Белки групп Trithorax и Polycomb (TrxG/PcG) - ключевые эпигенетические факторы поддержания клеточной дифференцировки презентация
- Белки групп Trithorax и Polycomb (TrxG/PcG) - ключевые эпигенетические факторы поддержания клеточной дифференцировки

Содержание
- 2. МОДЕЛЬ ФОРМИРОВАНИЯ ПЕРЕДНЕ-ЗАДНИХ ГРАДИЕНТОВ Bcd Hb Как контролируется экспрессия Hox-генов на поздних стадиях развития?
- 3. Мутант по гену Polycomb Половой гребешок (sex comb) Дикий тип Мутант 1 нога 2 нога 3
- 4. Мутант по гену из семейства Polycomb -Suppressor of zeste 12 Мутация в не гомеозисном гене может
- 5. Мутации другой группы генов TRX-C приводит к репрессии гомеозисных генов mor kis Репрессия Ubx Репрессия AbdB
- 6. Активация группы генов TRX-C приводит к дерепрессии гомеозисных генов По: SadasivamSadasivam,Sadasivam, Huang 2016. PLoS Genet
- 7. Реактивация гомеозисных генов усиливается на фоне мутации генов Hira и dom, кодирующих гистоновые шапероны Эктопическая экспрессия
- 8. PcG и TrxG - поддержание экспрессии генов Ранее развитие Детерминация, инициация Mатеринские гены, Gap, Pair-rule, segment
- 9. ДНК ДНК Polyhomeotic
- 10. Связывание необходимо для поддержания репрессии (или усиления уровня экспрессии) репортерных генов Связаны с PcG/TrxG-белками in vivo
- 11. w w w «Pairing»-чувствительная репрессия: усиление репрессии репортерного гена mini-white при увеличении копий PRE (Fab-7) --->
- 12. P P В трансгенной линии Fab-X, элемент PRE (Fab-7 ) в гомозиготном состоянии репрессирует ген, расположенный
- 13. Взаимодействие между гомологичными копиями PREs
- 14. Fab-7 0 5 10 15 20 25 + + - Endogenous Fab-7 at the BX-C (Chr.
- 15. Два идентичных Fab-7, находящиеся на различных хромосомах (!) могут взаимодействовать в ядре, что ведет к усилению
- 16. Филогенетическое сравнение сайтов PRE/TRE района bxd гена Ubx
- 17. Процессы, контролируемые PcG и TrxG
- 18. Исходно открытые у Drosophila как регуляторы гомейозисных генов HOX, отвечают за спецификацию плана формирования тела, а
- 19. Что такое эпигенетическая регуляция?
- 20. На уровне базовой транскрипции (базовые (TFII) и специфические транскрипционные факторы) На уровне компактизации хроматина (факторы, ремодулирующие
- 21. Схема преинициаторного транскрипционного комплекса (Регуляция на уровне транскрипции) (TAFIIs) TBP - главный TFII (TFII) РНК-полимераза II
- 22. Белки групп PcG и TrxG участвуют в «ремоделировании» хроматина, изменяя доступность ДНК для других факторов, требующихся
- 23. Где на хромосомах находится «закрытый» и «открытый» хроматин? «Закрытый» конденсированный хроматин «Открытый» хроматин Конститутивный гетерохроматин Теломеры
- 24. Нуклеосома СХЕМА УПАКОВКИ ХРОМАТИНА Нуклеомера Хромонема Хроматида Хромомера 1. ДНК 2.Нуклеосома 3. Нуклеомера 4. Хромомера 5.
- 25. Хроматин является не просто пассивным «упаковщиком» ДНК, но и носителем эпигенетической информации. Механизмы, кодирующие эпигенетическую информацию:
- 26. Метилирование ДНК и связанные с ним процессы
- 27. Молекулярные основы эпигенетики Метилирование ДНК Б.Ф. Ванюшин Robin Holliday Впервые определил природу метилируемых последовательностей ДНК у
- 28. Метилирование ДНК Метилированию в норме подвергается от 2 до 7% всех цитозиновых остатков ДНК клетки. При
- 29. Высокометилированые последовательности: Сателлитная ДНК Повторяющиеся элементы (в т.ч. транспозоны) Уникальная межгенная ДНК Экзоны генов De novo
- 30. Механизмы инактивации гена в результате метилирования промоторной области 1. Метильные группы нарушают ДНК-белковые взаимодействия, выступая в
- 31. CpG – островки неметилированные участки длиной 1 kb - в 5`-концах 60% промоторов активных генов Что
- 32. Репрессия транскрипции посредством метилирования ДНК
- 33. Варианты эпигенетической регуляции с участием метилирования ДНК
- 34. Геномный импринтинг — эпигенетический процесс, при котором экспрессия определенных генов осуществляется в зависимости от того, от
- 35. Примеры импринтинга
- 36. Примеры импринтинга Лигр – папа лев, мама тигрица Тигрон – папа тигр мама лев
- 37. Неравнозначность пронуклеусов Мужской и женский пронуклеусы можно считать генетически эквивалентными, однако результаты проведённых исследований свидетельствуют о
- 38. Эксперименты по трансплантации пронуклеусов у мышей (или почему невозможен партеногенез у млекопитающих)
- 39. Зачем нужны такие гены???
- 40. Импринтинг генов в составе генома показан только для млекопитающих кроме яйцекладущих. Импринтинг – редкий процесс –
- 41. Оба родителя стремятся увеличить шансы на эволюционный успех своих генов за счет ресурсов только одного из
- 42. Портрет Евгении Мартинес Валеджо 1680. Музей Прадо, Мадрид. Считается, что девочка страдала синдромом Прадера-Вилли. На картине
- 43. Общая схема структуры кластера импринтированных генов Белок кодирующий ген Некодирующая РНК Район, контролирующий импринтинг (ICR) Отцовский
- 44. Схема работы импринтированного локуса инсулиноподобного фактора роста 2 Igf2/H19 Дифференциально метилированный район
- 45. Посттрансляционные модификации гистонов «Гистоновый код»
- 46. Структура нуклеосомы Ядро нуклеосомы состоит из гистонов четырех типов (по два каждого) H2A, H2B, H3 и
- 47. Нуклеосомы - рентгеноструктурный анализ в двух проекциях
- 48. Молекула каждого гистона состоит из структурированного центрального домена (альфа-спиральные участки, соединенные петельным сегментами) и неструктурированного N-терминального
- 50. «Гистоновый код» – совокупность ковалентных модификаций гистонов
- 52. Фиолетовый - модификации, характерные для репрессированного хроматина, красный – для активного хроматина. Серый - модификации, связанные
- 53. Ацетилирование /деацетилирование гистонов и ремоделирование хроматина Ацетилирование связано с активацией транскрипции белки, осуществляющие ацетилирование - гистоновые
- 54. Домены белков, распознающие модификацию - метилированные лизины, ацетилированные лизины, фосфорилированные серины - принадлежат белкам PcG и
- 55. Как формируются и функционируют комплексы PcG и TxG?
- 56. PcG-белки выявляются в составе многокомпонентных комплексов 1 комплекс, Polycomb Repressive Complex 1 (PRC1), содержит белки Polycomb
- 57. 1. Связывание комплекса PhoRC 2. Привлечение комплекса PRC2 через взаимодействие с PhoRC 3. PRC2-зависимая модификация гистонов
- 58. Комплексы PcG у дрозофилы Bantignies and Cavalli (2011) Trends in Genetics 1 2 5 4 3
- 59. Разнообразие комплексов PcG. (из Schuettengruber and Cavalli, 2009). (A) Три принципиальных типа комплексов белков группы Поликомба
- 60. Комплексы и белки группы TrxG у разных видов
- 61. Нуклеосом ремоделирующие комплексы белков TrxG содержат субъединицы с АТФазной активностью АТФазный домен в этих субъединицах разделён
- 62. Эволюционное разнообразие SWI/SNF-комплексов (из [Ho and Crabtree, 2010]). Гомологичные субъединицы комплексов дрожжей Saccharomyces cerevisiae (Yeast Swi/Snf),
- 63. Комплексы и белки группы TrxG у разных видов Гистон-модифицирующие комплексы Ацетилтрансфераза гистона (HAT)
- 64. Комплексы и белки группы TrxG у разных видов АТФ-зависимые ремоделирующие комплексы АТФаза Brahma
- 65. Из обзора Schuettengruber et al. (2011) Nature, 12: 799-814 Как рекртируются комплексы Trx-G?
- 66. Суммарный механизм функционирования модуля клеточной памяти (CMM) CBP CMM dRing PH PC PRC1/other PcG related complexes
- 67. Если PRE детерминируется во время раннего развития… dRing PH PC PSC dRing PH PC PSC CMM
- 68. … и когда функция TRE активируется CMM PRE TRE
- 69. Как модули «клеточной памяти» передают информацию дочерним клеткам ? Или Как стабильно наследуется потенциально обратимые модификации
- 70. Гипотетический механизм воспроизведения (сохранения) модификаций гистонов и связанных с ними белков на дочерних цепях ДНК после
- 71. TrxG and PcG Proteins but Not Methylated Histones Remain Associated with DNA through Replication Petruk et
- 72. TrxG and PcG Proteins but Not Methylated Histones Remain Associated with DNA through Replication Petruk et
- 73. The proliferating cell nuclear antigen (PCNA) protein acts as a platform to recruit DNA polymerase and
- 74. Взаимосвязь механизмов эпигенетической регуляции Процессы модификации ДНК и хроматина отражают разные стороны одного механизма. В частности,
- 75. Метилирование ДНК активирует модификации гистонов DAMT – ДНК метилтрансфераза Гистоновая метил трансфераза HMT
- 76. Мир некодирующих РНК (нкРНК) Недавние исследования показали, что большая часть генома человека транскрибируется как нкРНК Белок-кодирующая
- 77. Эпигенетическая регуляция: Антисмысловые транскрипты формирует структуру, которая узнаётся различными гистон-модифицирующими комплексами. Рекрутируемые комплексы могут действовать, как
- 78. Пример эпигенетической регуляции в цис-положении
- 79. Current Opinion in Genetics & Development 2012, 22:172–178 upstream-antisense RNAs enhancer-templated RNAs RNA Marks Enhancer Elements:
- 81. Скачать презентацию
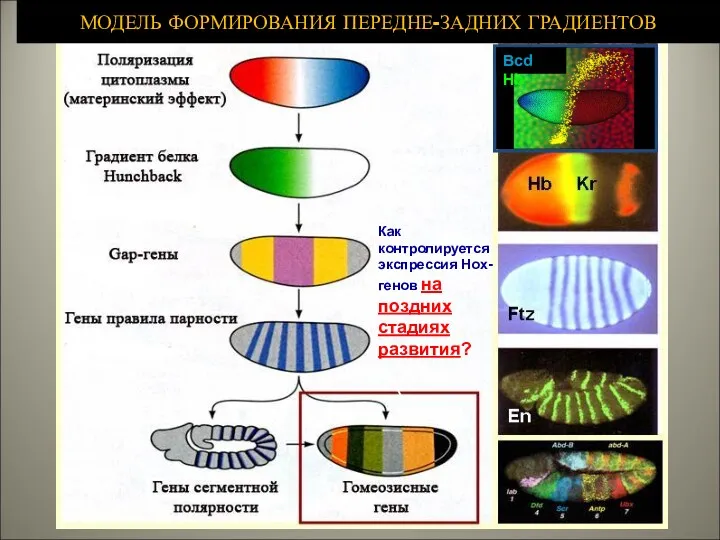
МОДЕЛЬ ФОРМИРОВАНИЯ ПЕРЕДНЕ-ЗАДНИХ ГРАДИЕНТОВ
Bcd Hb
Как контролируется экспрессия Hox-генов на поздних стадиях
МОДЕЛЬ ФОРМИРОВАНИЯ ПЕРЕДНЕ-ЗАДНИХ ГРАДИЕНТОВ
Bcd Hb
Как контролируется экспрессия Hox-генов на поздних стадиях

