- Биология опухолевого роста

Содержание
- 2. Определение Опухоль – это патологическое разрастание тканей, состоящих из качественно изменившихся клеток, ставших атипичными в отношении
- 3. Основные этапы опухолевого роста
- 4. Hallmarks of cancer
- 5. Поддержка сигнала к пролиферации Деление клеток в популяции контролируется ростовыми факторами (митогенами). Ростовые факторы, связываются с
- 6. Протоонкоген – онкоген – онкопротеин
- 7. Онкогенные вирусы
- 8. Рецепторы факторов роста
- 9. Способы поддержки постоянного сигнала к пролиферации Самостоятельная секреция факторов роста клетками опухоли Стимулирование близлежащих стромальных клеток
- 10. Сигнальные пути MAPK
- 13. Уклонение от супрессоров пролиферации В нормальных клетках существуют системы защиты, подавляющие бесконтрольный рост клеток. Данные системы
- 17. Уклонение от апоптоза Процесс апоптоза является естественным препятствием на пути к росту опухоли. В рамках многоклеточного
- 18. Механизмы ухода от апоптоза. Нарушении функции гена p53 Увеличение экспресии антиапоптотических регуляторов (BCL-2) Увеличение экспресии факторов
- 19. Внутренний и внешний пути апоптоза
- 20. Обретение репликативного бессмертия Подавляющее большинство клеток имеют предел количеству делений (около 60 делений), после чего они
- 23. Предположительная модель положительной обратной связи, описывающей взаимодействие обратной транскриптазы теломеразы (TERT) с Wnt/b-catenin и NF-kB signaling
- 24. Ангиогенез Опухоли необходимы новые сосуды, для поддержания её развития и выживания. Запуск ангиогенеза происходит еще на
- 25. Angiogenic siwtch
- 26. Лимфоангиогенез и ангиогенез
- 28. Предотвращение развития сосудов в опухоли.
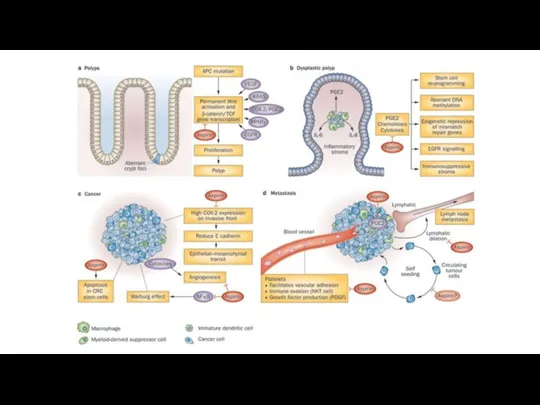
- 29. Инвазия и метастазирование Клетки опухолей способны терять связь с окружающим их внеклеточным матриксом и другими клетками.
- 30. Эпительиально-мезенхимальный переход Среди механизмов EMT можно выделить несколько ключевых моментов: Подавление экспрессии гена Е-кадгерина (E-cadherin (CDH1))
- 31. Эпителиально-мезенхимальный переход Первый этап эпителиально - мезенхимального перехода – разрушение эпителиальных межклеточных контактов, плотных соединений, слипающихся
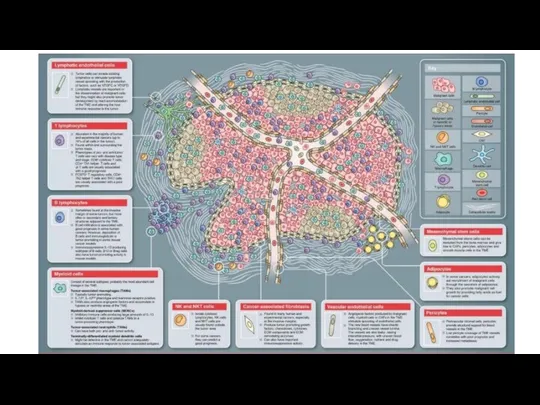
- 32. Опухолевое микроокружение
- 33. EMT – это локальный феномен в первичной опухоли, который происходит при взаимодействии клеток карциномы с микроокружением,
- 35. Стволовые клетки опухоли Нестабильность генома, иммортализация, воспаление, изменение энергетики, отказ от самоуничтожения, гипоксия клетки приводит к
- 36. На данный момент предполагаетсяя, что РСК, не являются истинно стволовыми клетками, они лишь обладают их некоторыми
- 39. Метастазирование Процесс метастазирования разделен на несколько этапов: инвазия в окружающие ткани, интравазация, экстравазация, микрометастазирование, колонизация.
- 40. Органотропность
- 41. Формирование преметастатической ниши
- 42. Процессы, происходящие на ранних этапах метастазирования
- 43. Процесс гематогенного дисеминирования
- 44. Hallmarks of cancer
- 45. Изменение клеточной энергетики Опухолевые клетки используют анаэробный гликолиз даже при условии доступа к достаточному количеству кислорода.
- 46. Метаболические пути, активные в пролиферирующих клетках, напрямую управляются сигнальными путями, в которых участвуют онкогены и онкосупрессоры.
- 47. A Продукция ингибиторных метаболитов, таких как R-2‑гидроксиглутарат (R-2HG) и S-2HG мутантной изоцитрат дегидрогеназой(IDH) и активность лактат
- 48. Нестабильность генома Приобретение черт из Hallmarks of cancer зависит от нестабильности генома клетки. Каждая вышеперечисленная мутация
- 49. Накопление изменений в ДНК клетки, приводит к опухолевому росту.
- 51. DNA damage response
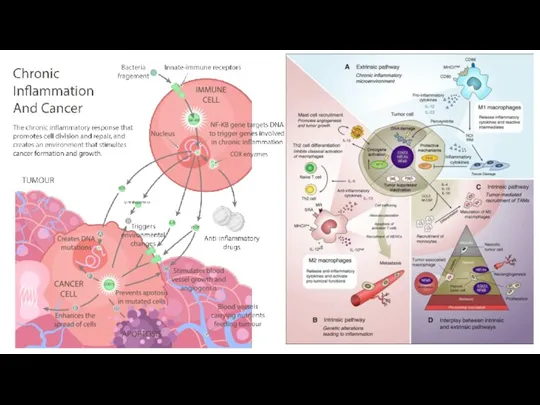
- 53. Иммуноонкология В этом разделе объединены механизмы ухода опухоли от иммунного ответа и вклад воспаления в онкогенез.
- 54. Концепция иммунного надзора
- 55. Благодаря концепции иммунного надзора были открыты опухолевые антигены
- 56. Разработаны современные иммунотерапевтические препараты
- 57. Проходят клинические испытания новые терапевтические методы
- 58. Концепция иммунного редактирования (Immunoediting)
- 61. Myeloid derived suppressor cell
- 62. Tumor associated macrophage
- 65. Скачать презентацию

