- Эффекторные свойства иммуноглобулинов и гуморальный ответ в разных частях организма

Содержание
- 2. Антитела имеют дело с экстраклеточными (вне хозяйской клетки) формами патогенов и их токсических продуктов. Для доставки
- 3. Эффекторные свойства разных классов иммуноглобулинов определяются их константными областями – способностью связываться с Fc-рецепторами и комплементом.
- 4. IgM – пентамер, связанный дисульфидными связями и J-цепью. 970kDa. Первые АТ в ходе иммунного ответа, не
- 5. Эффекторные свойства разных классов иммуноглобулинов определяются их константными областями – способностью связываться с Fc-рецепторами и комплементом.
- 6. IgG, IgA, IgE: Меньше по размерам, чем IgM, легко проходят из крови в ткани IgG/IgE –
- 7. Для выполнения своей функции – узнавания и взаимодействия с АГ, - молекула АТ должна быть доставлена
- 8. IgA ПОПАДАЕТ НА ПОВЕРХНОСТЬ ЭПИТЕЛИЯ ПУТЕМ ТРАНСЦИТОЗА = dimeric IgA+secretory component of pIgR IgA-защита эпителиальных поверхностей
- 9. Трансцитоз IgA из места синтеза на поверхность слизистого эпителия
- 10. Кроме IgA антител от матери к ребенку попадают IgG антитела (у человека - до рождения, у
