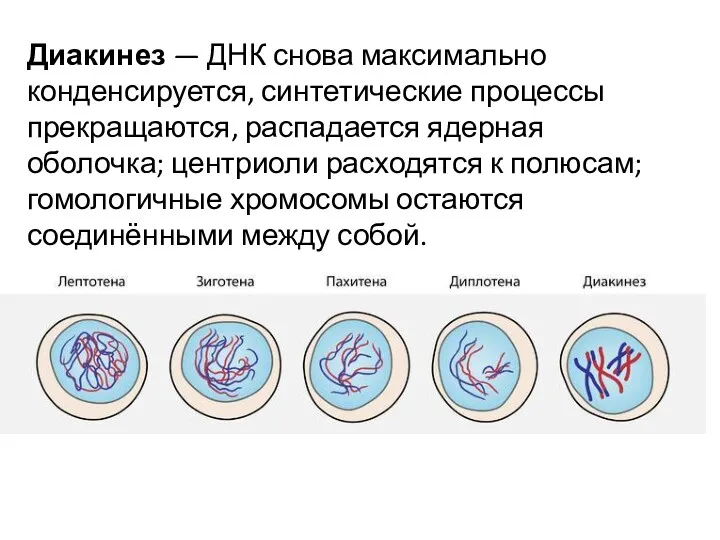
Диакинез — ДНК снова максимально конденсируется, синтетические процессы прекращаются, распадается ядерная оболочка; центриоли
Диакинез — ДНК снова максимально конденсируется, синтетические процессы прекращаются, распадается ядерная оболочка; центриоли

Метафаза I — бивалентные хромосомы выстраиваются вдоль экватора клетки.
Анафаза I — микротрубочки сокращаются, биваленты делятся,
Метафаза I — бивалентные хромосомы выстраиваются вдоль экватора клетки.
Анафаза I — микротрубочки сокращаются, биваленты делятся,