Определение
Опухоль – это патологическое разрастание тканей, состоящих из качественно изменившихся клеток,
Определение
Опухоль – это патологическое разрастание тканей, состоящих из качественно изменившихся клеток,

Основные этапы опухолевого роста
Основные этапы опухолевого роста
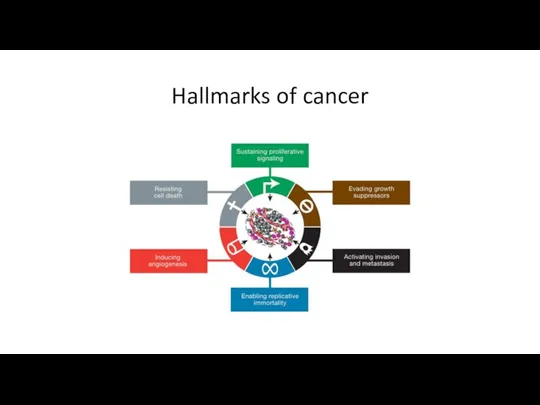
Hallmarks of cancer
Hallmarks of cancer

Поддержка сигнала к пролиферации
Деление клеток в популяции контролируется ростовыми факторами (митогенами).
Поддержка сигнала к пролиферации
Деление клеток в популяции контролируется ростовыми факторами (митогенами).
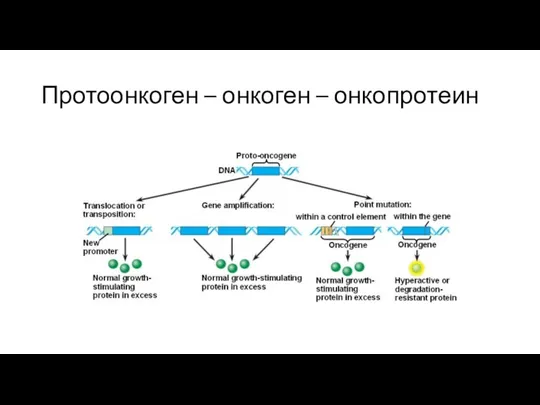
Протоонкоген – онкоген – онкопротеин
Протоонкоген – онкоген – онкопротеин
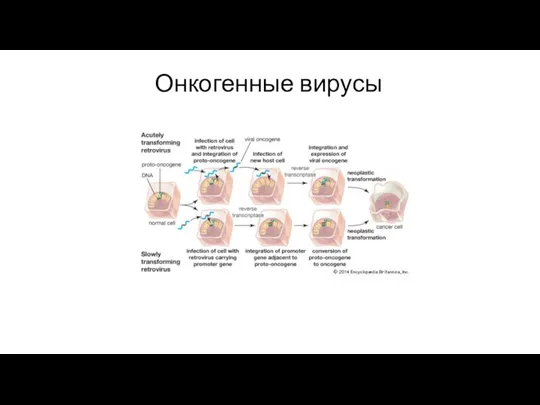
Онкогенные вирусы
Онкогенные вирусы
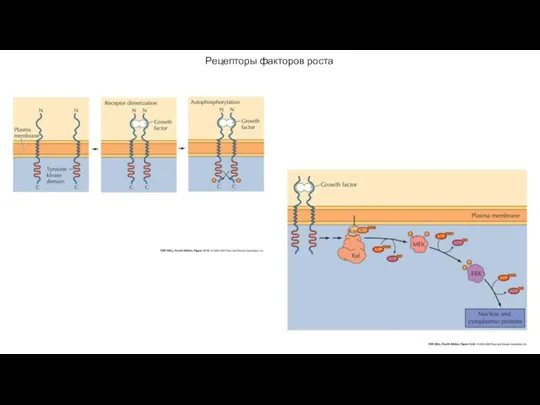
Рецепторы факторов роста
Рецепторы факторов роста