- 11. Трансцитоз через плаценту антител класса IgG у человека Ребенок рождается, имея тот же уровень и то
- 12. Распределение изотипов ИГ в тканях и органах человека отличается определенной селективностью. IgG/IgM – кровь IgG/мономерный IgA
- 13. Высокоаффинные IgG и IgA антитела могут нейтрализовать бактериальные токсины Нейтрализующие АТ – АТ, связывающие ту часть
- 14. Высокоаффинные IgG и IgA антитела могут нейтрализовать вирусы и предотвращать инфицирование организма. Вирусы попадают в организм,
- 15. IgG и IgA антитела могут блокировать адгезию бактерий на клетках хозяина. Бактерии имеют адгезины, позволяющие им
- 16. Антиген (патоген) после связывания с антителом во внеклеточном пространстве должен быть удален из организма! Как?
- 17. Способы элиминации антигена с помощью антител
- 18. Комплексы антиген/антитело активируют классический путь комплемента, связываясь с C1q.
- 19. Классический путь активации комплемента начинается с активации комплекса С1. С1q распознает константные области IgM и IgG,
- 20. Как обеспечивается специфичность фиксации комплемента на комплексе АГ-АТ? Почему комплемент не связывается со свободными АТ в
- 21. Как обеспечивается специфичность фиксации комплемента на комплексе АГ-АТ? Почему комплемент не связывается со свободными АТ в