Мутант по гену Polycomb
Половой гребешок
(sex comb)
Дикий тип
Мутант
1 нога 2 нога 3
Мутант по гену Polycomb
Половой гребешок
(sex comb)
Дикий тип
Мутант
1 нога 2 нога 3

Мутант по гену из семейства Polycomb -Suppressor of zeste 12
Мутация в
Мутант по гену из семейства Polycomb -Suppressor of zeste 12
Мутация в
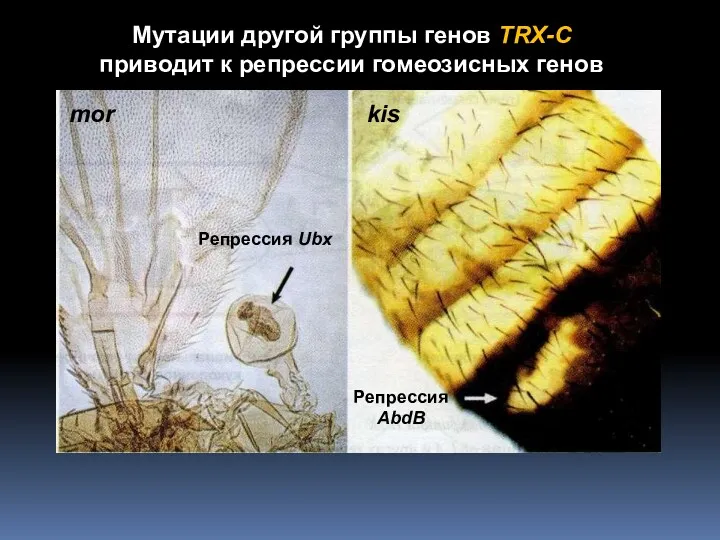
Мутации другой группы генов TRX-C
приводит к репрессии гомеозисных генов
mor
kis
Репрессия Ubx
Репрессия
Мутации другой группы генов TRX-C
приводит к репрессии гомеозисных генов
mor
kis
Репрессия Ubx
Репрессия
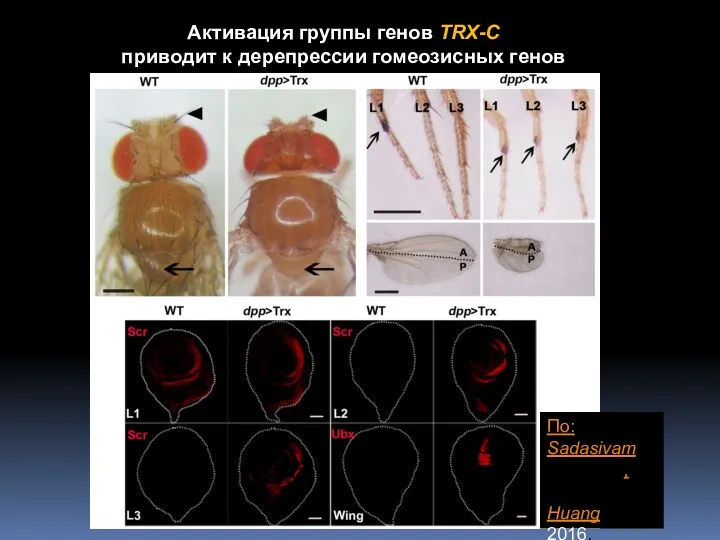
Активация группы генов TRX-C
приводит к дерепрессии гомеозисных генов
По: SadasivamSadasivam,Sadasivam, Huang
Активация группы генов TRX-C
приводит к дерепрессии гомеозисных генов
По: SadasivamSadasivam,Sadasivam, Huang
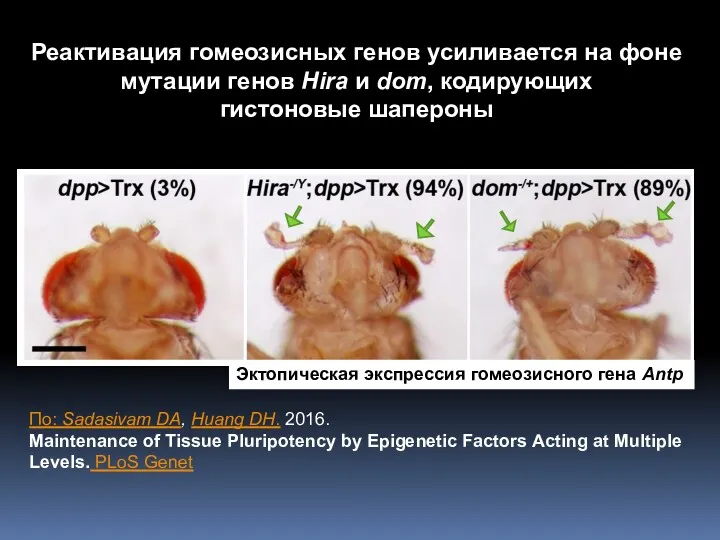
Реактивация гомеозисных генов усиливается на фоне
мутации генов Hira и dom,
Реактивация гомеозисных генов усиливается на фоне
мутации генов Hira и dom,
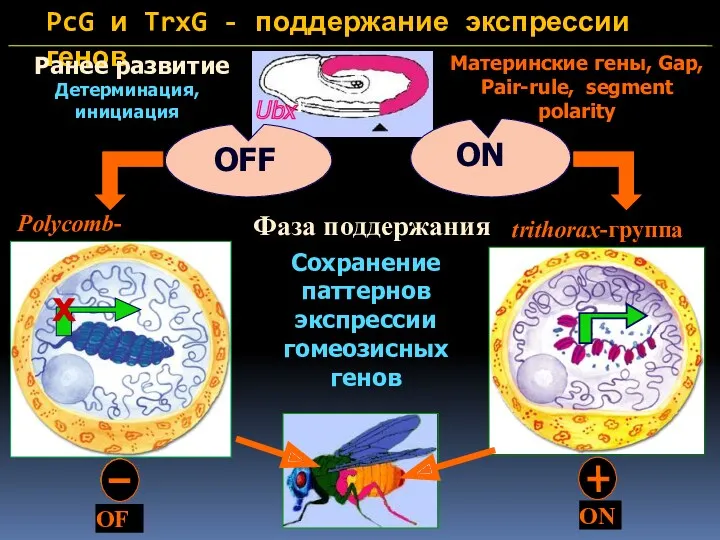
PcG и TrxG - поддержание экспрессии генов
Ранее развитие
Детерминация,
инициация
Mатеринские
PcG и TrxG - поддержание экспрессии генов
Ранее развитие
Детерминация,
инициация
Mатеринские
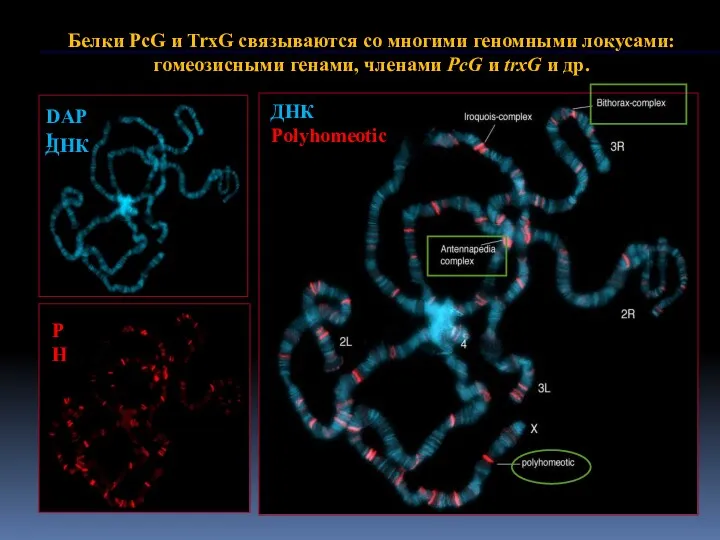
ДНК
ДНК
Polyhomeotic
ДНК
ДНК
Polyhomeotic

Связывание необходимо для поддержания репрессии (или усиления уровня экспрессии) репортерных
Связывание необходимо для поддержания репрессии (или усиления уровня экспрессии) репортерных
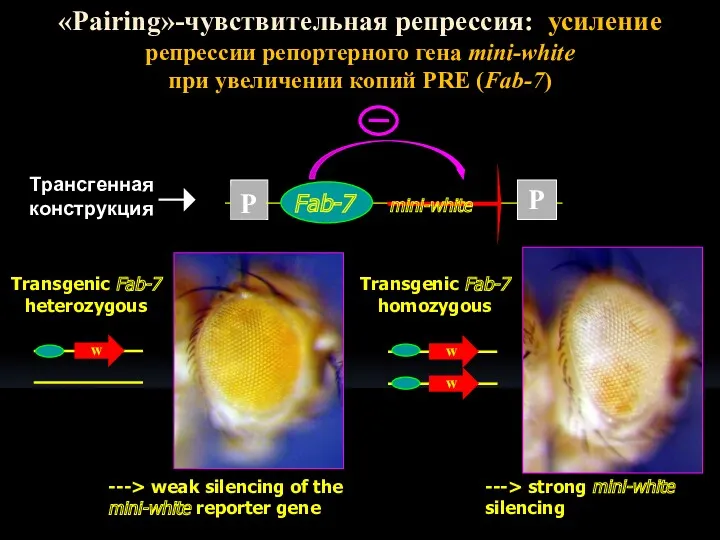
w
w
w
«Pairing»-чувствительная репрессия: усиление репрессии репортерного гена mini-white
при увеличении копий PRE
w
w
w
«Pairing»-чувствительная репрессия: усиление репрессии репортерного гена mini-white
при увеличении копий PRE
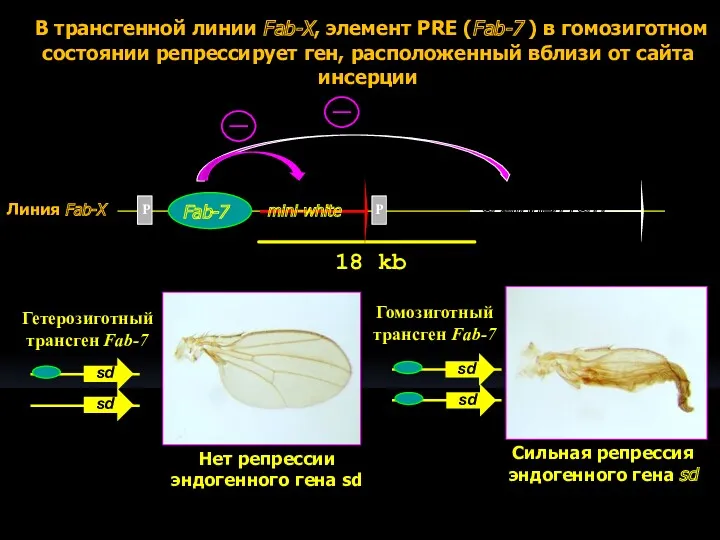
P
P
В трансгенной линии Fab-X, элемент PRE (Fab-7 ) в гомозиготном
P
P
В трансгенной линии Fab-X, элемент PRE (Fab-7 ) в гомозиготном