Способы поддержки постоянного сигнала к пролиферации
Самостоятельная секреция факторов роста клетками опухоли
Стимулирование
Способы поддержки постоянного сигнала к пролиферации
Самостоятельная секреция факторов роста клетками опухоли
Стимулирование
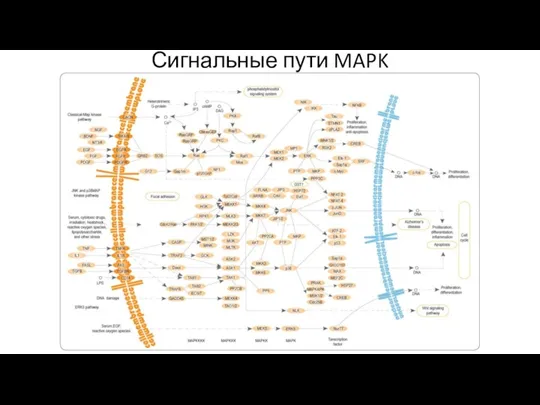
Сигнальные пути MAPK
Сигнальные пути MAPK
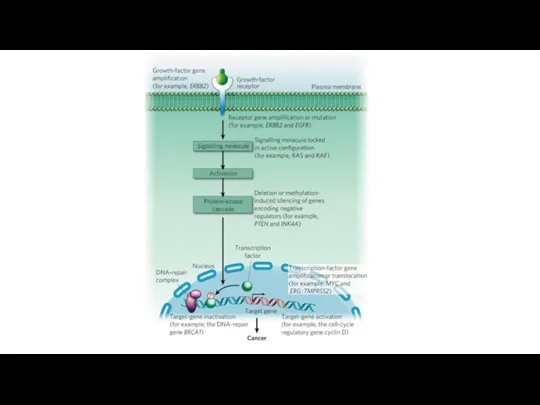
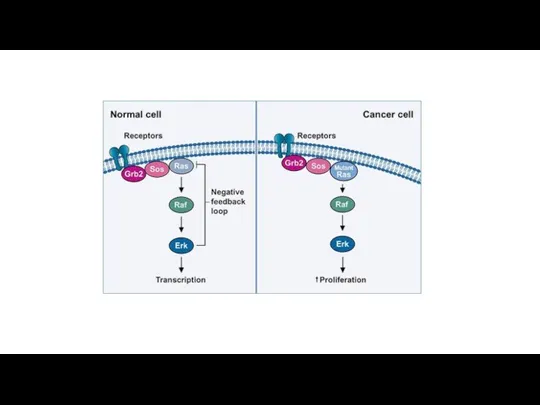

Уклонение от супрессоров пролиферации
В нормальных клетках существуют системы защиты, подавляющие бесконтрольный
Уклонение от супрессоров пролиферации
В нормальных клетках существуют системы защиты, подавляющие бесконтрольный

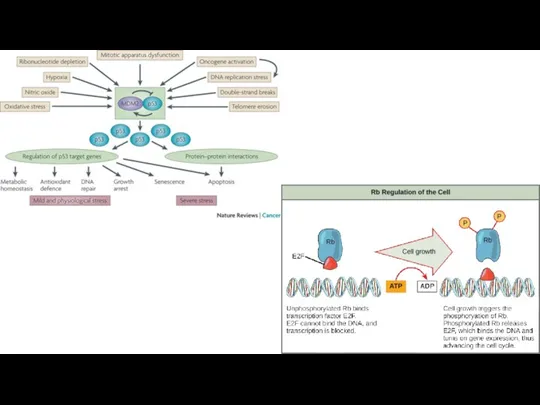
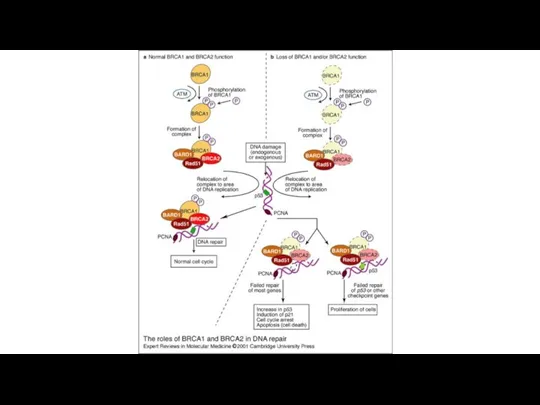

Уклонение от апоптоза
Процесс апоптоза является естественным препятствием на пути к росту
Уклонение от апоптоза
Процесс апоптоза является естественным препятствием на пути к росту

Механизмы ухода от апоптоза.
Нарушении функции гена p53
Увеличение экспресии антиапоптотических регуляторов (BCL-2)
Увеличение
Механизмы ухода от апоптоза.
Нарушении функции гена p53
Увеличение экспресии антиапоптотических регуляторов (BCL-2)
Увеличение
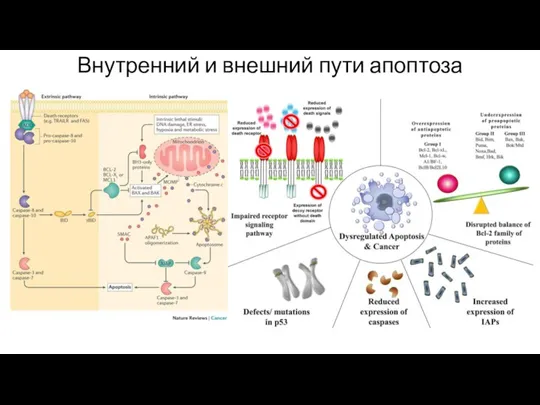
Внутренний и внешний пути апоптоза
Внутренний и внешний пути апоптоза