- 22. Антитела – IgM или IgG. Связь с C1q – только когда АТ связаны с АГ на
- 23. IgM и IgG начинают классический путь активации комплемента на поверхности патогена. IgM более эффективен чем IgG
- 24. Классический путь активации комплемента генерирует С3 конвертазу (комплекс С4b,C2a, ковалентно связанный с поверхностью патогена), которая осаждает
- 25. Рецепторы к комплементу необходимы для удаления иммунных комплексов из циркуляции. Иммунные комплексы (ИК) – небольшие комплексы
- 26. АТ обнаруживают патоген и связываются с ним. НО: АТ не могут удалить патоген из организма. Для
- 27. Fc рецепторы – семейство поверхностных молекул, суперсемейства ИГ. Каждый рецептор связывает один или несколько близкородственных изотипов
- 28. Разные Fc рецепторы связывают разные изотипы ИГ и экспрессируются на разных типах клеток. Fc рецепторы –
- 29. Fc рецепторы на фагоцитах активируются антителами, связанными с поверхностью патогена, активируют фагоциты и позволяют им захватить
- 30. В случае больших патогенов (гельминты) фагоцитоз не работает. Макрофаги и нефагоцитирующие клетки (эозинофилы, базофилы, тучные клетки)
- 31. Тучные клетки, базофилы и активированные эозинофилы связывают IgE антитела через высокоаффинный FcεRI рецептор. Весь секретируемый IgE
- 33. Скачать презентацию
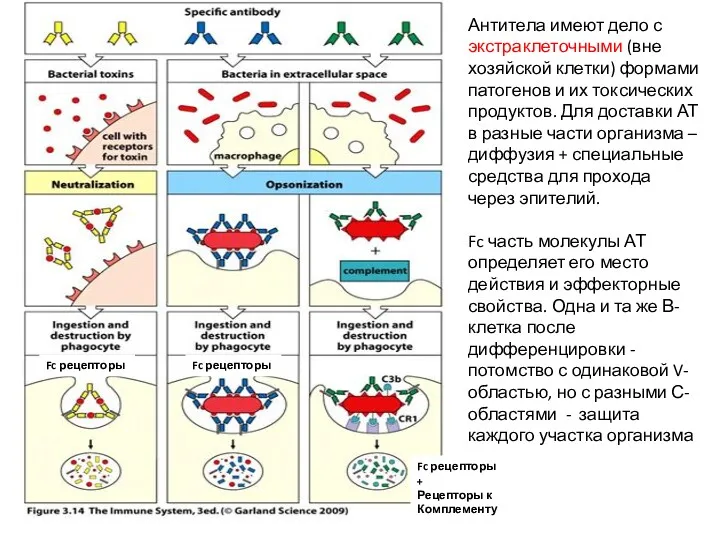
Антитела имеют дело с экстраклеточными (вне хозяйской клетки) формами патогенов и
Антитела имеют дело с экстраклеточными (вне хозяйской клетки) формами патогенов и
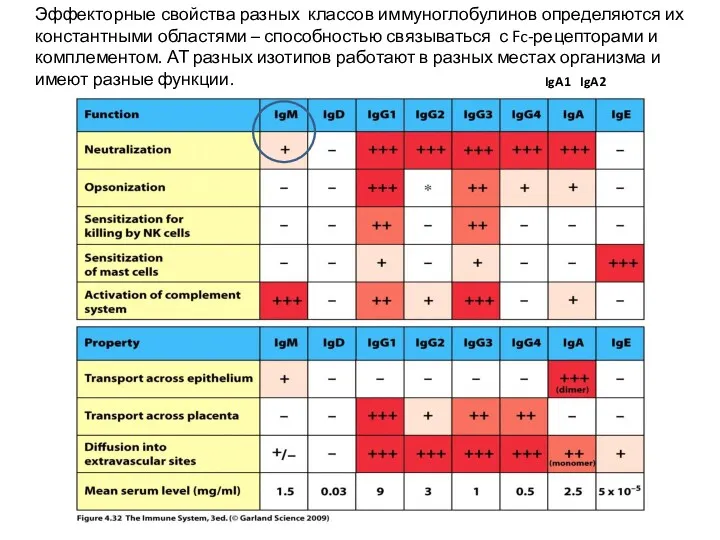
Эффекторные свойства разных классов иммуноглобулинов определяются их константными областями – способностью
Эффекторные свойства разных классов иммуноглобулинов определяются их константными областями – способностью
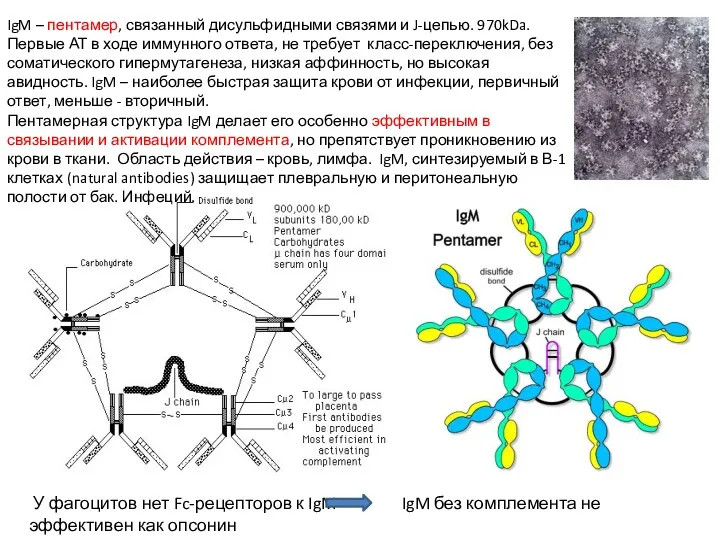
IgM – пентамер, связанный дисульфидными связями и J-цепью. 970kDa. Первые АТ
IgM – пентамер, связанный дисульфидными связями и J-цепью. 970kDa. Первые АТ
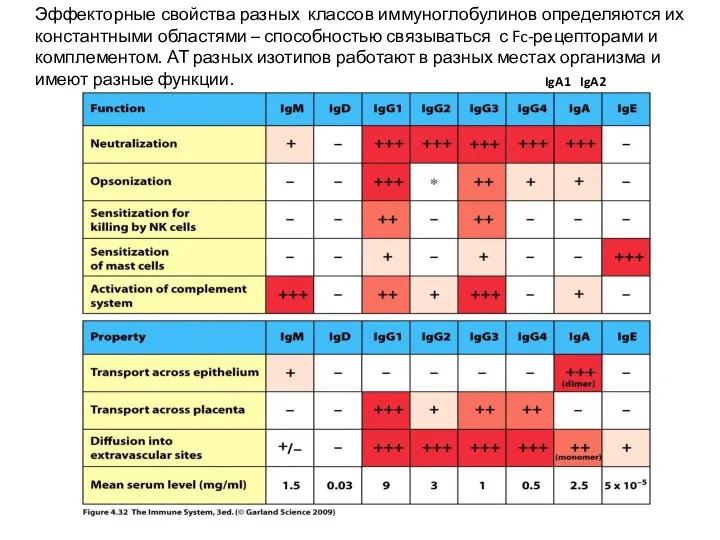
Эффекторные свойства разных классов иммуноглобулинов определяются их константными областями – способностью
Эффекторные свойства разных классов иммуноглобулинов определяются их константными областями – способностью

IgG, IgA, IgE:
Меньше по размерам, чем IgM, легко проходят
IgG, IgA, IgE:
Меньше по размерам, чем IgM, легко проходят
