- Энергетика химических реакций. Элементы химической термодинамики

Содержание
- 2. Солнечная энергия – основной источник жизни. Фотосинтез идет в автотрофных клетках. 6nCO2 + 5hH2O → (C6H10O5)n
- 3. В гетеротрофных клетках идут процессы диссимиляции АТФ + H2O → АДФ + Фн + ΔH Макроэргические
- 4. Химическая термодинамика – наука о взаимопревращениях теплоты и энергии, в том числе энергии химических процессов (середина
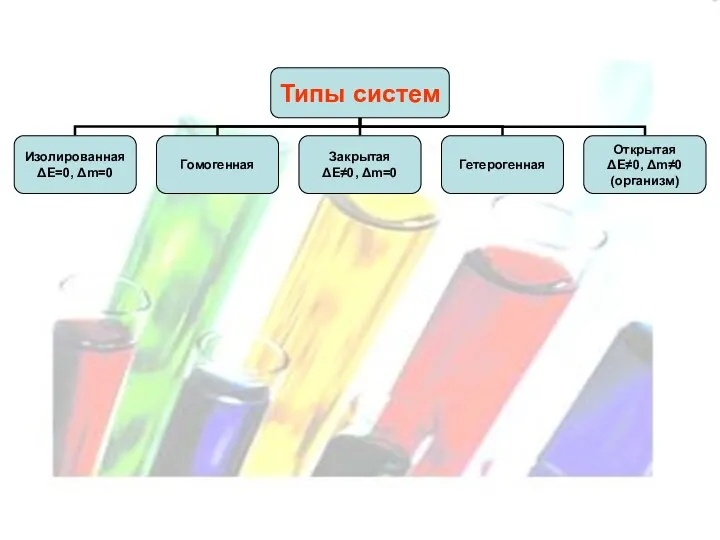
- 6. СОСТОЯНИЕ СИСТЕМЫ
- 8. Энергия – мера движения и взаимодействия материальных систем
- 9. Теплота – неупорядоченная форма передачи энергии (Q) Работа – упорядоченная форма передачи энергии (W)
- 10. I начало (закон) термодинамики (Р. Майер, Д. Джоуль, Г.Л. Гельмгольц) Это закон сохранения энергии Энергия не
- 11. Q = ΔU + W Для изобарного процесса W = pΔV Qp = ΔU + pΔV
- 12. Закон Гесса (1840 г.) (Следствие из I начала термодинамики) Конечное состояние Исходное состояние Промежуточное состояние ΔН1
- 13. Следствие из закона Гесса (следствие из I начала термодинамики) ΔНор-ии = Σm⋅ΔНообр.(прод.) - Σn⋅ΔНообр.(исх.в-в) ΔНор-ии =
- 14. Процесс – переход системы из одного состояния в другое
- 15. Новая термодинамическая функция состояния – энтропия (S). Она характеризует степень неупорядоченности материи. S = k ⋅
- 16. II начало (закон) термодинамики Невозможен самопроизвольный переход тепла от тела менее нагретого к телу более нагретому.
- 17. ∆Sобр = Qобр. / Т или в общем виде ∆S ≥ Q / T, т.е. ∆S
- 18. Д.У. Гиббс – новая функция состояния (G) – энергия Гиббса (изобарно-изотермический потенциал) ∆G = ∆H -
- 19. Для неизолированных систем самопроизвольное протекание процессов характеризуется: Энтальпийным фактором (∆Н) Энтропийным фактором (∆S)
- 20. Направленность протекания процессов
- 21. Соотношение важнейших термодинамических функций
- 22. Экзергонические реакции –∆G Эндергонические реакции –∆G > 0
- 23. Принцип энергетического сопряжения Энергия, необходимая для протекания эндергонических реакций, поступает за счет осуществления экзергонических реакций. Интермедиат
- 24. АТФ + Н2О ↔ АДФ + Н3РО4 ∆Gо = -29,2 кДж/моль Экзергоническая реакция С6Н12О6 + Н3РО4
- 25. Применение химической термодинамики к биологическим системам