Обретение репликативного бессмертия
Подавляющее большинство клеток имеют предел количеству делений (около 60
Обретение репликативного бессмертия
Подавляющее большинство клеток имеют предел количеству делений (около 60
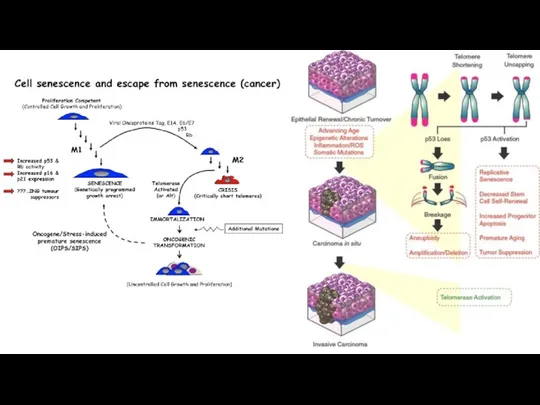
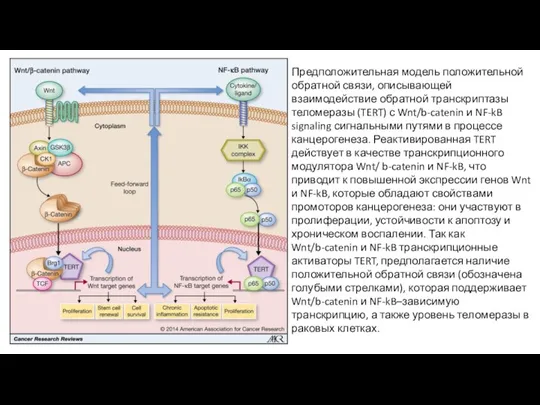
Предположительная модель положительной обратной связи, описывающей взаимодействие обратной транскриптазы теломеразы (TERT)
Предположительная модель положительной обратной связи, описывающей взаимодействие обратной транскриптазы теломеразы (TERT)

Ангиогенез
Опухоли необходимы новые сосуды, для поддержания её развития и выживания. Запуск
Ангиогенез
Опухоли необходимы новые сосуды, для поддержания её развития и выживания. Запуск
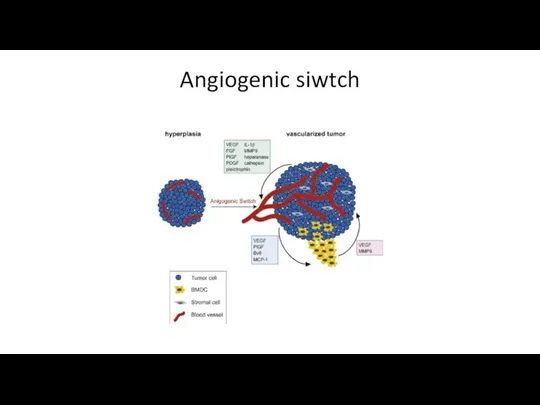
Angiogenic siwtch
Angiogenic siwtch
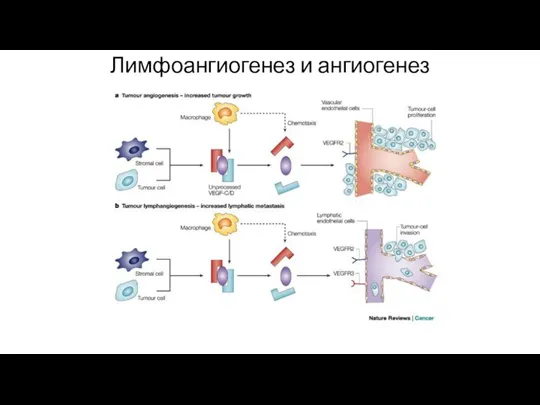
Лимфоангиогенез и ангиогенез
Лимфоангиогенез и ангиогенез
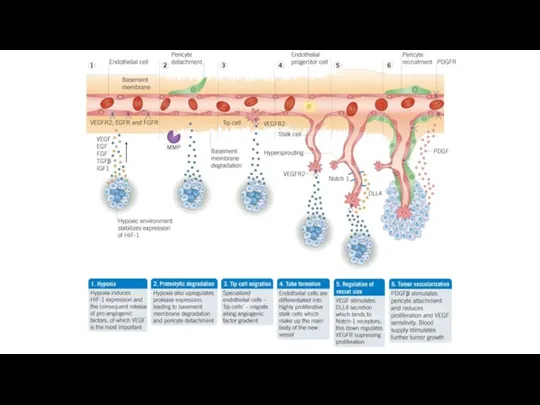
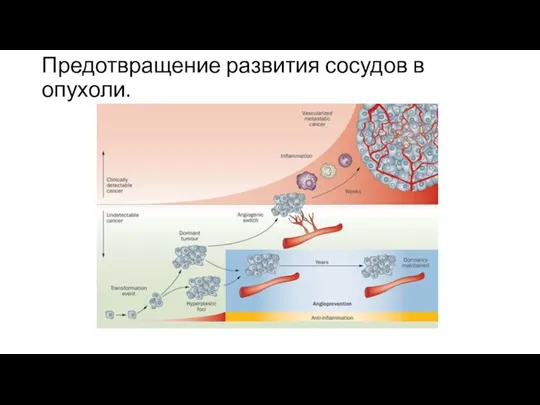
Предотвращение развития сосудов в опухоли.
Предотвращение развития сосудов в опухоли.