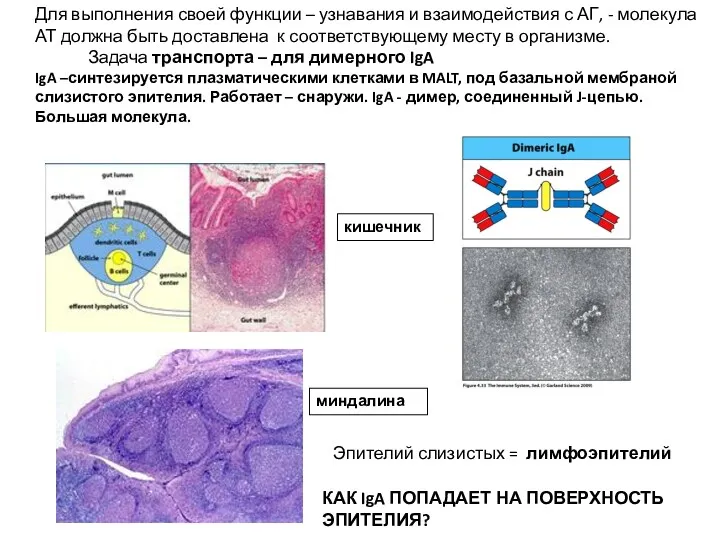
Для выполнения своей функции – узнавания и взаимодействия с АГ, -
Для выполнения своей функции – узнавания и взаимодействия с АГ, -
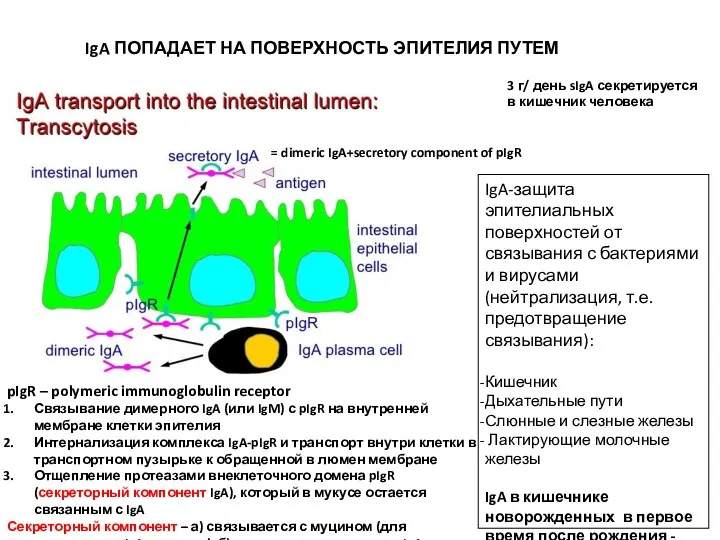
IgA ПОПАДАЕТ НА ПОВЕРХНОСТЬ ЭПИТЕЛИЯ ПУТЕМ ТРАНСЦИТОЗА
= dimeric IgA+secretory component of
IgA ПОПАДАЕТ НА ПОВЕРХНОСТЬ ЭПИТЕЛИЯ ПУТЕМ ТРАНСЦИТОЗА
= dimeric IgA+secretory component of
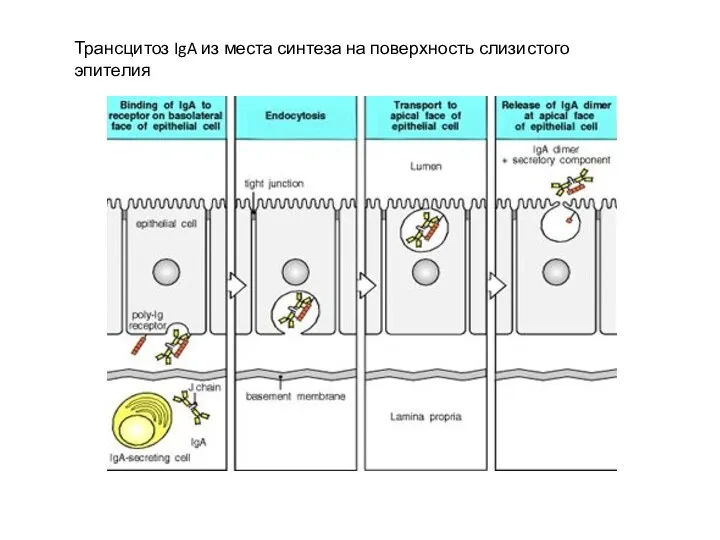
Трансцитоз IgA из места синтеза на поверхность слизистого эпителия
Трансцитоз IgA из места синтеза на поверхность слизистого эпителия
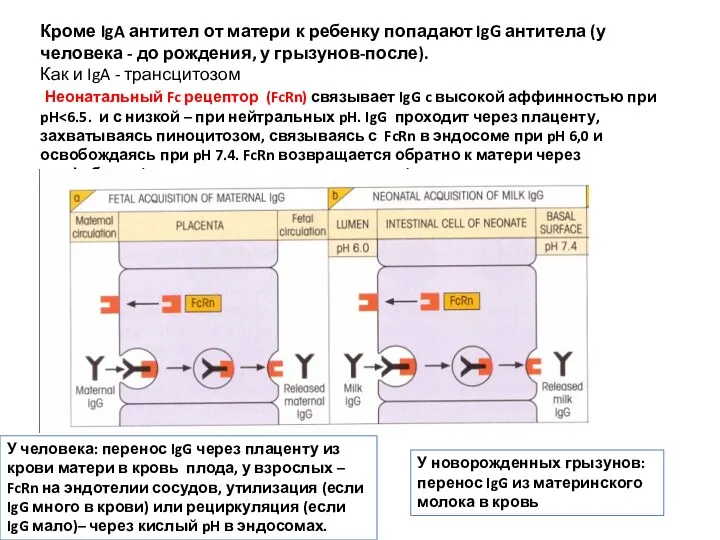
Кроме IgA антител от матери к ребенку попадают IgG антитела (у
Кроме IgA антител от матери к ребенку попадают IgG антитела (у
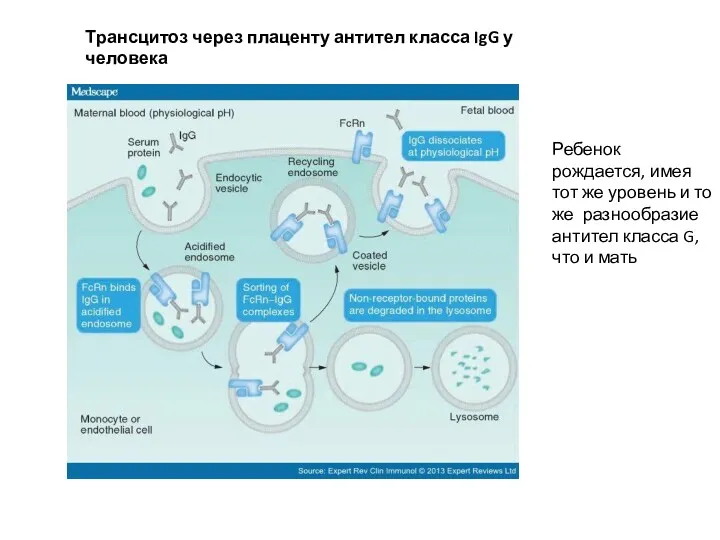
Трансцитоз через плаценту антител класса IgG у человека
Ребенок рождается, имея тот
Трансцитоз через плаценту антител класса IgG у человека
Ребенок рождается, имея тот
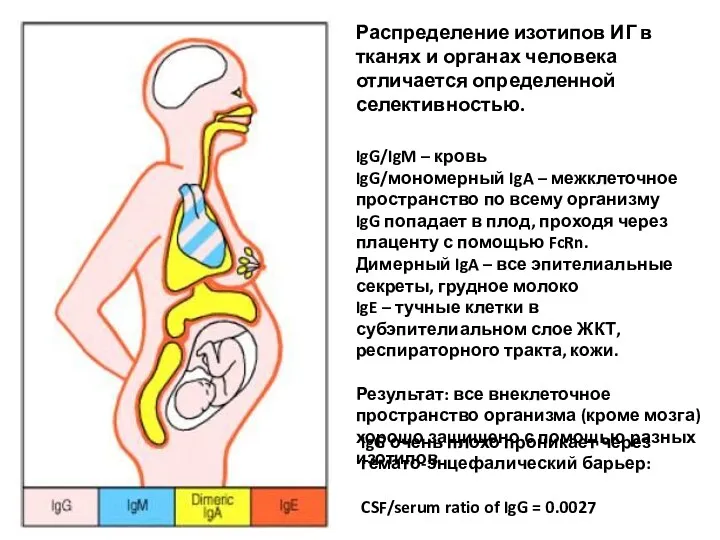
Распределение изотипов ИГ в тканях и органах человека отличается определенной селективностью.
Распределение изотипов ИГ в тканях и органах человека отличается определенной селективностью.