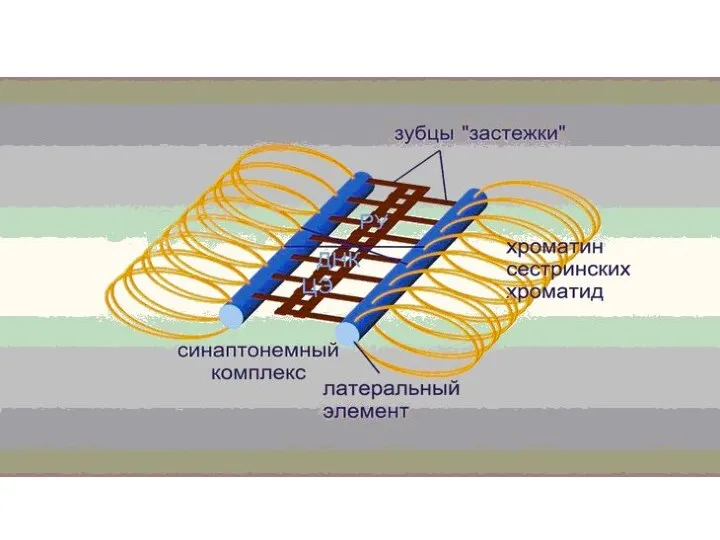
Синаптонемный комплекс
Синаптонемный ко́мплекс (СК) — белковый комплекс, формирующийся между гомологичными хромосомами (двумя парами сестринских хроматид)
Синаптонемный комплекс
Синаптонемный ко́мплекс (СК) — белковый комплекс, формирующийся между гомологичными хромосомами (двумя парами сестринских хроматид)
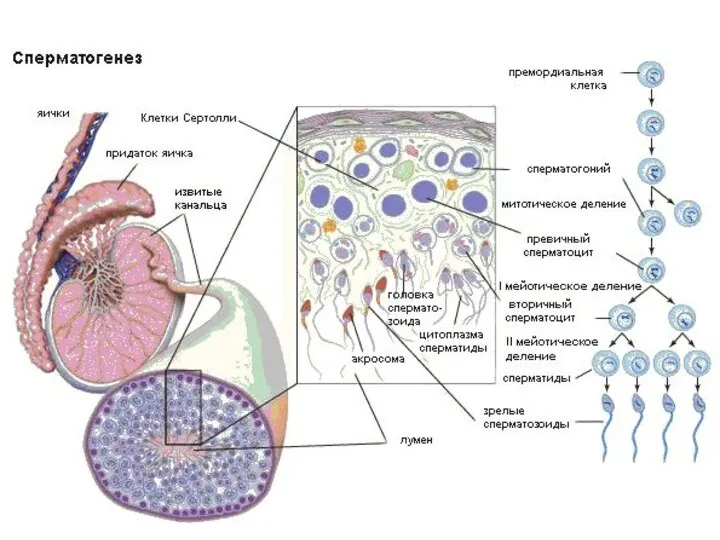
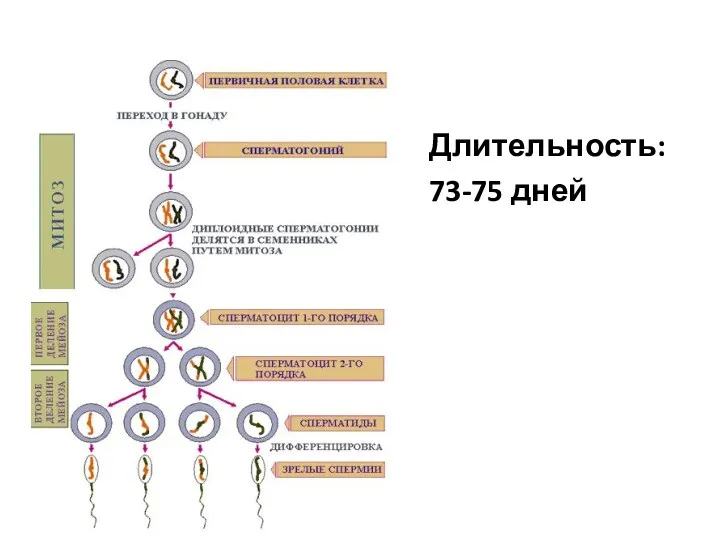
Длительность:
73-75 дней
Длительность:
73-75 дней
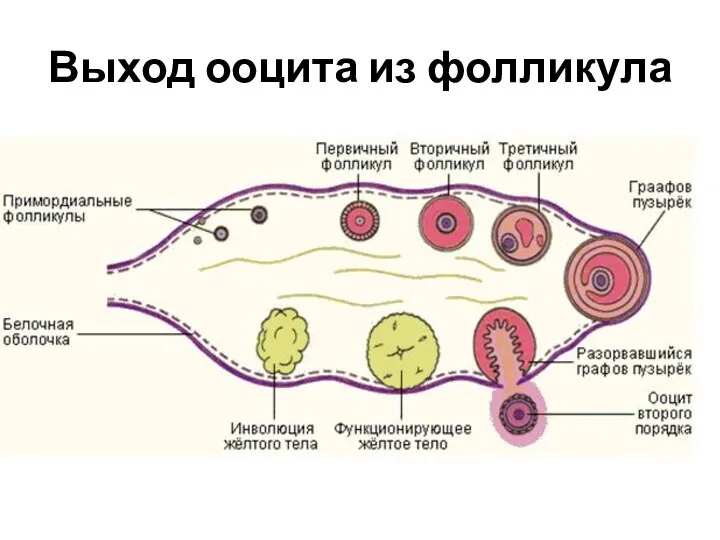
Выход ооцита из фолликула
Выход ооцита из фолликула

Хромосомные перестройки
- Делеции
- Инсерции
- Дупликации
- Инверсии
- Транслокации
Хромосомные перестройки
- Делеции
- Инсерции
- Дупликации
- Инверсии
- Транслокации
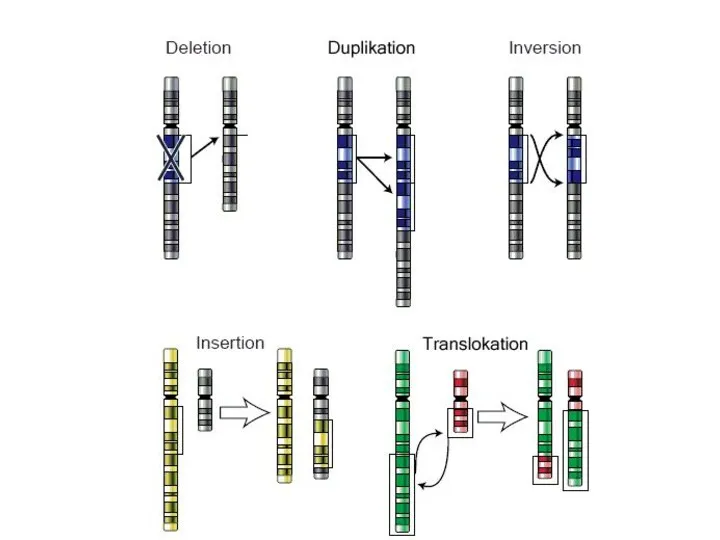