Инвазия и метастазирование
Клетки опухолей способны терять связь с окружающим их внеклеточным
Инвазия и метастазирование
Клетки опухолей способны терять связь с окружающим их внеклеточным
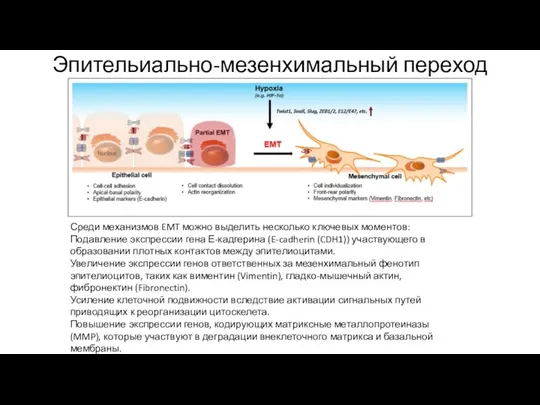
Эпительиально-мезенхимальный переход
Среди механизмов EMT можно выделить несколько ключевых моментов:
Подавление экспрессии гена
Эпительиально-мезенхимальный переход
Среди механизмов EMT можно выделить несколько ключевых моментов:
Подавление экспрессии гена

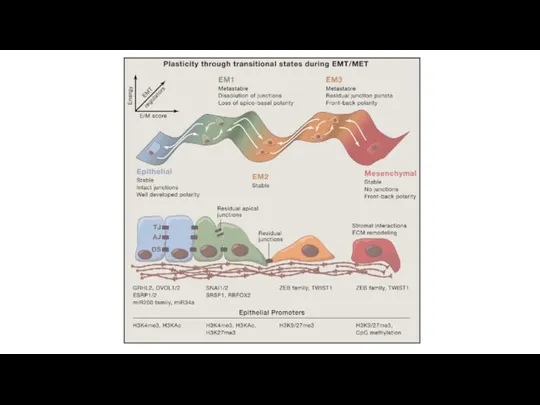
Эпителиально-мезенхимальный переход
Первый этап эпителиально - мезенхимального перехода – разрушение эпителиальных межклеточных
Эпителиально-мезенхимальный переход
Первый этап эпителиально - мезенхимального перехода – разрушение эпителиальных межклеточных

Опухолевое микроокружение
Опухолевое микроокружение
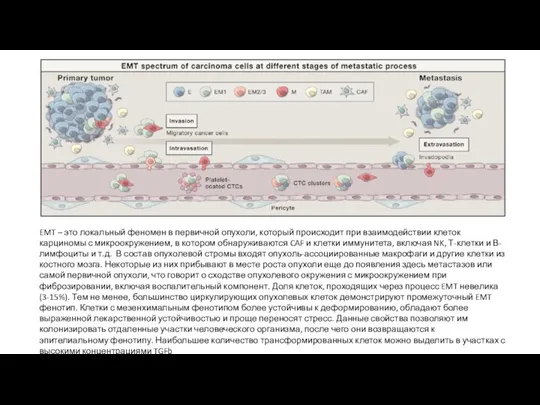
EMT – это локальный феномен в первичной опухоли, который происходит при
EMT – это локальный феномен в первичной опухоли, который происходит при

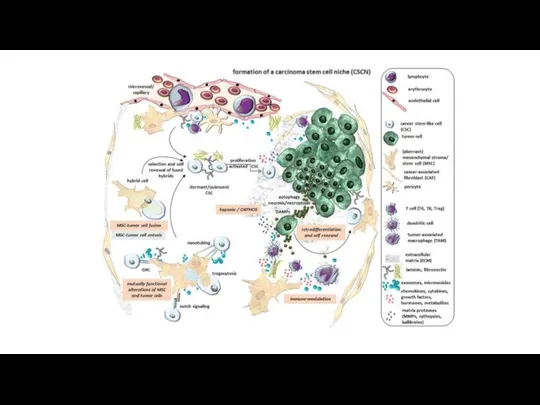
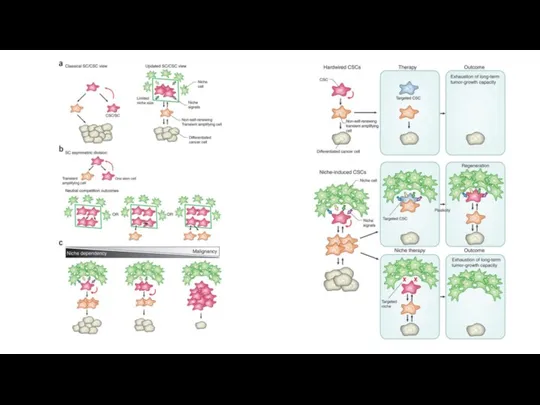
Стволовые клетки опухоли
Нестабильность генома, иммортализация, воспаление, изменение энергетики, отказ от
Стволовые клетки опухоли
Нестабильность генома, иммортализация, воспаление, изменение энергетики, отказ от

На данный момент предполагаетсяя, что РСК, не являются истинно стволовыми клетками,
На данный момент предполагаетсяя, что РСК, не являются истинно стволовыми клетками,