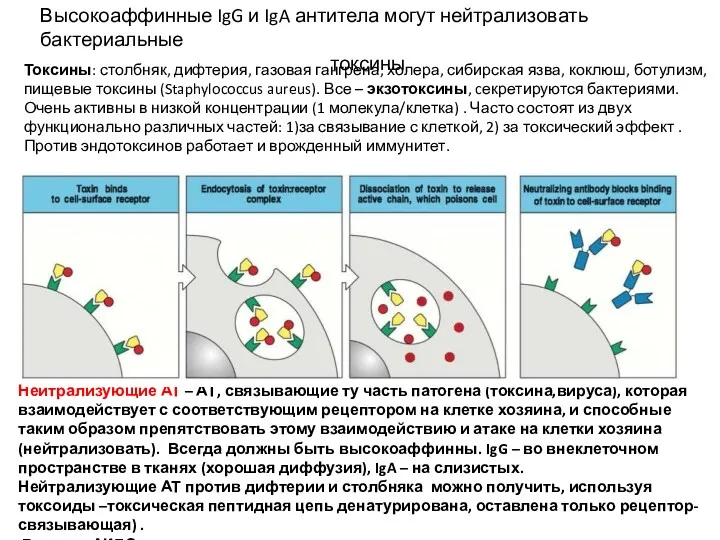
Высокоаффинные IgG и IgA антитела могут нейтрализовать бактериальные
токсины
Нейтрализующие АТ –
Высокоаффинные IgG и IgA антитела могут нейтрализовать бактериальные
токсины
Нейтрализующие АТ –
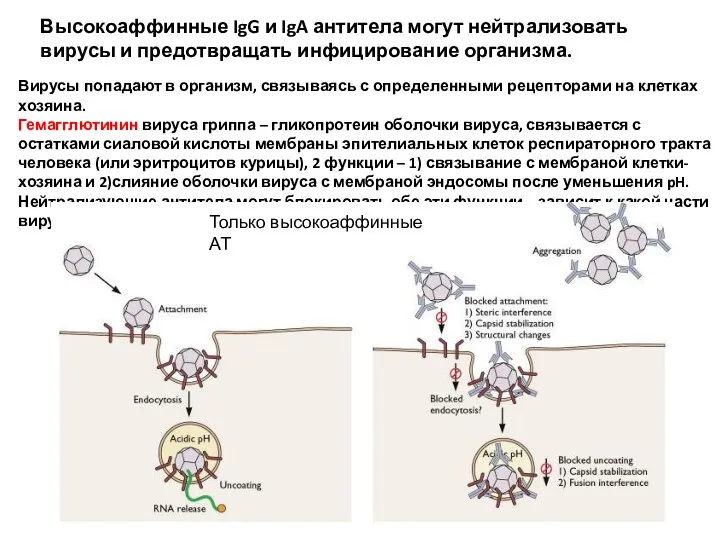
Высокоаффинные IgG и IgA антитела могут нейтрализовать вирусы и предотвращать инфицирование
Высокоаффинные IgG и IgA антитела могут нейтрализовать вирусы и предотвращать инфицирование
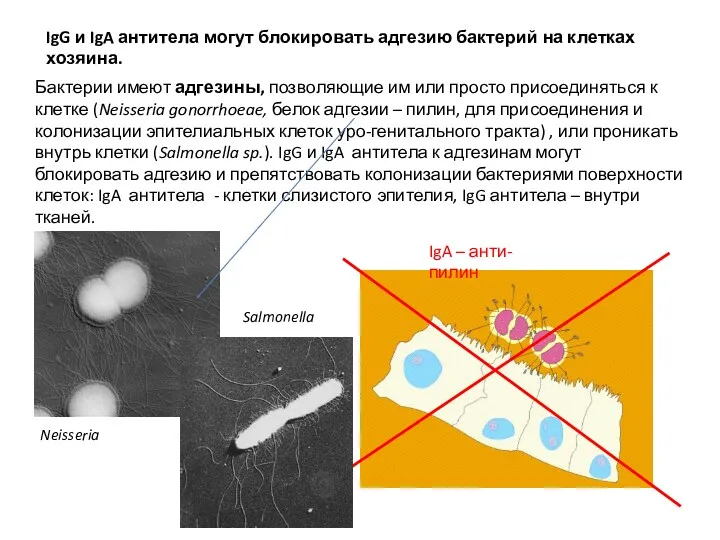
IgG и IgA антитела могут блокировать адгезию бактерий на клетках хозяина.
Бактерии
IgG и IgA антитела могут блокировать адгезию бактерий на клетках хозяина.