- 26. Применение I начала термодинамики Термодинамические условия равновесия. Энергия Гиббса биологического окисления вещества Экзергонические и эндергонические реакции
- 28. Точность – 99% 1 г жира – 9,3 ккал 1 г углевода и белка – 4,2
- 29. Вывод: Организм не является источником энергии, а полностью подчиняется I началу термодинамики
- 30. 2-ое начало термодинамики: ΔS≥0 и ΔG≤0 Состояние равновесия: ΔS=0 и ΔG=0 Химический потенциал (Гиббс): (на 1
- 31. Может ли живой организм производить работу только за счет теплоты окружающей среды? Ответ: Прямых доказательств нет.
- 32. Играет ли увеличение энтропии определяющую роль в направлении химических процессов обмена в биосистемах? Ответ: Энтропия всей
- 33. Могут ли живые организмы использовать молекулярные отклонения в отношении уменьшения энтропии? Ответ: До сих пор известен
- 34. Применима ли к биосистемам обычная трактовка энтропии как меры упорядоченности? Ответ: Применительно к биосистемам различают: 1.
- 35. Итак, ответ на все вопросы отрицательный. Биосистема и биохимия имеют дело не с равновесными, а со
- 36. Окисление углеводов (аэробный распад) имеет большое значение в энергетическом балансе организма С6Н12О6 тв + 6О2 газ
- 37. Гликолитический путь обмена
- 39. Окислительный распад жирных кислот – второй источник (после углеводов) энергии в организме. Этот процесс локализован в
- 40. В заключительной стадии под влиянием фермента кетотиолазы образуются ацетилкоэнзим и, новая молекула коэнзима А, вновь активированная
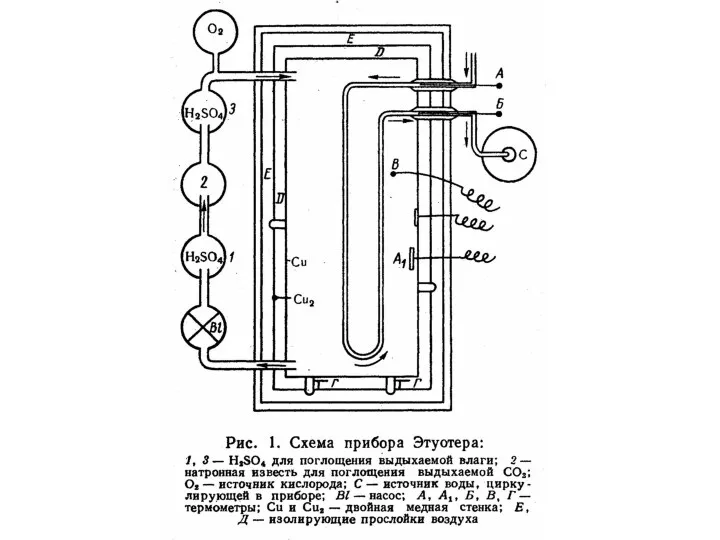
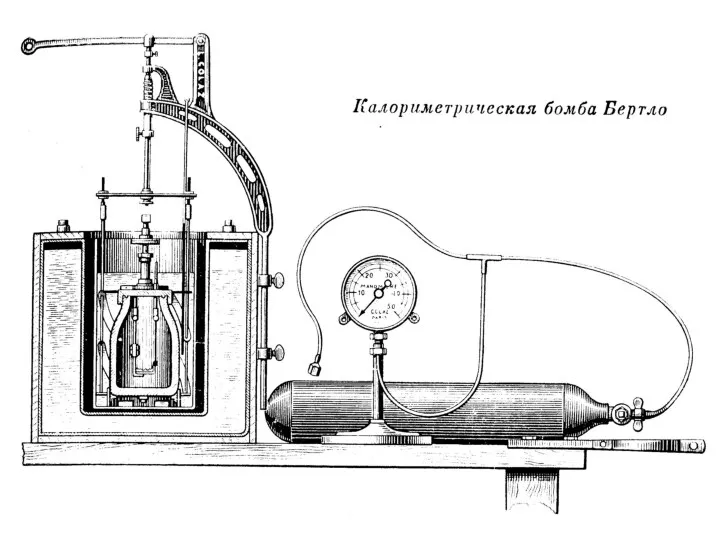
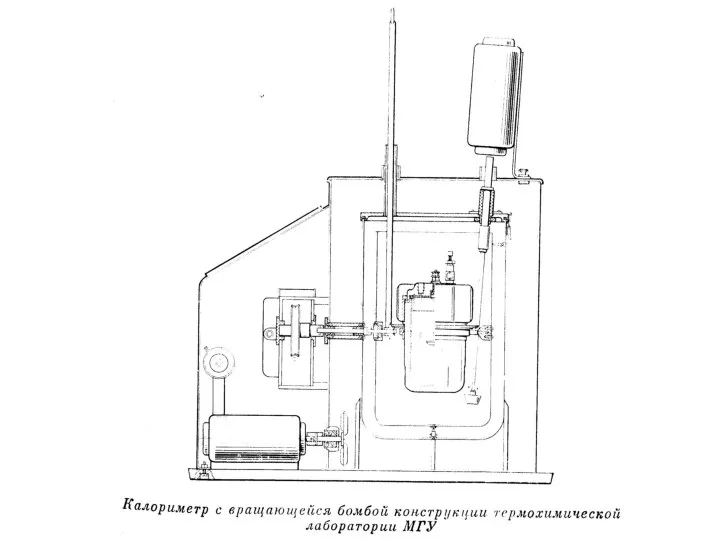
- 41. Рассмотрим образцы калориметров, используемых в разные века.
- 48. Вакуумный адиабатический калориметр
- 49. Дифференциальный сканирующий калориметр с программным управлением фирмы «СЕТАРАМ»
- 51. Скачать презентацию
Солнечная энергия – основной источник жизни.
Фотосинтез идет в автотрофных клетках.
6nCO2 +
Солнечная энергия – основной источник жизни.
Фотосинтез идет в автотрофных клетках.
6nCO2 +
В гетеротрофных клетках идут процессы диссимиляции
АТФ + H2O → АДФ +
В гетеротрофных клетках идут процессы диссимиляции
АТФ + H2O → АДФ +