Метастазирование
Процесс метастазирования разделен на несколько этапов: инвазия в окружающие ткани, интравазация,
Метастазирование
Процесс метастазирования разделен на несколько этапов: инвазия в окружающие ткани, интравазация,
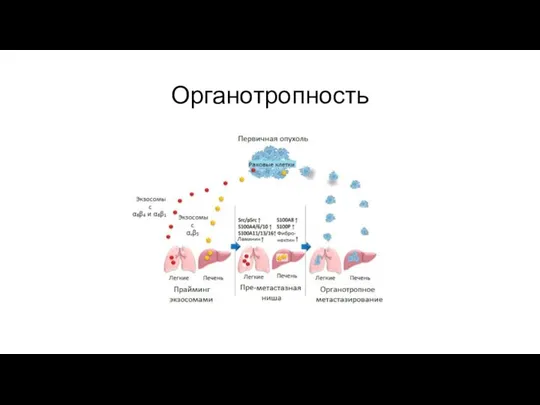
Органотропность
Органотропность
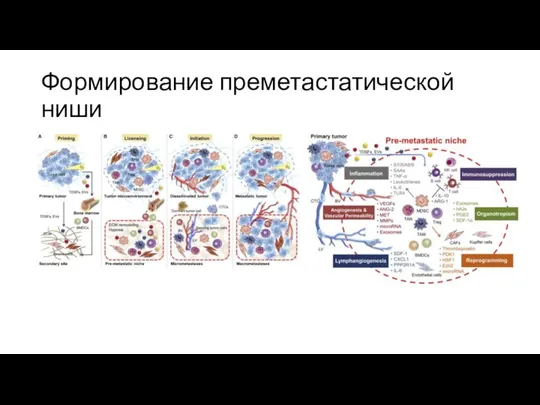
Формирование преметастатической ниши
Формирование преметастатической ниши
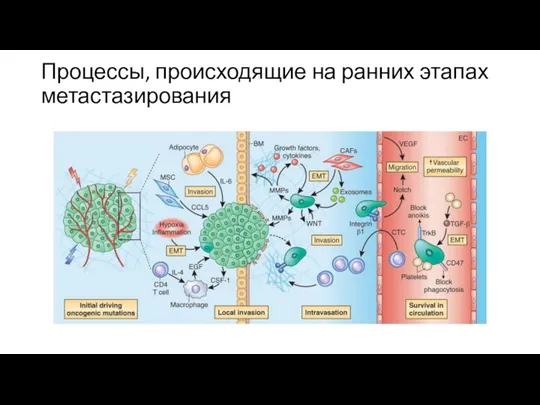
Процессы, происходящие на ранних этапах метастазирования
Процессы, происходящие на ранних этапах метастазирования
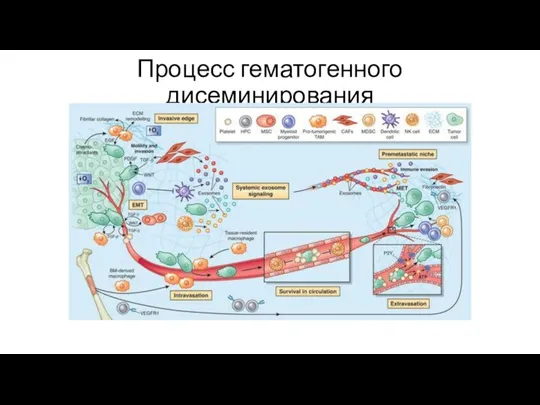
Процесс гематогенного дисеминирования
Процесс гематогенного дисеминирования
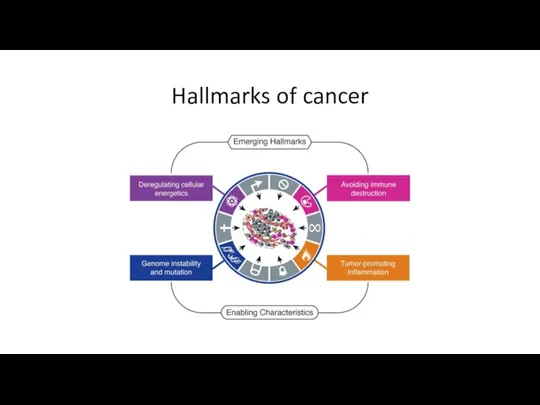
Hallmarks of cancer
Hallmarks of cancer

Изменение клеточной энергетики
Опухолевые клетки используют анаэробный гликолиз даже при условии доступа
Изменение клеточной энергетики
Опухолевые клетки используют анаэробный гликолиз даже при условии доступа
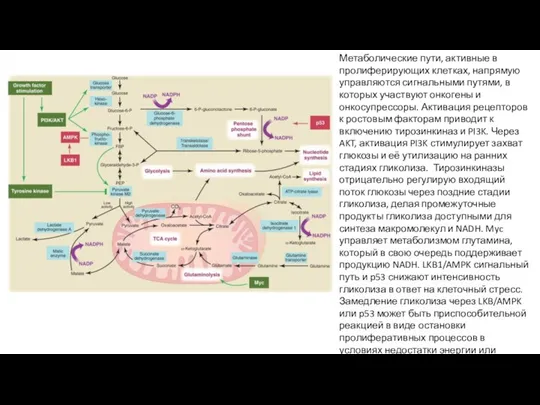
Метаболические пути, активные в пролиферирующих клетках, напрямую управляются сигнальными путями, в
Метаболические пути, активные в пролиферирующих клетках, напрямую управляются сигнальными путями, в
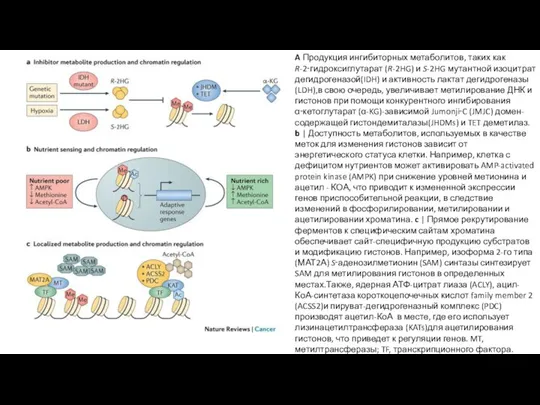
A Продукция ингибиторных метаболитов, таких как R-2‑гидроксиглутарат (R-2HG) и S-2HG мутантной
A Продукция ингибиторных метаболитов, таких как R-2‑гидроксиглутарат (R-2HG) и S-2HG мутантной