Изменения структуры хромосом в предшественниках гамет сопровождаются нарушением процесса конъюгации гомологов
Изменения структуры хромосом в предшественниках гамет сопровождаются нарушением процесса конъюгации гомологов


Конъюгация и последующее расхождение структур, образованных измененными хромосомами, приводит к появлению
Конъюгация и последующее расхождение структур, образованных измененными хромосомами, приводит к появлению
 7 самых ядовитых растений России
7 самых ядовитых растений России Строение сердца. Сердечный цикл
Строение сердца. Сердечный цикл Бактерии и их виды (часть 1)
Бактерии и их виды (часть 1) Клуб Почемучка. Что происходит с насекомыми зимой
Клуб Почемучка. Что происходит с насекомыми зимой Мінеральне та повітряне живлення рослин
Мінеральне та повітряне живлення рослин Тамырҙар. 4-се класс
Тамырҙар. 4-се класс ОГЭ-2020 по биологии. Новые требования
ОГЭ-2020 по биологии. Новые требования Китоподібні або дельфіноподібні
Китоподібні або дельфіноподібні Клеточная теория. Цитология
Клеточная теория. Цитология Паразитические плоские черви. Верно ли утверждение?
Паразитические плоские черви. Верно ли утверждение? Презентация к уроку по теме Деление клетки
Презентация к уроку по теме Деление клетки Жизнь в мезозое. Триасовый период
Жизнь в мезозое. Триасовый период Structure Of DNA & RNA
Structure Of DNA & RNA Зоология – наука о животных. Подготовка к ОГЭ и ЕГЭ по биологии
Зоология – наука о животных. Подготовка к ОГЭ и ЕГЭ по биологии Тип хордовые
Тип хордовые Многообразие и значение Пресмыкающихся
Многообразие и значение Пресмыкающихся Гаметогенез – процесс образования половых клеток
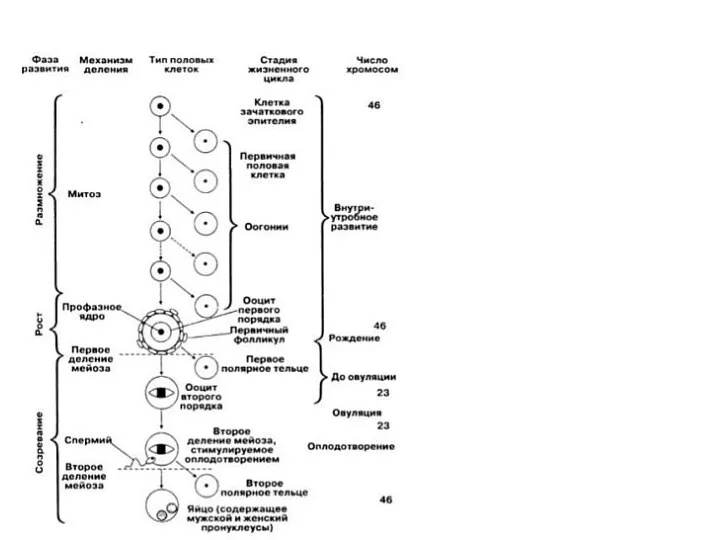
Гаметогенез – процесс образования половых клеток Партеногенез
Партеногенез Грудобрюшная диафрагма
Грудобрюшная диафрагма ВКР: Иммобилизованный биокатализатор на основе адгезированных амидазосодержащих клеток родококков для синтеза акриловой кислоты
ВКР: Иммобилизованный биокатализатор на основе адгезированных амидазосодержащих клеток родококков для синтеза акриловой кислоты Марикультура. Поняття про марикультуру. Види вирощування. Мануфактура в найбільш розвинених країнах. Перспективи розвитку
Марикультура. Поняття про марикультуру. Види вирощування. Мануфактура в найбільш розвинених країнах. Перспективи розвитку Лишайники. Место обитания
Лишайники. Место обитания Деление покрытосеменных растений на классы и семейства
Деление покрытосеменных растений на классы и семейства Анатомия и физиология сердечно-сосудистой системы
Анатомия и физиология сердечно-сосудистой системы Рослинний світ прісних водойм
Рослинний світ прісних водойм Неклеточные формы жизни
Неклеточные формы жизни Природа Астраханской области
Природа Астраханской области Методы изучения природы
Методы изучения природы