Химическая термодинамика – наука о взаимопревращениях теплоты и энергии, в том
Химическая термодинамика – наука о взаимопревращениях теплоты и энергии, в том
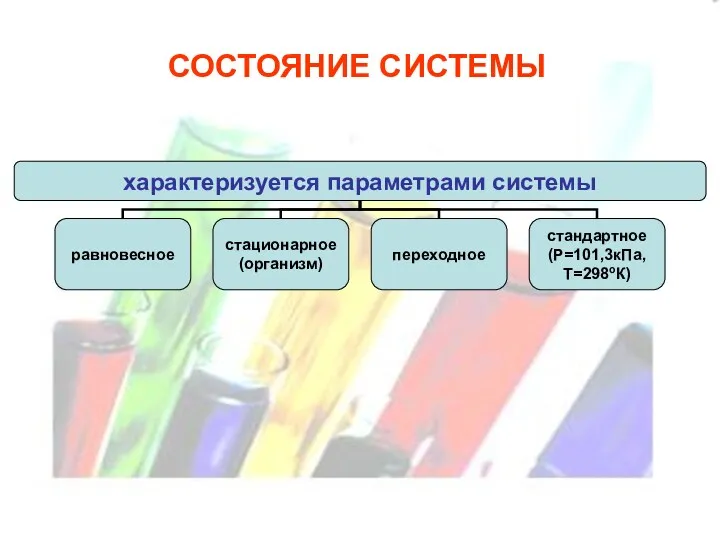
СОСТОЯНИЕ СИСТЕМЫ
СОСТОЯНИЕ СИСТЕМЫ
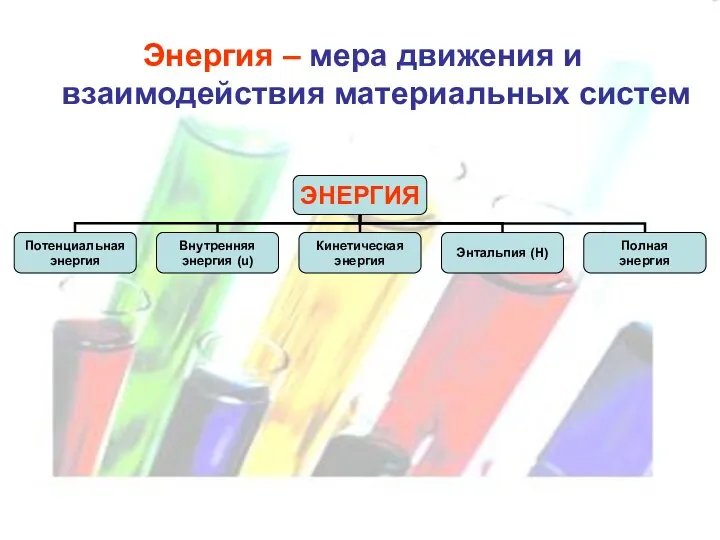
Энергия – мера движения и взаимодействия материальных систем
Энергия – мера движения и взаимодействия материальных систем
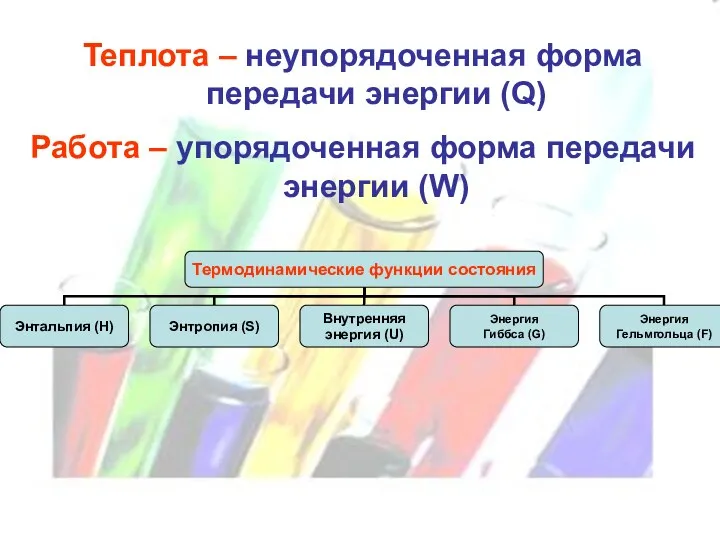
Теплота – неупорядоченная форма передачи энергии (Q)
Работа – упорядоченная форма передачи
Теплота – неупорядоченная форма передачи энергии (Q)
Работа – упорядоченная форма передачи