Взаимодействие между гомологичными копиями PREs
Взаимодействие между гомологичными копиями PREs
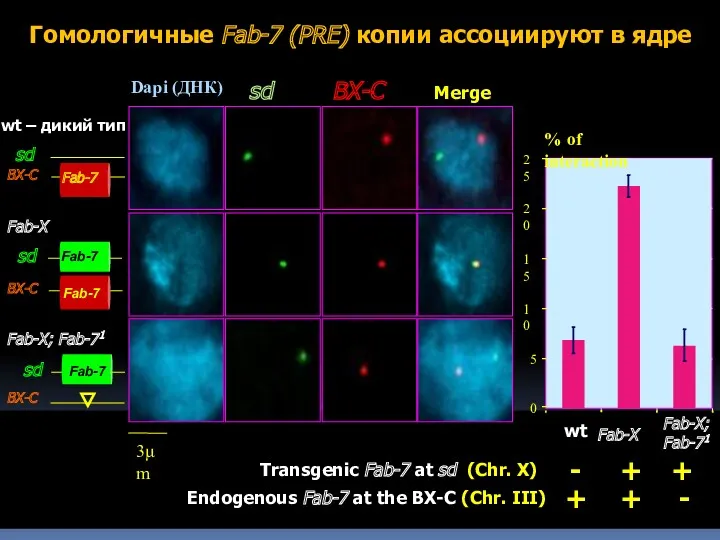
Fab-7
0
5
10
15
20
25
+
+
-
Endogenous Fab-7 at the BX-C (Chr. III)
3μm
-
-
-
-
-
-
Fab-X
Fab-X;
Fab-71
wt
wt – дикий тип
Fab-X
Fab-X;
Fab-7
0
5
10
15
20
25
+
+
-
Endogenous Fab-7 at the BX-C (Chr. III)
3μm
-
-
-
-
-
-
Fab-X
Fab-X;
Fab-71
wt
wt – дикий тип
Fab-X
Fab-X;
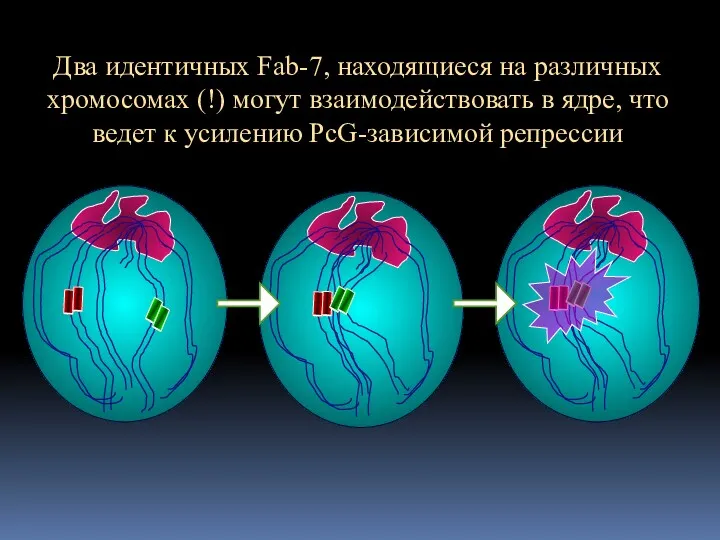
Два идентичных Fab-7, находящиеся на различных хромосомах (!) могут взаимодействовать в
Два идентичных Fab-7, находящиеся на различных хромосомах (!) могут взаимодействовать в

Филогенетическое сравнение сайтов PRE/TRE
района bxd гена Ubx
Филогенетическое сравнение сайтов PRE/TRE
района bxd гена Ubx
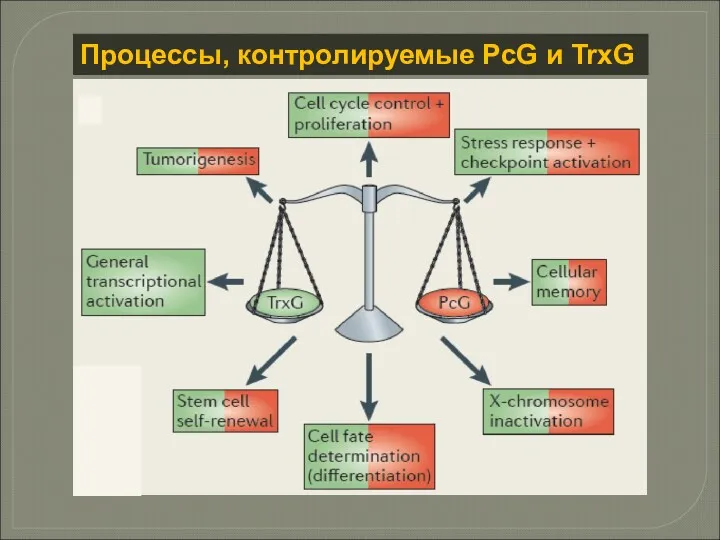
Процессы, контролируемые PcG и TrxG
Процессы, контролируемые PcG и TrxG

Исходно открытые у Drosophila как регуляторы гомейозисных генов HOX, отвечают за

Что такое эпигенетическая регуляция?
Что такое эпигенетическая регуляция?

На уровне базовой транскрипции (базовые (TFII) и специфические транскрипционные
На уровне базовой транскрипции (базовые (TFII) и специфические транскрипционные
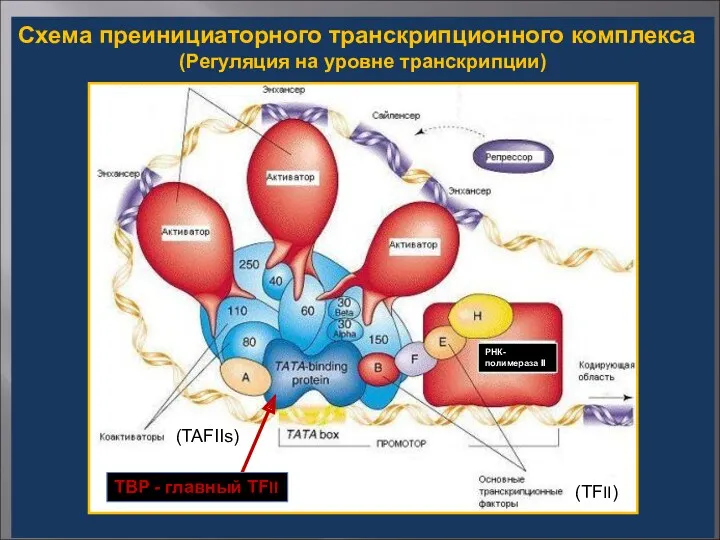
Схема преинициаторного транскрипционного комплекса
(Регуляция на уровне транскрипции)
(TAFIIs)
TBP - главный TFII
(TFII)
РНК-полимераза
Схема преинициаторного транскрипционного комплекса
(Регуляция на уровне транскрипции)
(TAFIIs)
TBP - главный TFII
(TFII)
РНК-полимераза

Белки групп PcG и TrxG участвуют в «ремоделировании» хроматина, изменяя доступность
Белки групп PcG и TrxG участвуют в «ремоделировании» хроматина, изменяя доступность
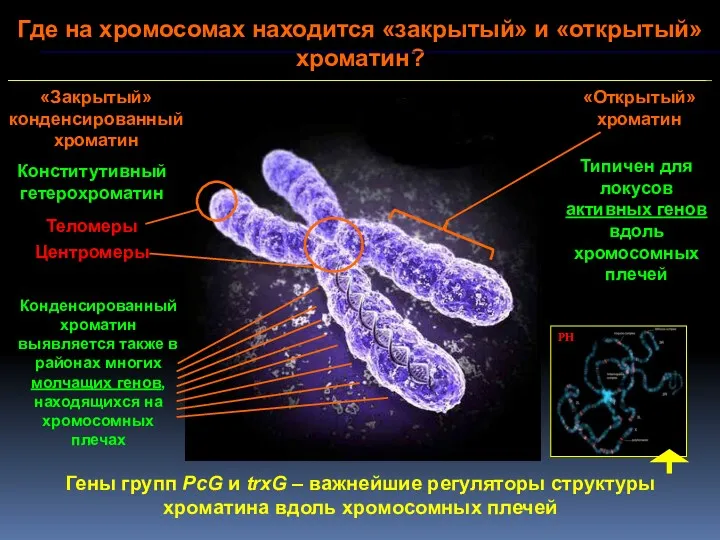
Где на хромосомах находится «закрытый» и «открытый» хроматин?
«Закрытый» конденсированный хроматин
«Открытый»
Где на хромосомах находится «закрытый» и «открытый» хроматин?
«Закрытый» конденсированный хроматин
«Открытый»
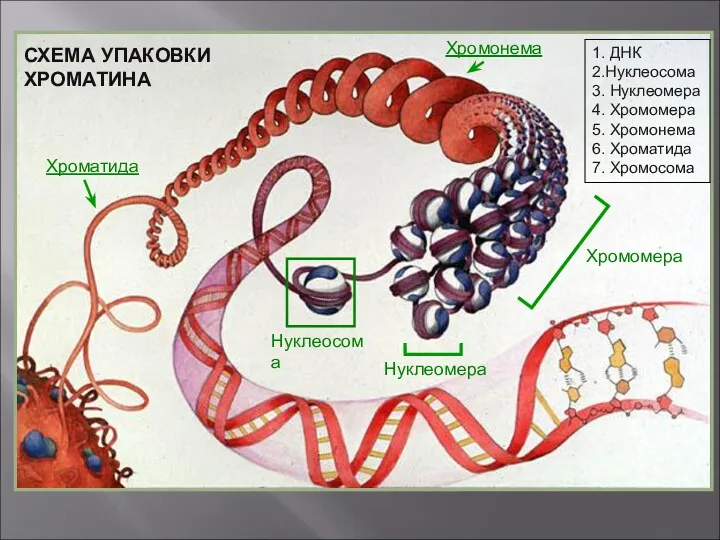
Нуклеосома
СХЕМА УПАКОВКИ ХРОМАТИНА
Нуклеомера
Хромонема
Хроматида
Хромомера
1. ДНК
2.Нуклеосома
3. Нуклеомера
4. Хромомера
5. Хромонема
6. Хроматида
7. Хромосома
Нуклеосома
СХЕМА УПАКОВКИ ХРОМАТИНА
Нуклеомера
Хромонема
Хроматида
Хромомера
1. ДНК
2.Нуклеосома
3. Нуклеомера
4. Хромомера
5. Хромонема
6. Хроматида
7. Хромосома