Бактерии

Антиген (патоген) после связывания с антителом во внеклеточном пространстве должен быть
Антиген (патоген) после связывания с антителом во внеклеточном пространстве должен быть
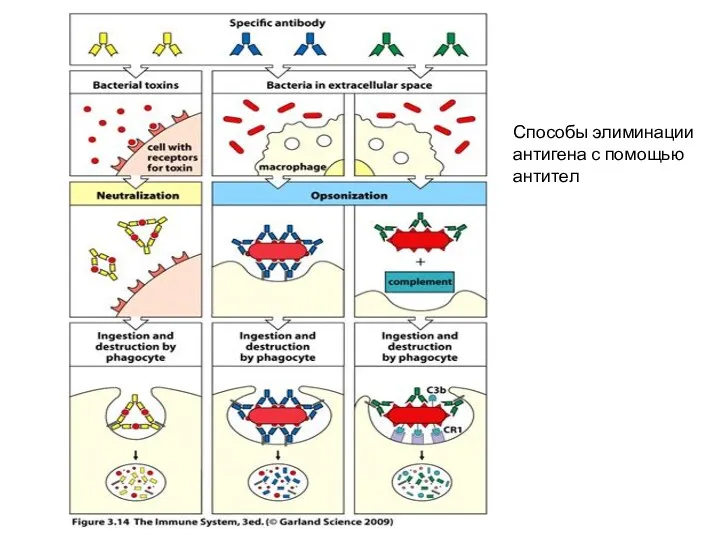
Способы элиминации антигена с помощью антител
Способы элиминации антигена с помощью антител

Комплексы антиген/антитело активируют классический путь комплемента, связываясь с C1q.
Комплексы антиген/антитело активируют классический путь комплемента, связываясь с C1q.
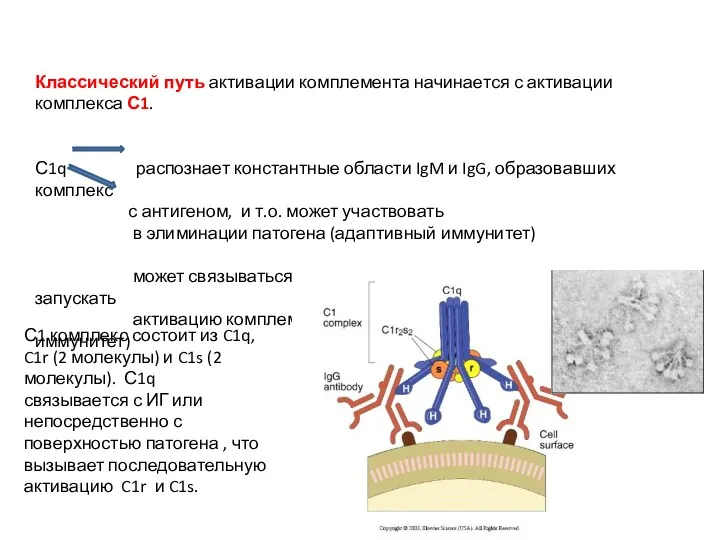
Классический путь активации комплемента начинается с активации комплекса С1.
С1q распознает константные
Классический путь активации комплемента начинается с активации комплекса С1.
С1q распознает константные

Как обеспечивается специфичность фиксации комплемента на комплексе АГ-АТ?
Почему комплемент не связывается
Как обеспечивается специфичность фиксации комплемента на комплексе АГ-АТ?
Почему комплемент не связывается

Как обеспечивается специфичность фиксации комплемента на комплексе АГ-АТ?
Почему комплемент не связывается
Как обеспечивается специфичность фиксации комплемента на комплексе АГ-АТ?