I начало (закон) термодинамики
(Р. Майер, Д. Джоуль, Г.Л. Гельмгольц)
Это закон сохранения
I начало (закон) термодинамики
(Р. Майер, Д. Джоуль, Г.Л. Гельмгольц)
Это закон сохранения

Q = ΔU + W
Для изобарного процесса W = pΔV
Qp =
Q = ΔU + W
Для изобарного процесса W = pΔV
Qp =
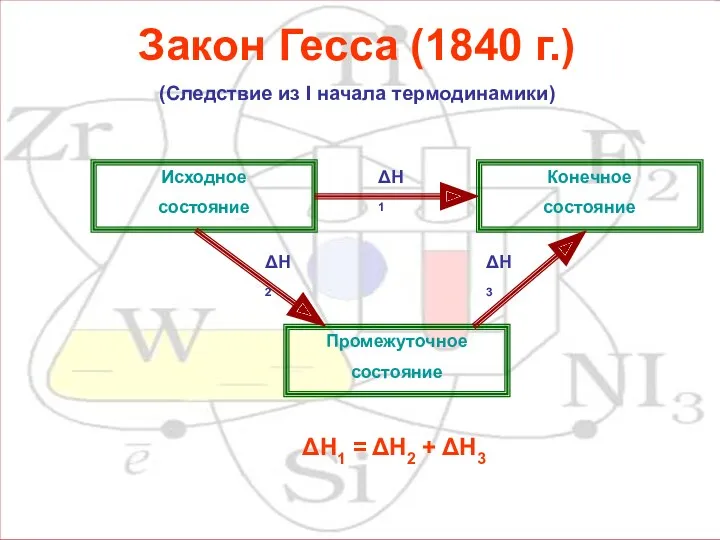
Закон Гесса (1840 г.)
(Следствие из I начала термодинамики)
Конечное
состояние
Исходное
состояние
Промежуточное
состояние
ΔН1
ΔН2
ΔН3
ΔН1
Закон Гесса (1840 г.)
(Следствие из I начала термодинамики)
Конечное
состояние
Исходное
состояние
Промежуточное
состояние
ΔН1
ΔН2
ΔН3
ΔН1

Следствие из закона Гесса
(следствие из I начала термодинамики)
ΔНор-ии = Σm⋅ΔНообр.(прод.)
Следствие из закона Гесса
(следствие из I начала термодинамики)
ΔНор-ии = Σm⋅ΔНообр.(прод.)