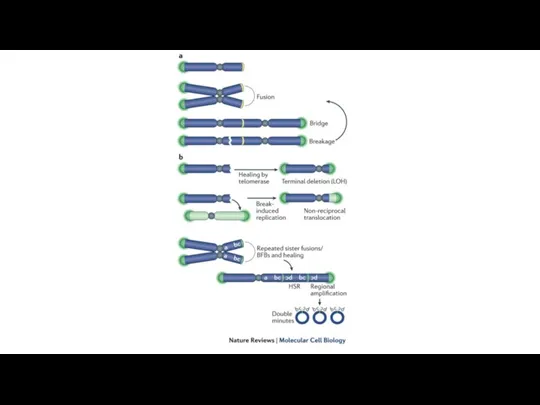
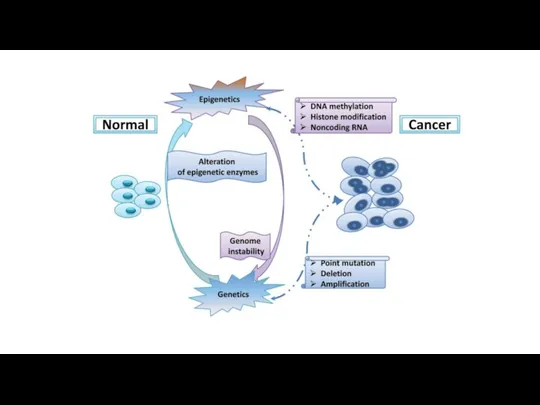
Нестабильность генома
Приобретение черт из Hallmarks of cancer зависит от нестабильности генома
Нестабильность генома
Приобретение черт из Hallmarks of cancer зависит от нестабильности генома

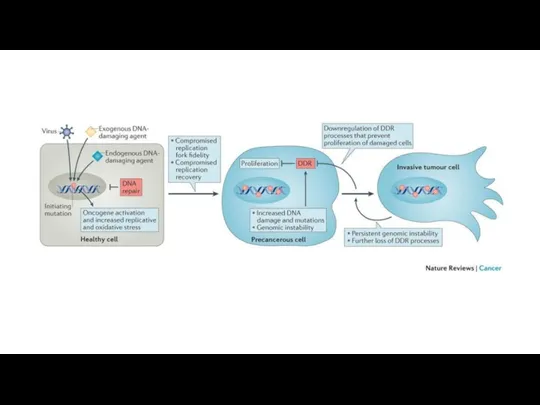
Накопление изменений в ДНК клетки, приводит к опухолевому росту.
Накопление изменений в ДНК клетки, приводит к опухолевому росту.
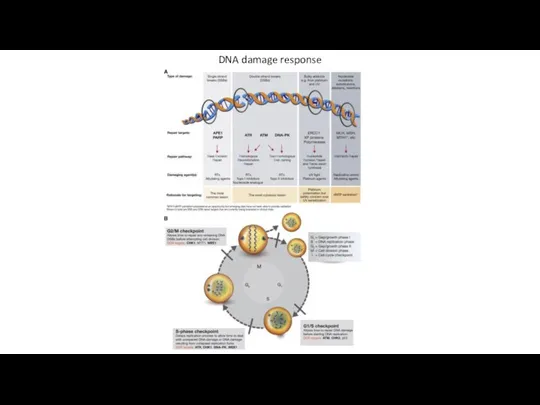
DNA damage response
DNA damage response

Иммуноонкология
В этом разделе объединены механизмы ухода опухоли от иммунного ответа
Иммуноонкология
В этом разделе объединены механизмы ухода опухоли от иммунного ответа
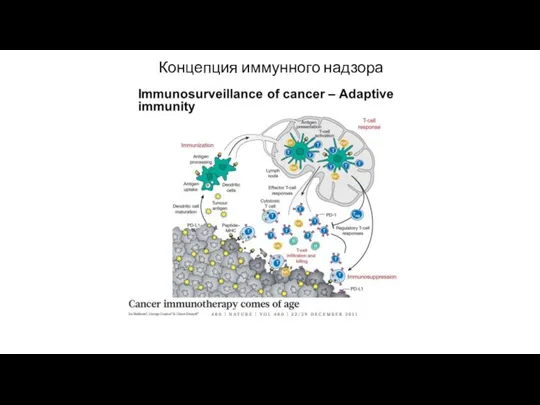
Концепция иммунного надзора
Концепция иммунного надзора
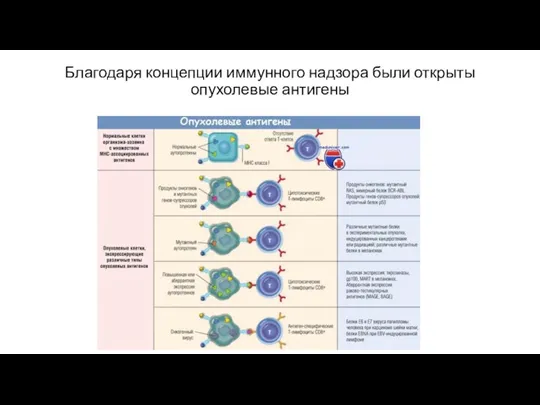
Благодаря концепции иммунного надзора были открыты опухолевые антигены
Благодаря концепции иммунного надзора были открыты опухолевые антигены
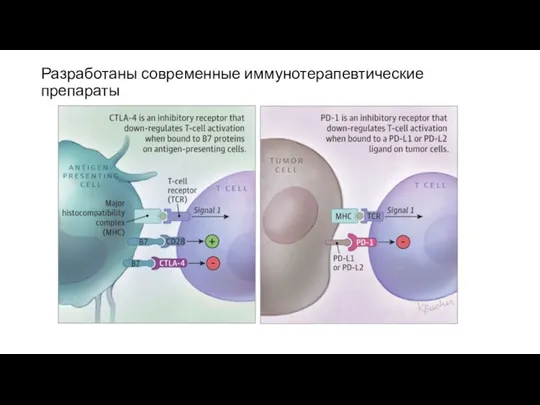
Разработаны современные иммунотерапевтические препараты
Разработаны современные иммунотерапевтические препараты
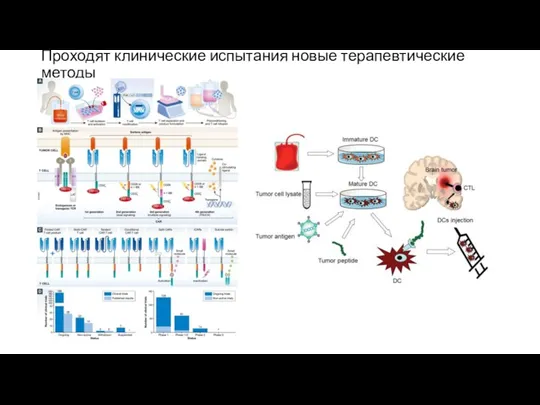
Проходят клинические испытания новые терапевтические методы
Проходят клинические испытания новые терапевтические методы