Хроматин является не просто пассивным «упаковщиком» ДНК, но и носителем эпигенетической
Хроматин является не просто пассивным «упаковщиком» ДНК, но и носителем эпигенетической
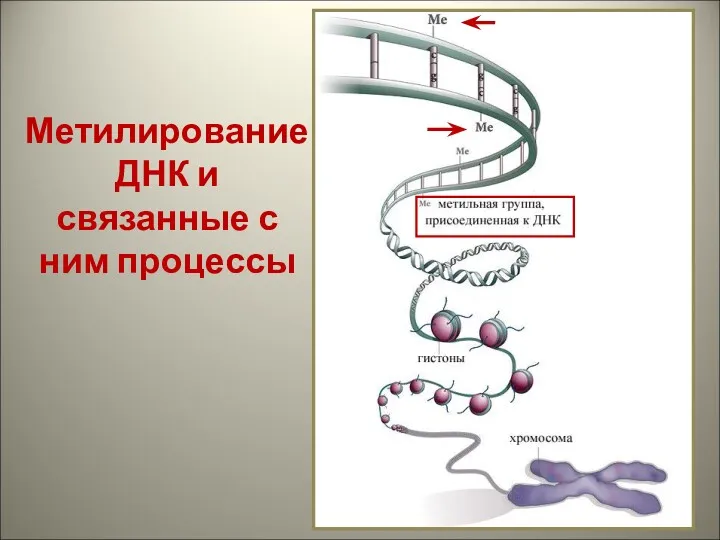
Метилирование ДНК и связанные с ним процессы
Метилирование ДНК и связанные с ним процессы

Молекулярные основы эпигенетики
Метилирование ДНК
Б.Ф. Ванюшин
Robin Holliday
Впервые определил природу метилируемых последовательностей ДНК
Молекулярные основы эпигенетики
Метилирование ДНК
Б.Ф. Ванюшин
Robin Holliday
Впервые определил природу метилируемых последовательностей ДНК
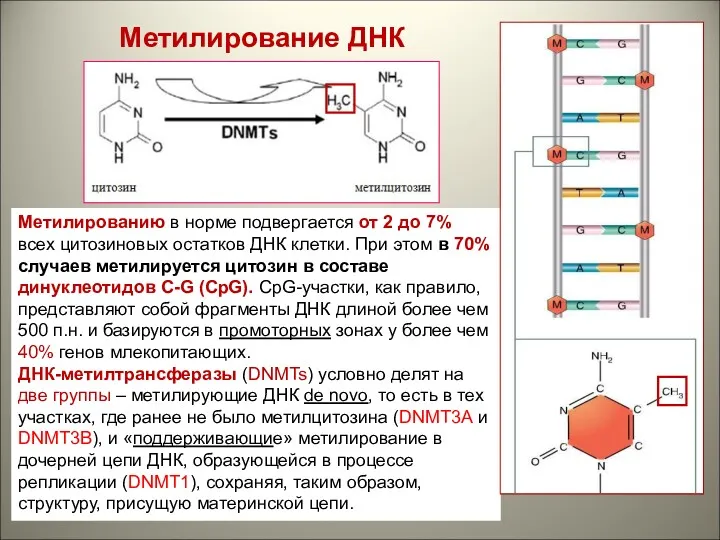
Метилирование ДНК
Метилированию в норме подвергается от 2 до 7%
Метилирование ДНК
Метилированию в норме подвергается от 2 до 7%
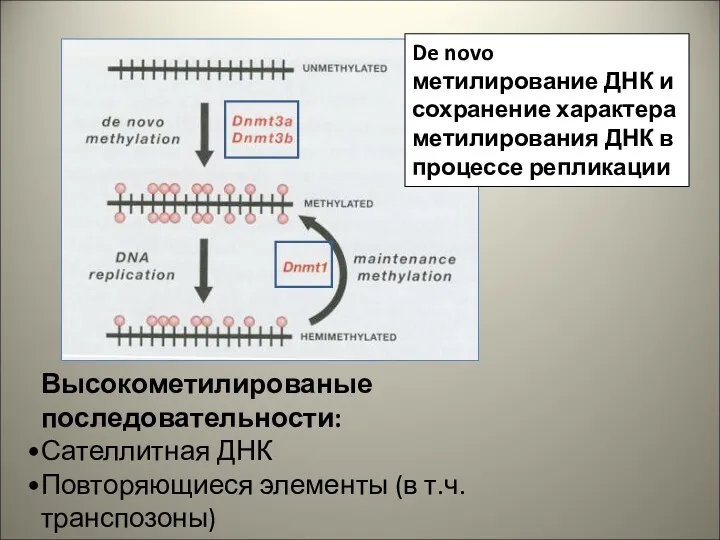
Высокометилированые последовательности:
Сателлитная ДНК
Повторяющиеся элементы (в т.ч. транспозоны)
Уникальная межгенная ДНК
Экзоны генов
De novo
Высокометилированые последовательности:
Сателлитная ДНК
Повторяющиеся элементы (в т.ч. транспозоны)
Уникальная межгенная ДНК
Экзоны генов
De novo

Механизмы инактивации гена в результате метилирования промоторной области
1. Метильные группы
Механизмы инактивации гена в результате метилирования промоторной области
1. Метильные группы
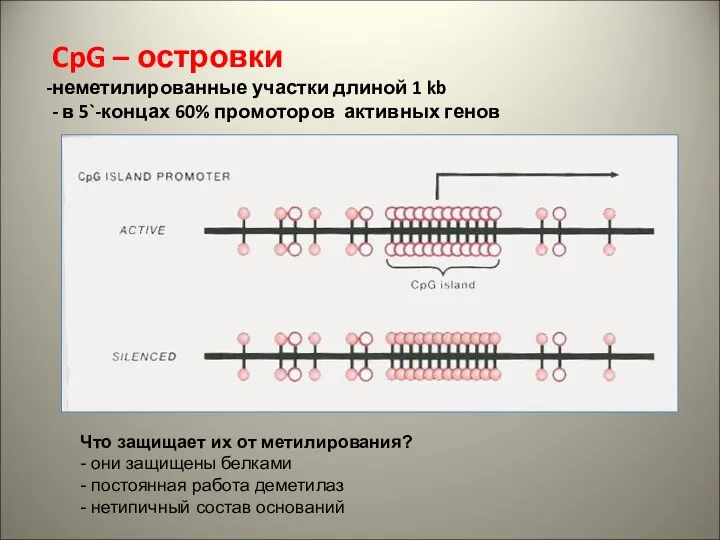
CpG – островки
неметилированные участки длиной 1 kb
- в 5`-концах 60%
CpG – островки
неметилированные участки длиной 1 kb
- в 5`-концах 60%
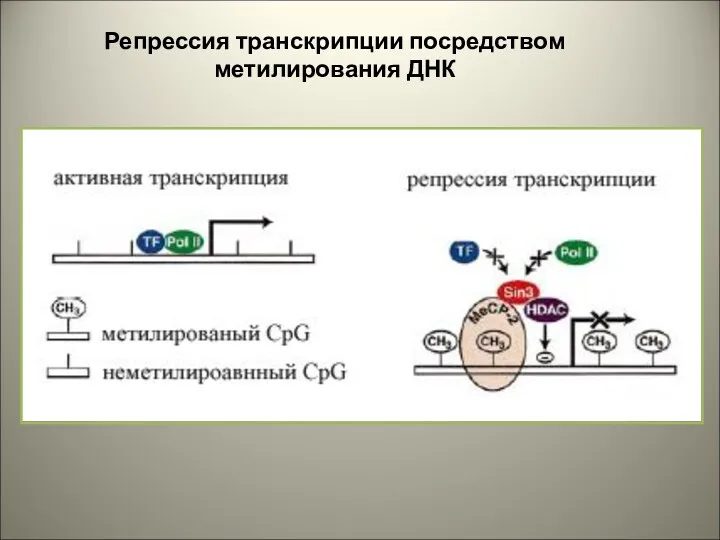
Репрессия транскрипции посредством метилирования ДНК
Репрессия транскрипции посредством метилирования ДНК

Варианты эпигенетической регуляции с участием метилирования ДНК
Варианты эпигенетической регуляции с участием метилирования ДНК

Геномный импринтинг — эпигенетический процесс, при котором экспрессия определенных генов осуществляется
Геномный импринтинг — эпигенетический процесс, при котором экспрессия определенных генов осуществляется

Примеры импринтинга
Примеры импринтинга

Примеры импринтинга
Лигр –
папа лев,
мама тигрица
Тигрон –
папа тигр
мама лев
Примеры импринтинга
Лигр –
папа лев,
мама тигрица
Тигрон –
папа тигр
мама лев
Неравнозначность пронуклеусов
Мужской и женский пронуклеусы можно считать генетически эквивалентными, однако
Неравнозначность пронуклеусов
Мужской и женский пронуклеусы можно считать генетически эквивалентными, однако
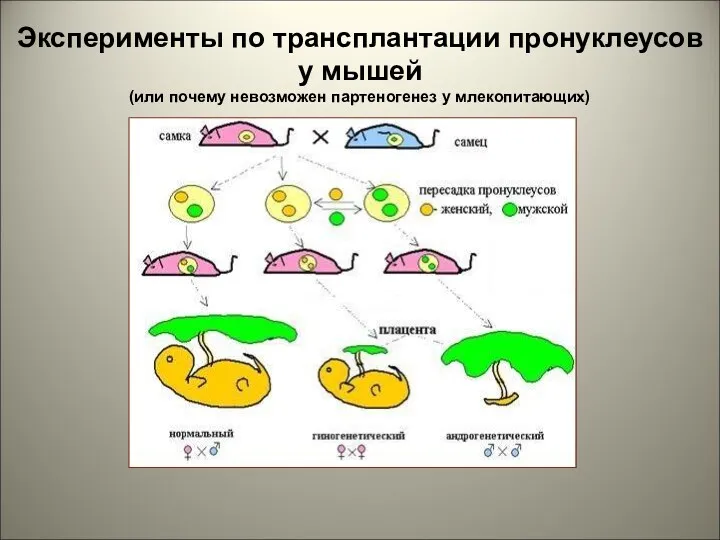
Эксперименты по трансплантации пронуклеусов у мышей
(или почему невозможен партеногенез у
Эксперименты по трансплантации пронуклеусов у мышей
(или почему невозможен партеногенез у
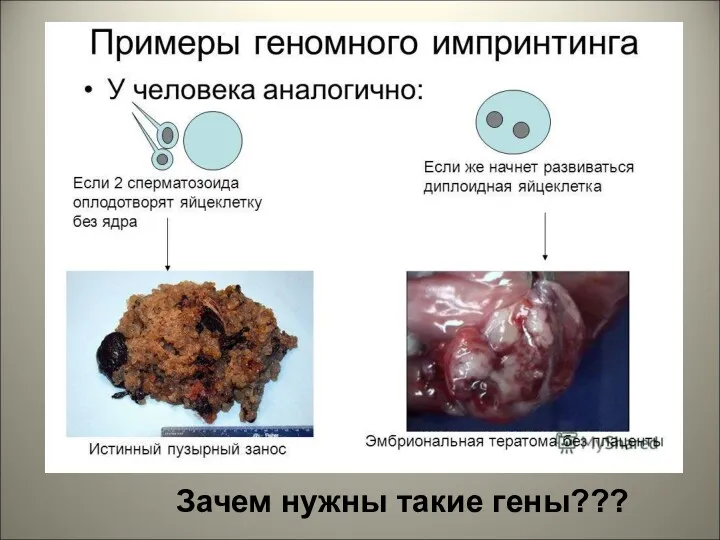
Зачем нужны такие гены???
Зачем нужны такие гены???
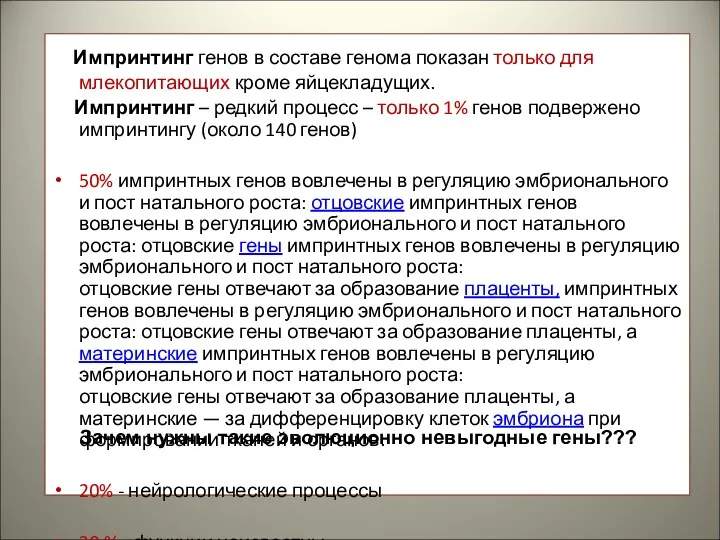
Импринтинг генов в составе генома показан только для млекопитающих кроме
Импринтинг генов в составе генома показан только для млекопитающих кроме