Процесс – переход системы из одного состояния в другое
Процесс – переход системы из одного состояния в другое
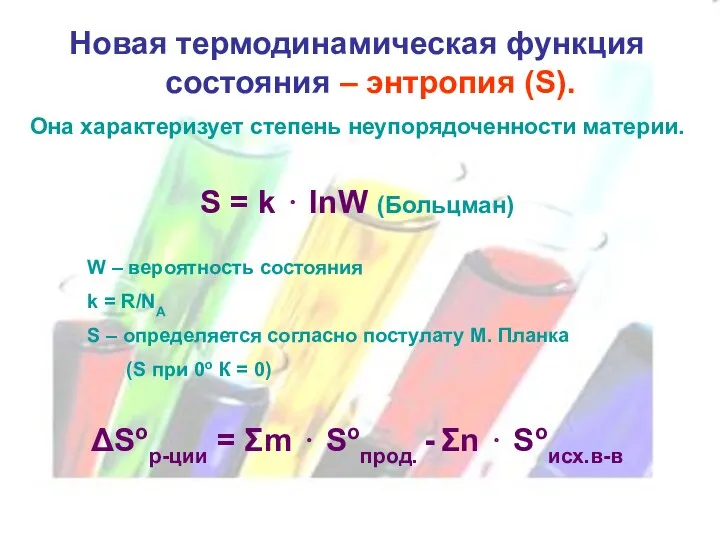
Новая термодинамическая функция состояния – энтропия (S).
Она характеризует степень неупорядоченности
Новая термодинамическая функция состояния – энтропия (S).
Она характеризует степень неупорядоченности

II начало (закон) термодинамики
Невозможен самопроизвольный переход тепла от тела менее нагретого
II начало (закон) термодинамики
Невозможен самопроизвольный переход тепла от тела менее нагретого

∆Sобр = Qобр. / Т
или в общем виде
∆S ≥ Q /
∆Sобр = Qобр. / Т
или в общем виде
∆S ≥ Q /
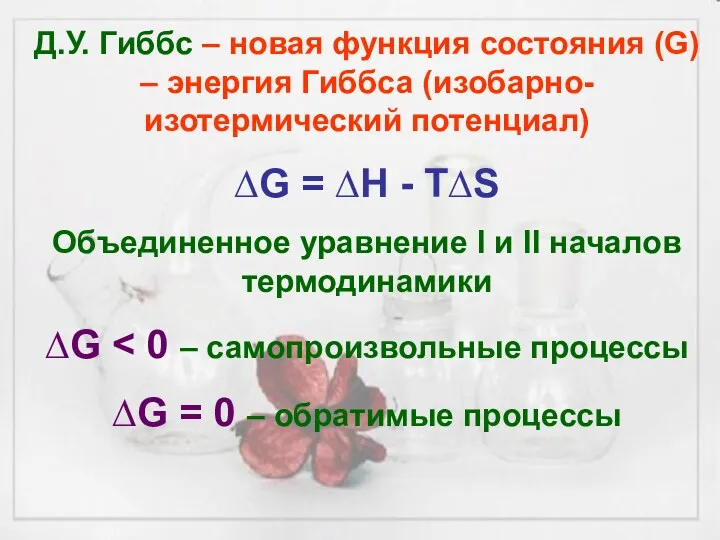
Д.У. Гиббс – новая функция состояния (G) – энергия Гиббса (изобарно-изотермический
Д.У. Гиббс – новая функция состояния (G) – энергия Гиббса (изобарно-изотермический

Для неизолированных систем самопроизвольное протекание процессов характеризуется:
Энтальпийным фактором (∆Н)
Энтропийным фактором (∆S)
Для неизолированных систем самопроизвольное протекание процессов характеризуется:
Энтальпийным фактором (∆Н)
Энтропийным фактором (∆S)

Направленность протекания процессов
Направленность протекания процессов

Соотношение важнейших термодинамических функций
Соотношение важнейших термодинамических функций

Экзергонические реакции –∆G < 0
Эндергонические реакции –∆G > 0
Экзергонические реакции –∆G < 0
Эндергонические реакции –∆G > 0

Принцип энергетического сопряжения
Энергия, необходимая для протекания эндергонических реакций, поступает за
Принцип энергетического сопряжения
Энергия, необходимая для протекания эндергонических реакций, поступает за

АТФ + Н2О ↔ АДФ + Н3РО4
∆Gо = -29,2 кДж/моль
АТФ + Н2О ↔ АДФ + Н3РО4
∆Gо = -29,2 кДж/моль

Применение химической термодинамики к биологическим системам
Применение химической термодинамики к биологическим системам

Применение I начала термодинамики
Термодинамические условия равновесия. Энергия Гиббса биологического окисления вещества
Экзергонические
Применение I начала термодинамики
Термодинамические условия равновесия. Энергия Гиббса биологического окисления вещества
Экзергонические

Точность – 99%
1 г жира – 9,3 ккал
1 г углевода и
Точность – 99%
1 г жира – 9,3 ккал
1 г углевода и