Концепция иммунного редактирования (Immunoediting)
Концепция иммунного редактирования (Immunoediting)
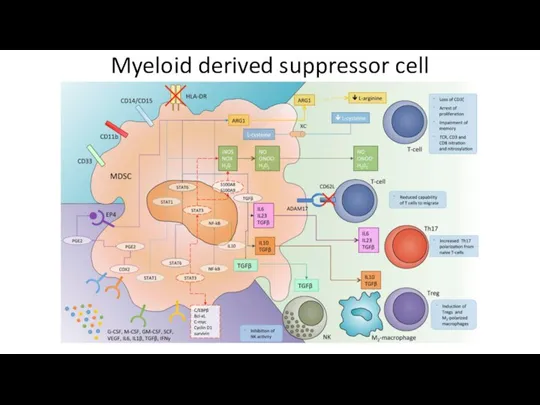
Myeloid derived suppressor cell
Myeloid derived suppressor cell
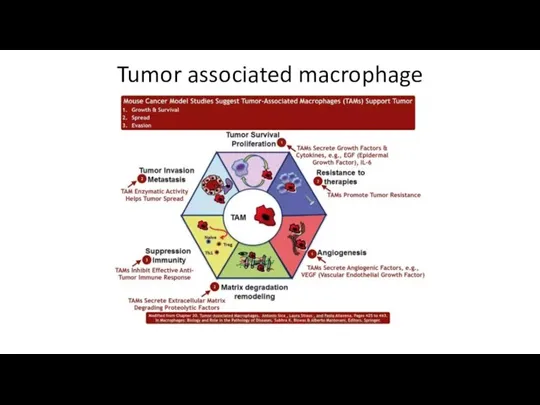
Tumor associated macrophage
Tumor associated macrophage
 Дія променевої енергії на організм тварини
Дія променевої енергії на організм тварини Өндірістік ортадағы жұмыс орнын қорғаудың қазіргі кездегі мәселелері
Өндірістік ортадағы жұмыс орнын қорғаудың қазіргі кездегі мәселелері Урологиядағы зерттеудің рентгенологиялық әдістері
Урологиядағы зерттеудің рентгенологиялық әдістері Судорожный синдром у детей. Эпидемиология судорожных состояний
Судорожный синдром у детей. Эпидемиология судорожных состояний Стирание твердых тканей зуба, физиологическое, патологическое
Стирание твердых тканей зуба, физиологическое, патологическое Правильное питание – здоровый ребенок
Правильное питание – здоровый ребенок Принципы преобразования органов хордовых
Принципы преобразования органов хордовых Кома жағдайлардың ажырату диагностикасы
Кома жағдайлардың ажырату диагностикасы Травма груди
Травма груди Физиология эндокринной системы
Физиология эндокринной системы Основы эпидемиологии, инфекционные заболевания
Основы эпидемиологии, инфекционные заболевания Патофизиология CCC
Патофизиология CCC Хроническая ревматическая болезнь сердца (ХРБС)
Хроническая ревматическая болезнь сердца (ХРБС) Рак лёгких. Хирургическое лечение
Рак лёгких. Хирургическое лечение Симптоматические артериальные гипертензии
Симптоматические артериальные гипертензии Рутина доения
Рутина доения Интраоперационный мониторинг
Интраоперационный мониторинг Возбудители кокковых инфекций
Возбудители кокковых инфекций Свойства материалов и их влияние на ткани зуба. Материаловедение. Лекция № 1. Тема 2
Свойства материалов и их влияние на ткани зуба. Материаловедение. Лекция № 1. Тема 2 Аддиктивті бұзылыстар. Түрлері. Химиялық және химиялық емес аддиктивті бұзылыстардың клиникалық ерекшеліктері
Аддиктивті бұзылыстар. Түрлері. Химиялық және химиялық емес аддиктивті бұзылыстардың клиникалық ерекшеліктері Организация лечебно-эвакуационного обеспечения населения при чрезвычайных ситуациях
Организация лечебно-эвакуационного обеспечения населения при чрезвычайных ситуациях Инфекционный эндокардит
Инфекционный эндокардит Особиста програма зміцнення здоров’я
Особиста програма зміцнення здоров’я Микробиологическая диагностика зоонозных инфекций
Микробиологическая диагностика зоонозных инфекций Вибрационная болезнь. Холодовой нейроваскулит
Вибрационная болезнь. Холодовой нейроваскулит Лишний вес. Ожирение
Лишний вес. Ожирение Герпетическая инфекция
Герпетическая инфекция Видеотрансляция операции Лабиринт IIIB
Видеотрансляция операции Лабиринт IIIB