Оба родителя стремятся увеличить шансы на эволюционный успех своих генов за

Портрет
Евгении Мартинес Валеджо 1680. Музей Прадо, Мадрид.
Считается, что девочка
Портрет
Евгении Мартинес Валеджо 1680. Музей Прадо, Мадрид.
Считается, что девочка
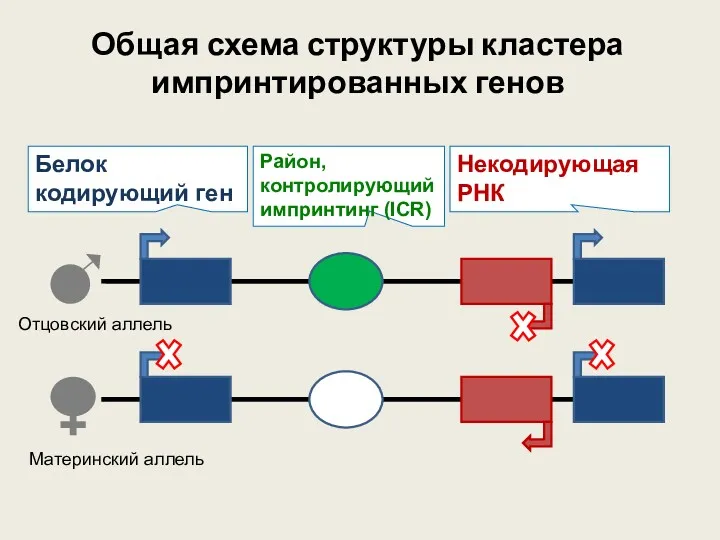
Общая схема структуры кластера импринтированных генов
Белок кодирующий ген
Некодирующая РНК
Район, контролирующий импринтинг
Общая схема структуры кластера импринтированных генов
Белок кодирующий ген
Некодирующая РНК
Район, контролирующий импринтинг
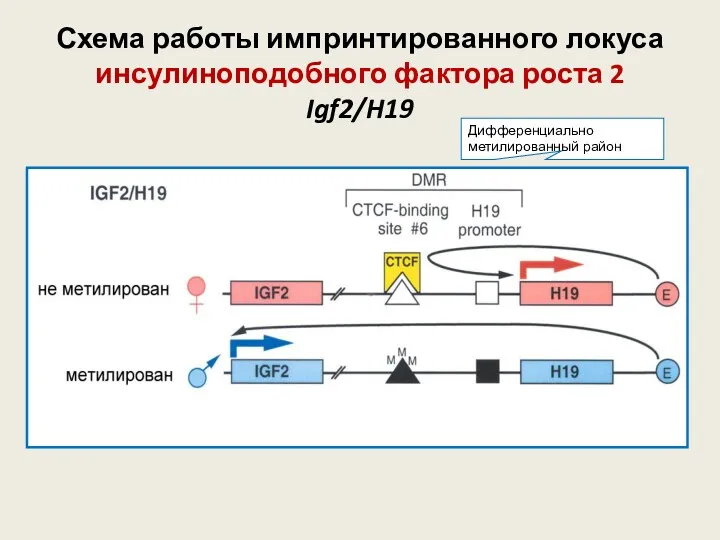
Схема работы импринтированного локуса инсулиноподобного фактора роста 2
Igf2/H19
Дифференциально метилированный район
Схема работы импринтированного локуса инсулиноподобного фактора роста 2
Igf2/H19
Дифференциально метилированный район
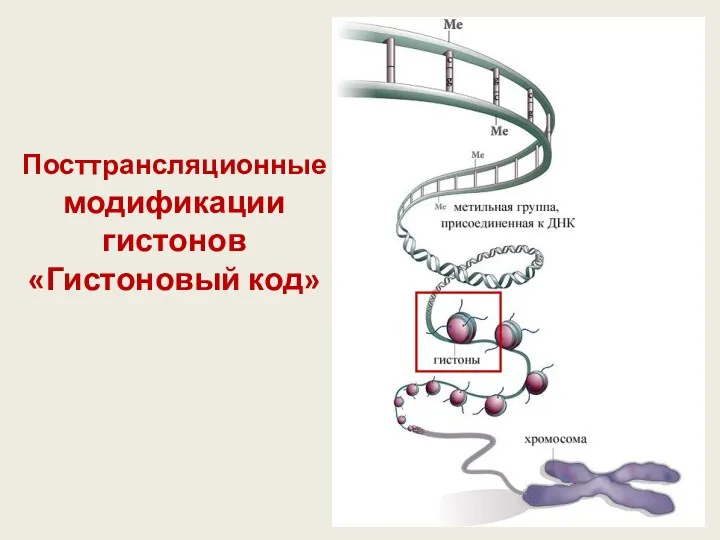
Посттрансляционные модификации
гистонов
«Гистоновый код»
Посттрансляционные модификации
гистонов
«Гистоновый код»
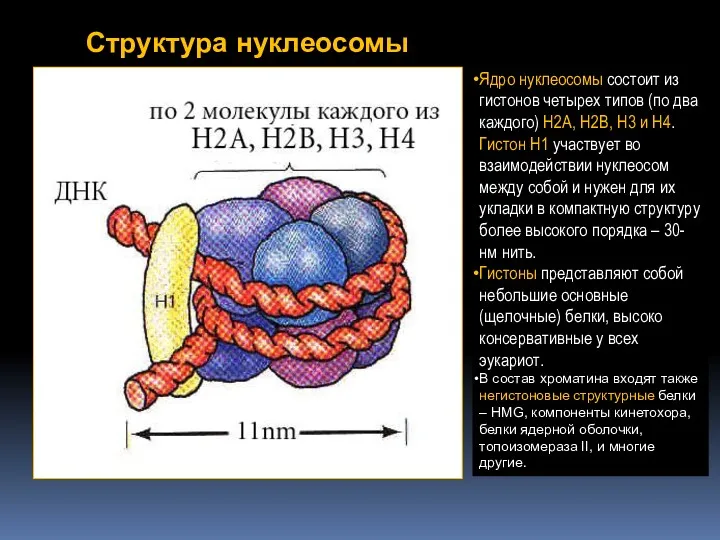
Структура нуклеосомы
Ядро нуклеосомы состоит из гистонов четырех типов (по два каждого)
Структура нуклеосомы
Ядро нуклеосомы состоит из гистонов четырех типов (по два каждого)

Нуклеосомы - рентгеноструктурный анализ в двух проекциях
Нуклеосомы - рентгеноструктурный анализ в двух проекциях
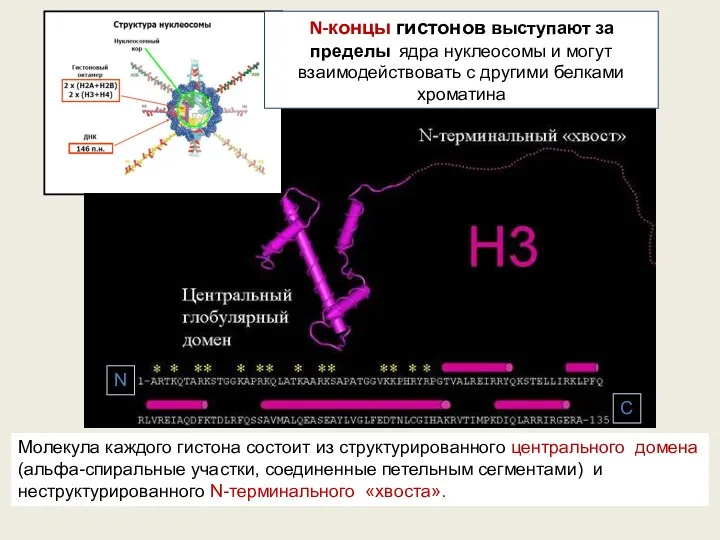
Молекула каждого гистона состоит из структурированного центрального домена
(альфа-спиральные участки, соединенные
Молекула каждого гистона состоит из структурированного центрального домена
(альфа-спиральные участки, соединенные
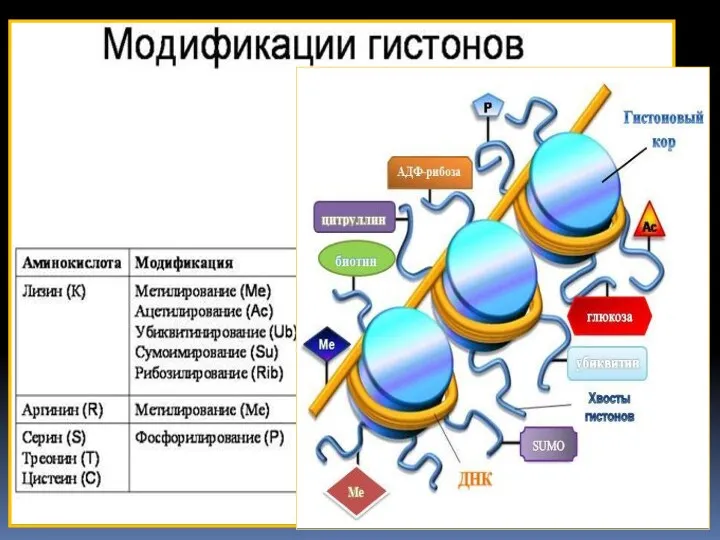

«Гистоновый код» –
совокупность ковалентных модификаций гистонов
«Гистоновый код» –
совокупность ковалентных модификаций гистонов