Вывод:
Организм не является источником энергии, а полностью подчиняется I началу термодинамики
Вывод:
Организм не является источником энергии, а полностью подчиняется I началу термодинамики
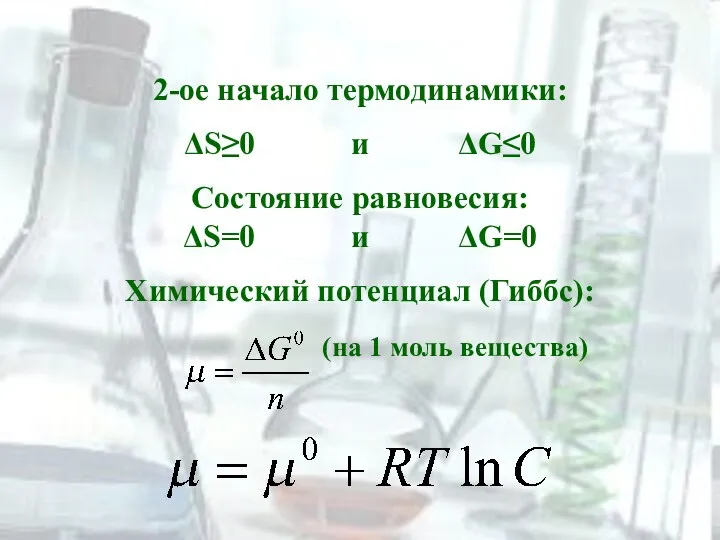
2-ое начало термодинамики:
ΔS≥0 и ΔG≤0
Состояние равновесия:
ΔS=0 и ΔG=0
Химический потенциал (Гиббс):
2-ое начало термодинамики:
ΔS≥0 и ΔG≤0
Состояние равновесия:
ΔS=0 и ΔG=0
Химический потенциал (Гиббс):

Может ли живой организм производить работу только за счет теплоты окружающей
Может ли живой организм производить работу только за счет теплоты окружающей

Играет ли увеличение энтропии определяющую роль в направлении химических процессов обмена
Играет ли увеличение энтропии определяющую роль в направлении химических процессов обмена

Могут ли живые организмы использовать молекулярные отклонения в отношении уменьшения энтропии?
Ответ:
До
Могут ли живые организмы использовать молекулярные отклонения в отношении уменьшения энтропии?
Ответ:
До

Применима ли к биосистемам обычная трактовка энтропии как меры упорядоченности?
Ответ:
Применительно к
Применима ли к биосистемам обычная трактовка энтропии как меры упорядоченности?
Ответ:
Применительно к

Итак, ответ на все вопросы отрицательный.
Биосистема и биохимия имеют дело не
Итак, ответ на все вопросы отрицательный.
Биосистема и биохимия имеют дело не

Окисление углеводов (аэробный распад) имеет большое значение в энергетическом балансе организма
С6Н12О6
Окисление углеводов (аэробный распад) имеет большое значение в энергетическом балансе организма
С6Н12О6
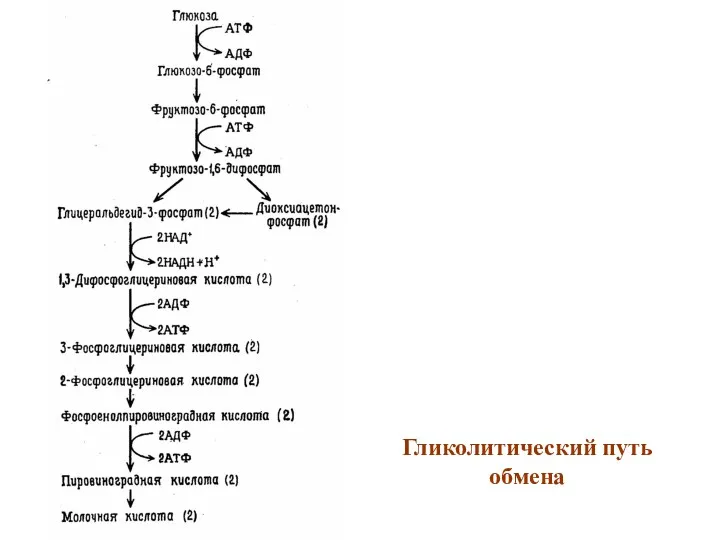
Гликолитический путь обмена
Гликолитический путь обмена


Окислительный распад жирных кислот – второй источник (после углеводов) энергии в
Окислительный распад жирных кислот – второй источник (после углеводов) энергии в

В заключительной стадии под влиянием фермента кетотиолазы образуются ацетилкоэнзим и, новая
В заключительной стадии под влиянием фермента кетотиолазы образуются ацетилкоэнзим и, новая

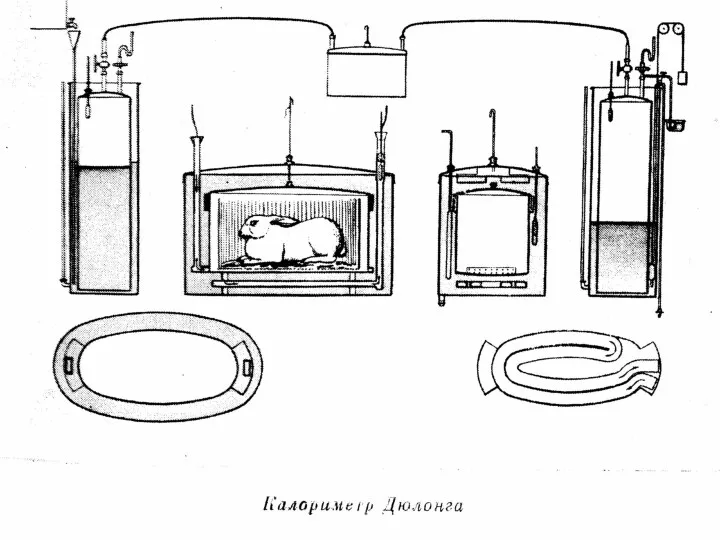
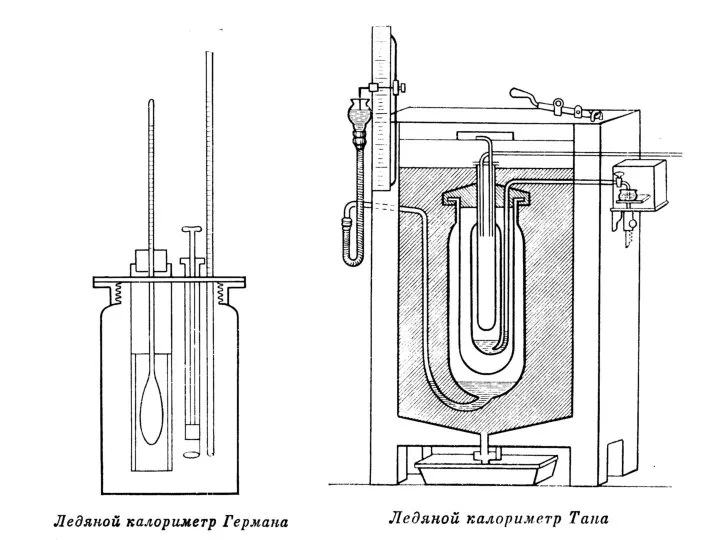
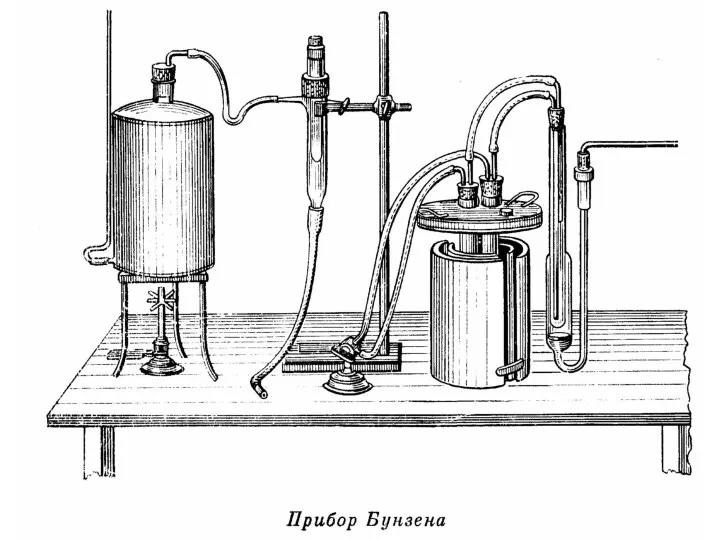
Рассмотрим образцы калориметров, используемых в разные века.
Рассмотрим образцы калориметров, используемых в разные века.