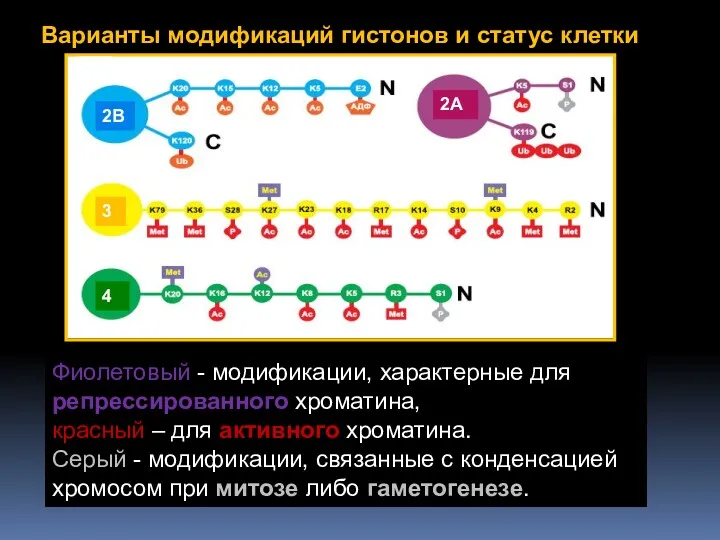
Фиолетовый - модификации, характерные для репрессированного хроматина,
красный – для активного
Фиолетовый - модификации, характерные для репрессированного хроматина,
красный – для активного
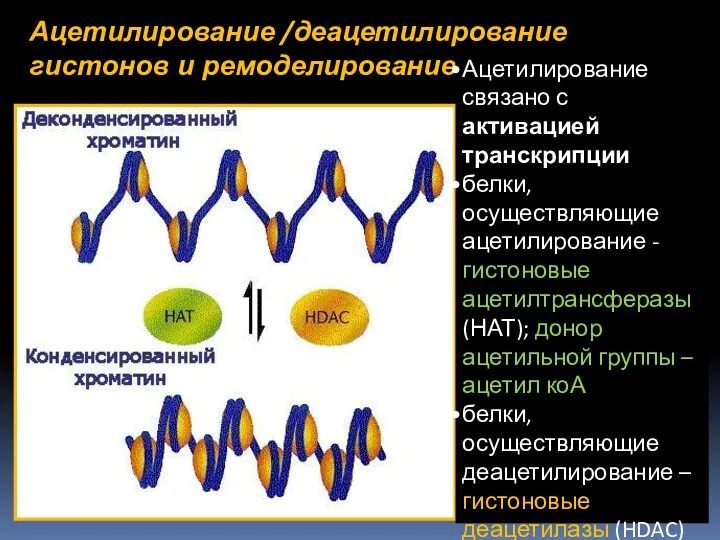
Ацетилирование /деацетилирование гистонов и ремоделирование хроматина
Ацетилирование связано с активацией транскрипции
белки,
Ацетилирование /деацетилирование гистонов и ремоделирование хроматина
Ацетилирование связано с активацией транскрипции
белки,
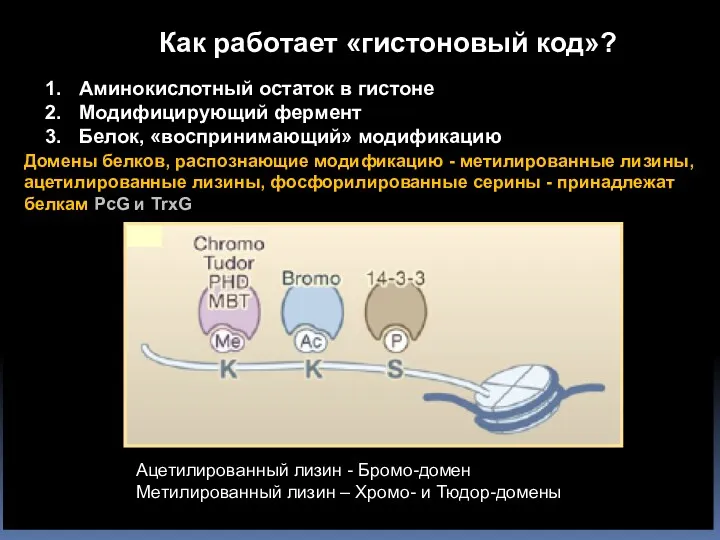
Домены белков, распознающие модификацию - метилированные лизины, ацетилированные лизины, фосфорилированные серины
Домены белков, распознающие модификацию - метилированные лизины, ацетилированные лизины, фосфорилированные серины

Как формируются и функционируют комплексы PcG и TxG?
Как формируются и функционируют комплексы PcG и TxG?

PcG-белки выявляются в составе многокомпонентных комплексов
1 комплекс, Polycomb Repressive Complex 1
PcG-белки выявляются в составе многокомпонентных комплексов
1 комплекс, Polycomb Repressive Complex 1

1. Связывание комплекса PhoRC
2. Привлечение комплекса PRC2 через взаимодействие с PhoRC
3.
1. Связывание комплекса PhoRC
2. Привлечение комплекса PRC2 через взаимодействие с PhoRC
3.
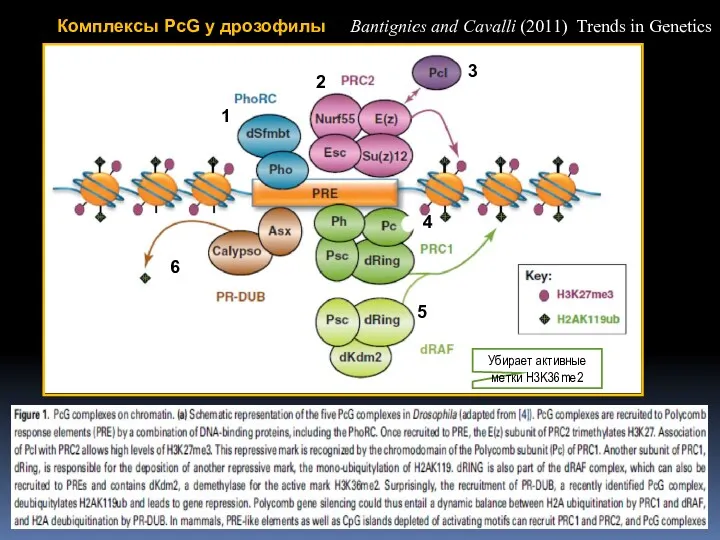
Комплексы PcG у дрозофилы
Bantignies and Cavalli (2011) Trends in Genetics
1
2
5
4
3
6
Убирает активные
Комплексы PcG у дрозофилы
Bantignies and Cavalli (2011) Trends in Genetics
1
2
5
4
3
6
Убирает активные
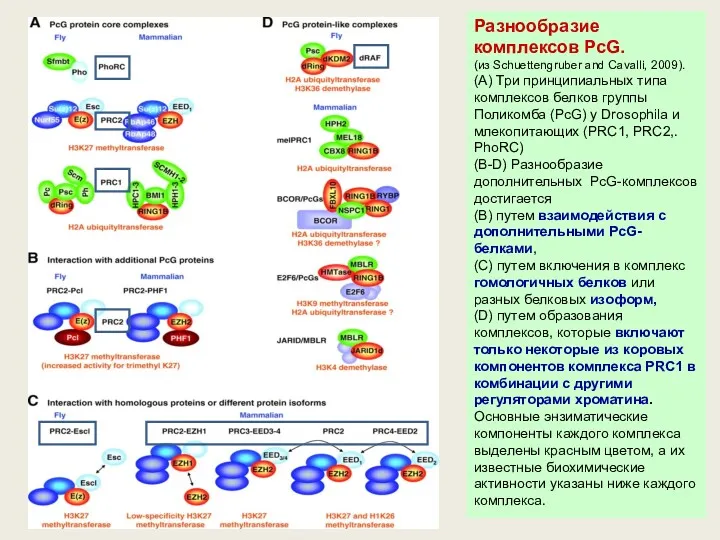
Разнообразие комплексов PcG.
(из Schuettengruber and Cavalli, 2009).
(A) Три принципиальных типа
Разнообразие комплексов PcG.
(из Schuettengruber and Cavalli, 2009).
(A) Три принципиальных типа
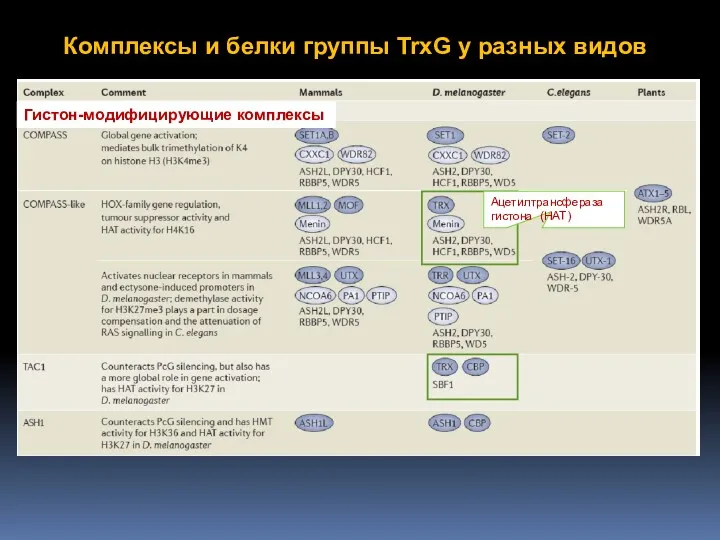
Комплексы и белки группы TrxG у разных видов
Комплексы и белки группы TrxG у разных видов
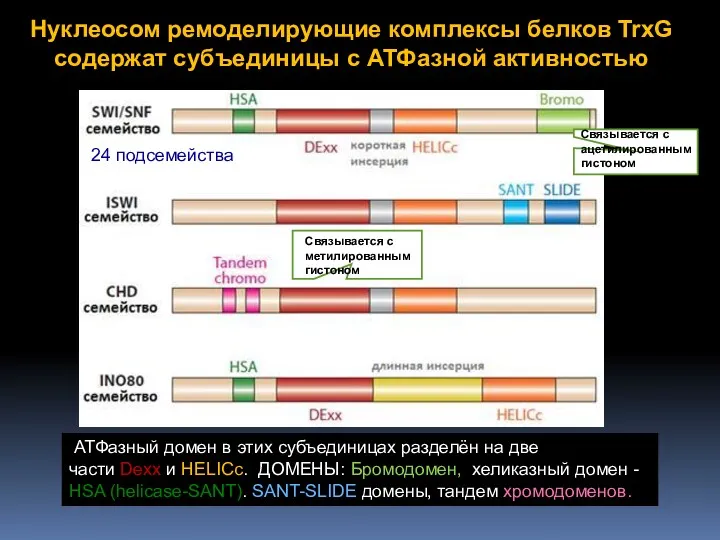
Нуклеосом ремоделирующие комплексы белков TrxG содержат субъединицы с АТФазной активностью
АТФазный домен в
Нуклеосом ремоделирующие комплексы белков TrxG содержат субъединицы с АТФазной активностью
АТФазный домен в
![Эволюционное разнообразие SWI/SNF-комплексов (из [Ho and Crabtree, 2010]). Гомологичные субъединицы](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/184893/slide-61.jpg)
Эволюционное разнообразие SWI/SNF-комплексов (из [Ho and Crabtree, 2010]).
Гомологичные субъединицы комплексов дрожжей
Эволюционное разнообразие SWI/SNF-комплексов (из [Ho and Crabtree, 2010]).
Гомологичные субъединицы комплексов дрожжей
Комплексы и белки группы TrxG у разных видов
Гистон-модифицирующие комплексы
Ацетилтрансфераза гистона
Комплексы и белки группы TrxG у разных видов
Гистон-модифицирующие комплексы
Ацетилтрансфераза гистона