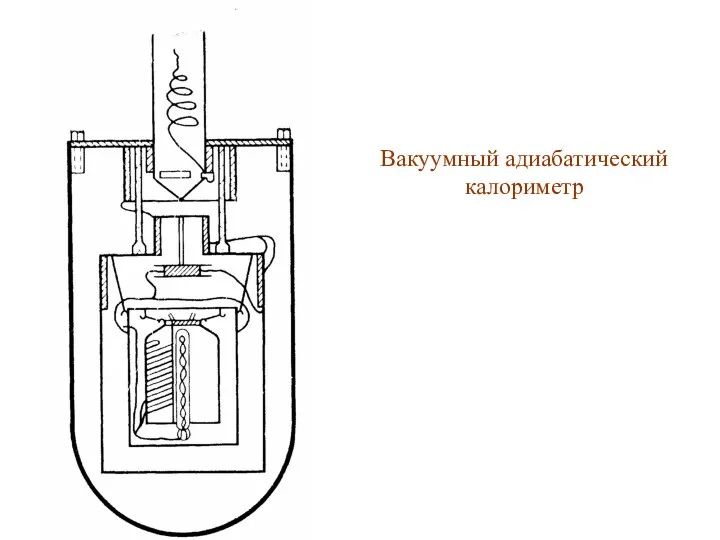
Вакуумный адиабатический калориметр
Вакуумный адиабатический калориметр

Дифференциальный сканирующий калориметр с программным управлением фирмы «СЕТАРАМ»
Дифференциальный сканирующий калориметр с программным управлением фирмы «СЕТАРАМ»
 Законы постоянного тока Тема урока: Электрический ток. Сила тока.
Законы постоянного тока Тема урока: Электрический ток. Сила тока. Електричний струм. Електрична провідність матеріалів. Дії електричного струму
Електричний струм. Електрична провідність матеріалів. Дії електричного струму Mohr circles. Equivalent stress definition and computation (1). 7 lesson
Mohr circles. Equivalent stress definition and computation (1). 7 lesson Закон Архимеда. Архимед (287 - 212 до н.э.)
Закон Архимеда. Архимед (287 - 212 до н.э.) Ультразвуковой контроль
Ультразвуковой контроль Режимы работы электродвигателей
Режимы работы электродвигателей Задачи 7 класс.
Задачи 7 класс. Урок в 8 классе Сила тока.
Урок в 8 классе Сила тока. Конвекция. Ламинарный тепловой погранслой при вынужденном движении жидкости вдоль плоской поверхности. (Тема 2. Лекции 8,9)
Конвекция. Ламинарный тепловой погранслой при вынужденном движении жидкости вдоль плоской поверхности. (Тема 2. Лекции 8,9) Обобщающий урок по теме Давление, физика, 7 класс
Обобщающий урок по теме Давление, физика, 7 класс Передача электрической энергии на расстояние
Передача электрической энергии на расстояние Свойства воды
Свойства воды к открытому уроку по теме: Простые механизмы
к открытому уроку по теме: Простые механизмы Будова атома. Закріпити знання про будову атома. Узагальнити знання про стан електронів у атомі
Будова атома. Закріпити знання про будову атома. Узагальнити знання про стан електронів у атомі Игра для профильного лагеря Умники и умницы по физике
Игра для профильного лагеря Умники и умницы по физике Классификация методов спектрального анализа и схемы его проведения
Классификация методов спектрального анализа и схемы его проведения Электростатическое поле в проводниках. Тема 4
Электростатическое поле в проводниках. Тема 4 Сучасна модель атома
Сучасна модель атома Наблюдение сплошного и линейчатых спектров (9 класс)
Наблюдение сплошного и линейчатых спектров (9 класс) презентация проблемноке обучение на уроках физики
презентация проблемноке обучение на уроках физики Лазеры
Лазеры Технология выполнения технического обслуживания и ремонта газораспределительного механизма автомобиля ЗИЛ-4333
Технология выполнения технического обслуживания и ремонта газораспределительного механизма автомобиля ЗИЛ-4333 Игра Звёздный час на тему Электрические явления
Игра Звёздный час на тему Электрические явления Формирование ключевых компетенций при решении творческих задач на уроках физики
Формирование ключевых компетенций при решении творческих задач на уроках физики Прилади акустоелектроніки. Резонатори. Хвилеводи. Лінії затримки
Прилади акустоелектроніки. Резонатори. Хвилеводи. Лінії затримки Техническая механика. Связи и их реакции
Техническая механика. Связи и их реакции Электростатическое поле в вакууме
Электростатическое поле в вакууме Магнитные цепи при постоянном магнитном потоке
Магнитные цепи при постоянном магнитном потоке