Комплексы и белки группы TrxG у разных видов
АТФ-зависимые ремоделирующие комплексы
АТФаза
Brahma
Комплексы и белки группы TrxG у разных видов
АТФ-зависимые ремоделирующие комплексы
АТФаза
Brahma
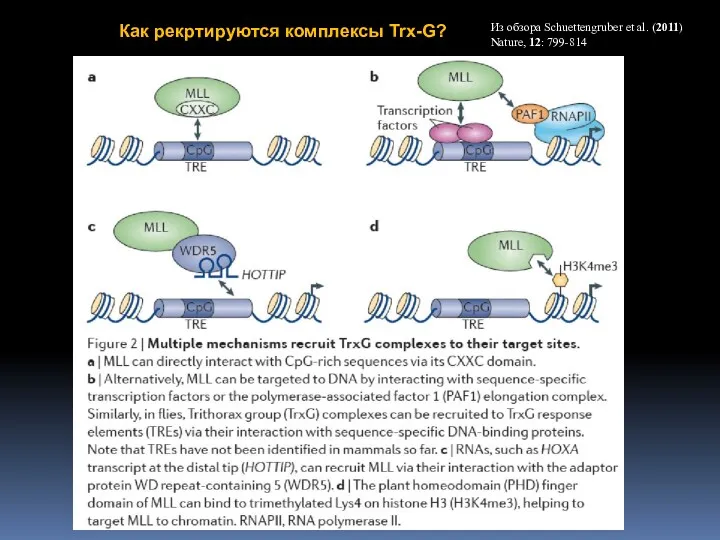
Из обзора Schuettengruber et al. (2011) Nature, 12: 799-814
Как рекртируются комплексы
Из обзора Schuettengruber et al. (2011) Nature, 12: 799-814
Как рекртируются комплексы
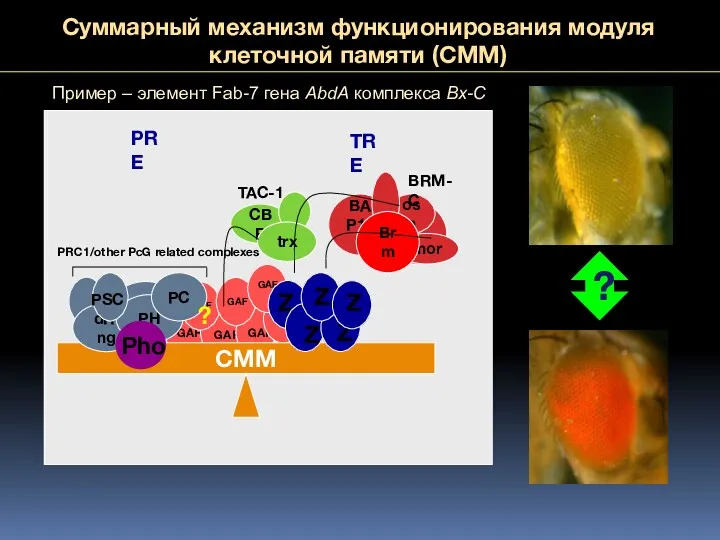
Суммарный механизм функционирования модуля клеточной памяти (CMM)
CBP
CMM
dRing
PH
PC
PRC1/other PcG related complexes
PSC
Z
Z
Z
Z
PRE
TRE
BAP111
osa
mor
BRM-C
Brm
?
trx
TAC-1
?
Z
Пример –
Суммарный механизм функционирования модуля клеточной памяти (CMM)
CBP
CMM
dRing
PH
PC
PRC1/other PcG related complexes
PSC
Z
Z
Z
Z
PRE
TRE
BAP111
osa
mor
BRM-C
Brm
?
trx
TAC-1
?
Z
Пример –
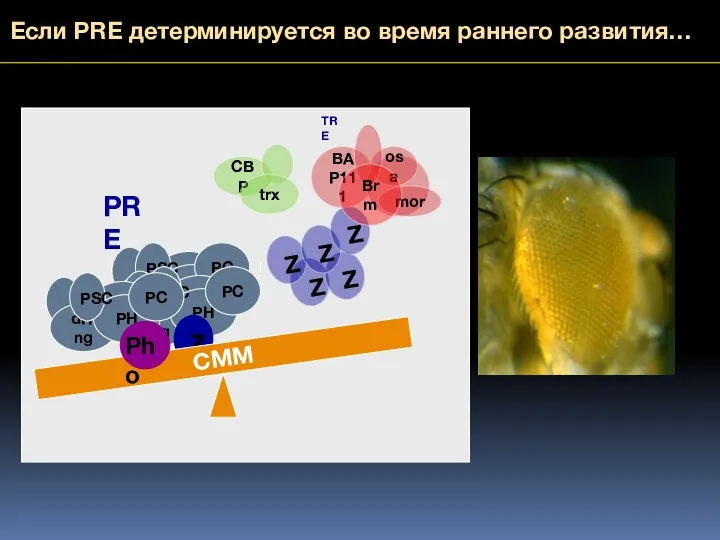
Если PRE детерминируется во время раннего развития…
dRing
PH
PC
PSC
dRing
PH
PC
PSC
CMM
dRing
PH
PC
PSC
PRE
CBP
Z
Z
Z
Z
Z
BAP111
osa
mor
Brm
trx
TRE
Если PRE детерминируется во время раннего развития…
dRing
PH
PC
PSC
dRing
PH
PC
PSC
CMM
dRing
PH
PC
PSC
PRE
CBP
Z
Z
Z
Z
Z
BAP111
osa
mor
Brm
trx
TRE
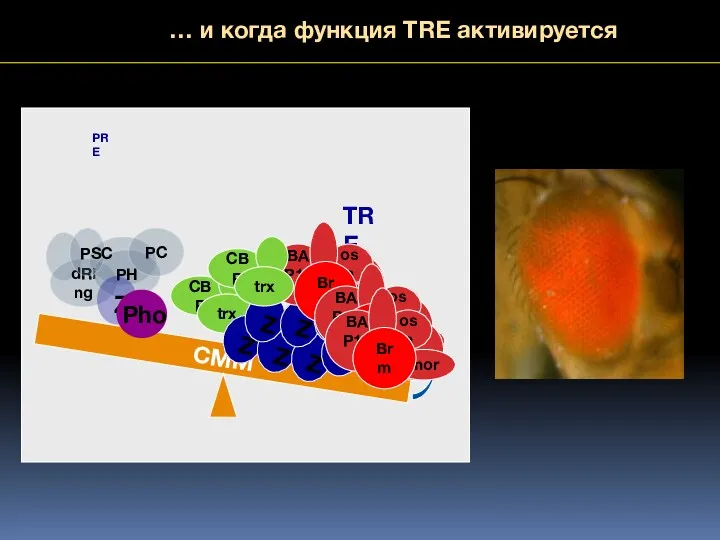
… и когда функция TRE активируется
CMM
PRE
TRE
… и когда функция TRE активируется
CMM
PRE
TRE

Как модули «клеточной памяти» передают информацию дочерним клеткам ?
Или
Как стабильно
Как модули «клеточной памяти» передают информацию дочерним клеткам ?
Или
Как стабильно
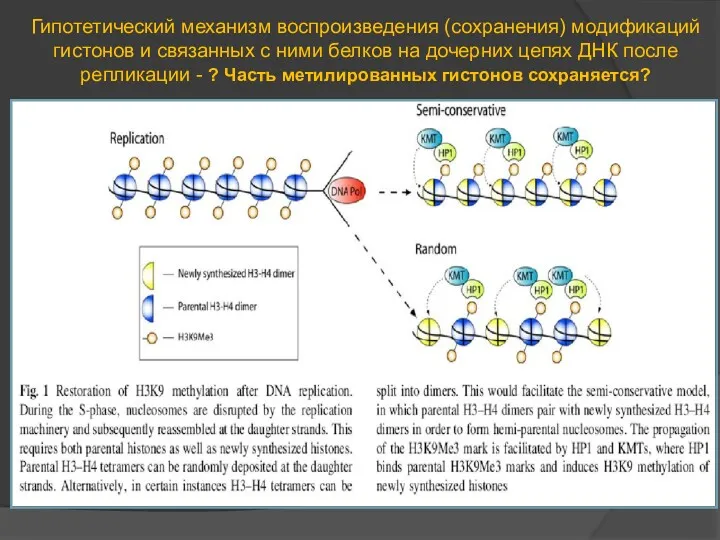
Гипотетический механизм воспроизведения (сохранения) модификаций гистонов и связанных с ними белков
Гипотетический механизм воспроизведения (сохранения) модификаций гистонов и связанных с ними белков

TrxG and PcG Proteins but Not Methylated Histones Remain Associated with
TrxG and PcG Proteins but Not Methylated Histones Remain Associated with
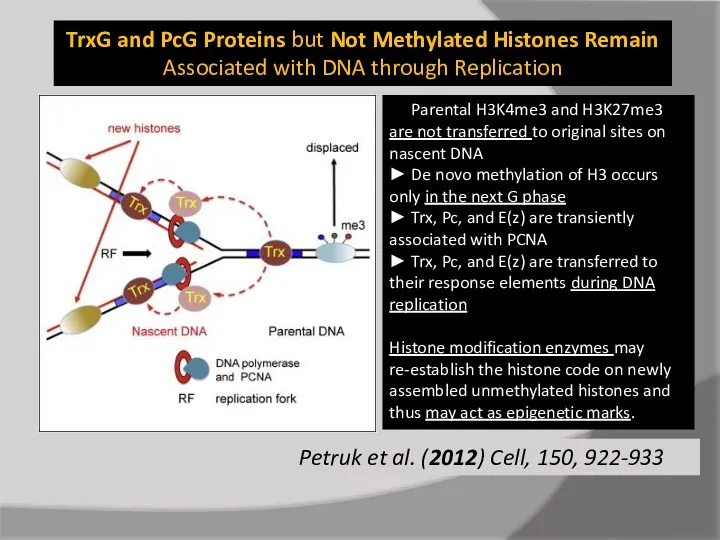
TrxG and PcG Proteins but Not Methylated Histones Remain Associated with
TrxG and PcG Proteins but Not Methylated Histones Remain Associated with
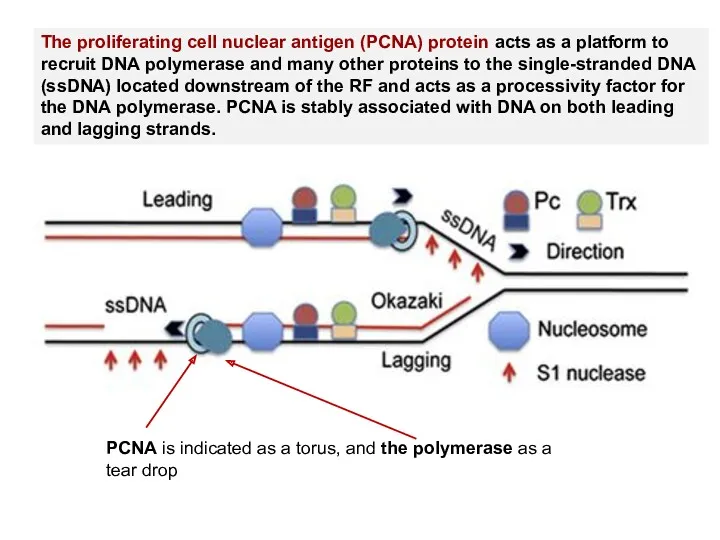
The proliferating cell nuclear antigen (PCNA) protein acts as a platform
The proliferating cell nuclear antigen (PCNA) protein acts as a platform
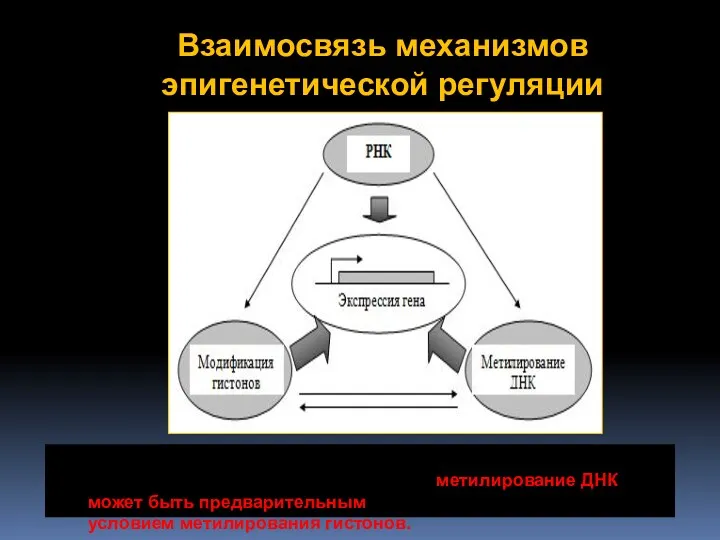
Взаимосвязь механизмов эпигенетической регуляции
Процессы модификации ДНК и хроматина отражают разные стороны
Взаимосвязь механизмов эпигенетической регуляции
Процессы модификации ДНК и хроматина отражают разные стороны
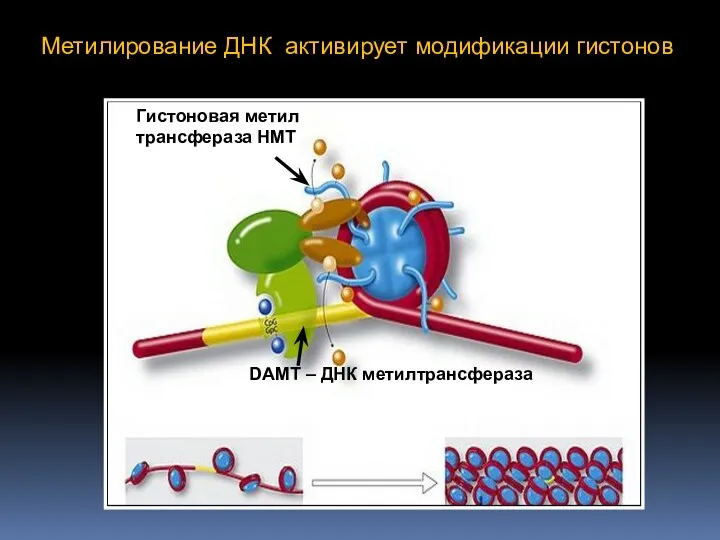
Метилирование ДНК активирует модификации гистонов
DAMT – ДНК метилтрансфераза
Гистоновая метил
трансфераза HMT
Метилирование ДНК активирует модификации гистонов
DAMT – ДНК метилтрансфераза
Гистоновая метил
трансфераза HMT

Мир некодирующих РНК (нкРНК)
Недавние исследования показали, что большая часть генома человека
Мир некодирующих РНК (нкРНК)
Недавние исследования показали, что большая часть генома человека
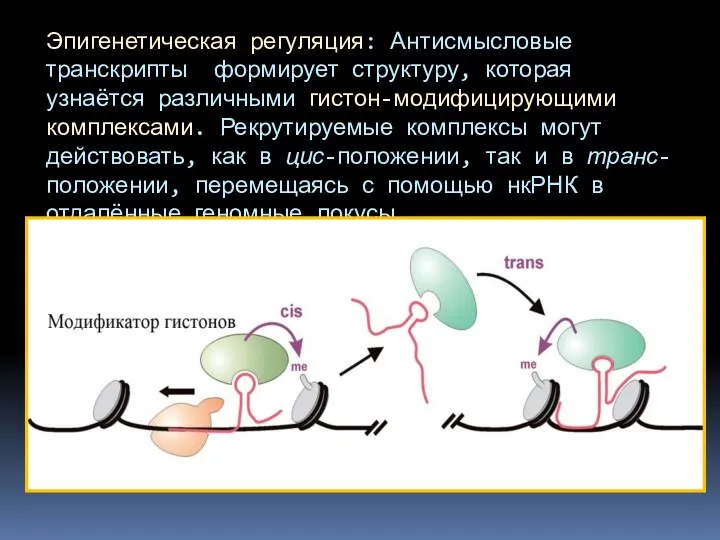
Эпигенетическая регуляция: Антисмысловые транскрипты формирует структуру, которая узнаётся различными гистон-модифицирующими комплексами.
Эпигенетическая регуляция: Антисмысловые транскрипты формирует структуру, которая узнаётся различными гистон-модифицирующими комплексами.
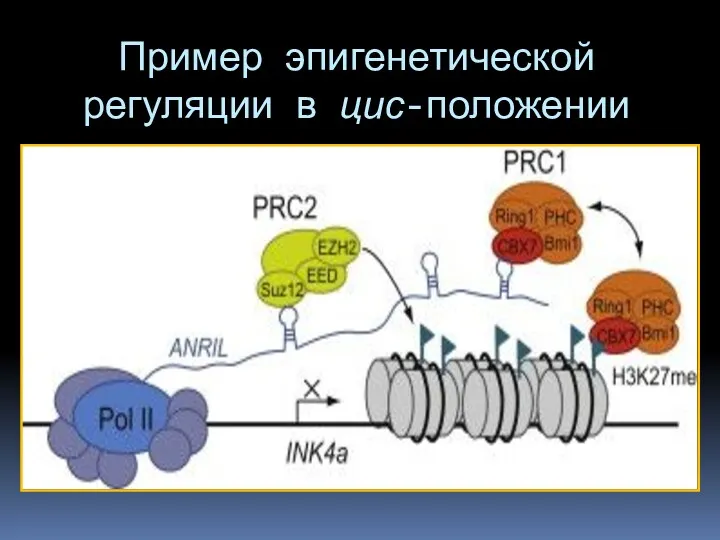
Пример эпигенетической регуляции в цис-положении
Пример эпигенетической регуляции в цис-положении
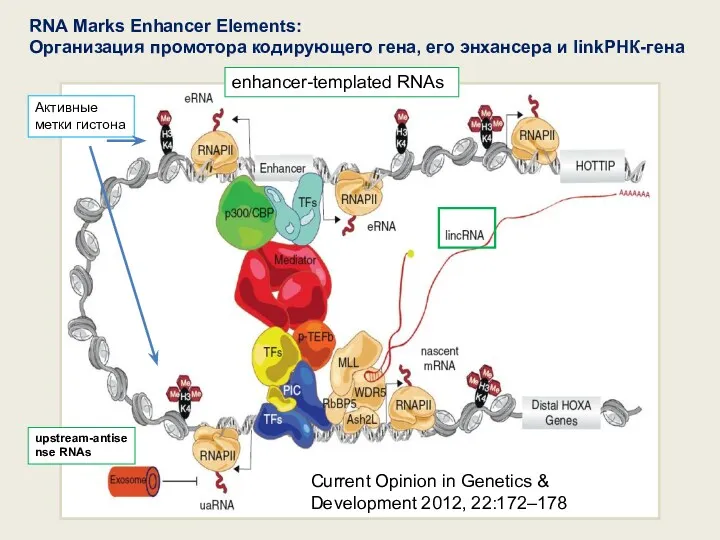
Current Opinion in Genetics & Development 2012, 22:172–178
upstream-antisense RNAs
enhancer-templated RNAs
RNA Marks Enhancer Elements:
Организация
Current Opinion in Genetics & Development 2012, 22:172–178
upstream-antisense RNAs
enhancer-templated RNAs
RNA Marks Enhancer Elements:
Организация
 III, IV, VI жұптар – көз алмасын қозғайтын, шығыр, әкеткіш нервтері
III, IV, VI жұптар – көз алмасын қозғайтын, шығыр, әкеткіш нервтері Полиомиелит, энтеровирусные инфекции
Полиомиелит, энтеровирусные инфекции Перитонит. Классификация перитонита
Перитонит. Классификация перитонита Нуклеопротеидттер алмасуының бұзылуы. Подагра
Нуклеопротеидттер алмасуының бұзылуы. Подагра Этиологическая и патогенетическая классификация анемий
Этиологическая и патогенетическая классификация анемий Туберкулез почек, мочевыводящей системы и мужских половых органов
Туберкулез почек, мочевыводящей системы и мужских половых органов Беременность и заболевания почек
Беременность и заболевания почек Патофизиология экстремальных состояний
Патофизиология экстремальных состояний Гіпертонічна хвороба
Гіпертонічна хвороба Внутрибольничные инфекции
Внутрибольничные инфекции Медицинская защита населения и спасателей в чрезвычайных ситуациях
Медицинская защита населения и спасателей в чрезвычайных ситуациях Телесно-ориентированные техники в психологической работе с паллиативными детьми раннего и младенческого возраста
Телесно-ориентированные техники в психологической работе с паллиативными детьми раннего и младенческого возраста Алгоритм действий медицинской сестры при почечной колике
Алгоритм действий медицинской сестры при почечной колике Тиреотоксикоз. Диффузный токсический зоб. Гипотиреоз
Тиреотоксикоз. Диффузный токсический зоб. Гипотиреоз Иммунитет как механизм регуляции и защиты
Иммунитет как механизм регуляции и защиты Нейродегенеративные заболевания
Нейродегенеративные заболевания Модель Гиппократа и проблема доверия к профессии врача
Модель Гиппократа и проблема доверия к профессии врача Классификация детей с ОВЗ
Классификация детей с ОВЗ Косметологиядағы криотерапия
Косметологиядағы криотерапия Компания Capsid Illuminesca. Вирусология
Компания Capsid Illuminesca. Вирусология Здоровый образ жизни
Здоровый образ жизни Рациональное питание
Рациональное питание Государственная система управления здравоохранением. Современные формы управления в системе здравоохранения
Государственная система управления здравоохранением. Современные формы управления в системе здравоохранения Гипс. Фиксация в кювете
Гипс. Фиксация в кювете Стерилизация тиімділігін бақылау әдістері
Стерилизация тиімділігін бақылау әдістері Местные средства профилактики кариеса
Местные средства профилактики кариеса Потери. Смерть. Горе
Потери. Смерть. Горе Рак яичников
Рак яичников