Почему комплемент не связывается
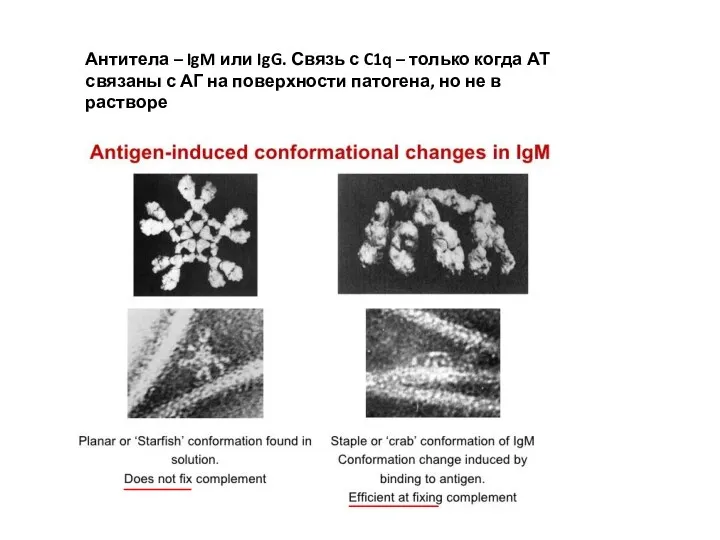
Антитела – IgM или IgG. Связь с C1q – только когда
Антитела – IgM или IgG. Связь с C1q – только когда
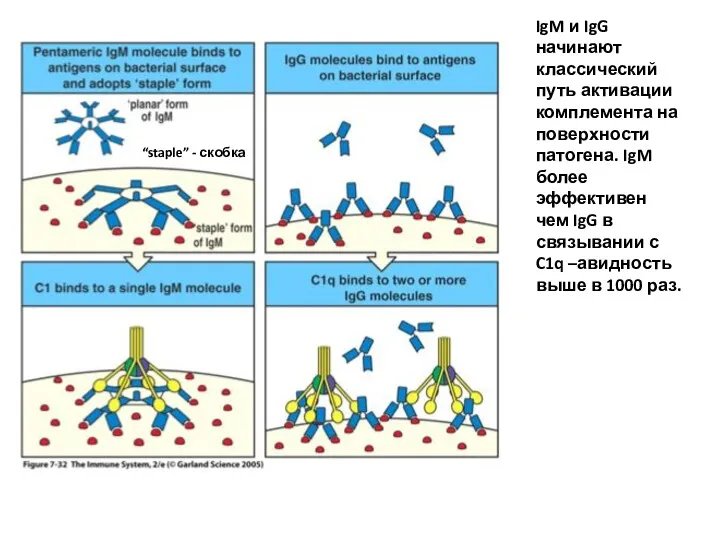
IgM и IgG начинают классический путь активации комплемента на поверхности патогена.
IgM и IgG начинают классический путь активации комплемента на поверхности патогена.
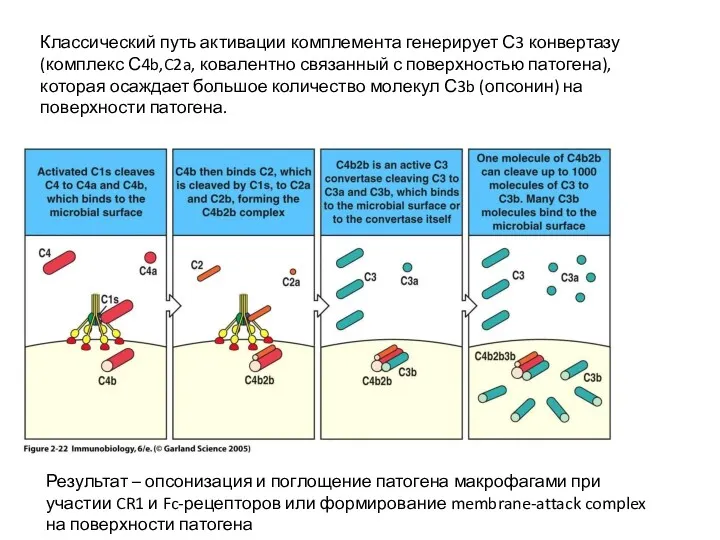
Классический путь активации комплемента генерирует С3 конвертазу (комплекс С4b,C2a, ковалентно связанный
Классический путь активации комплемента генерирует С3 конвертазу (комплекс С4b,C2a, ковалентно связанный
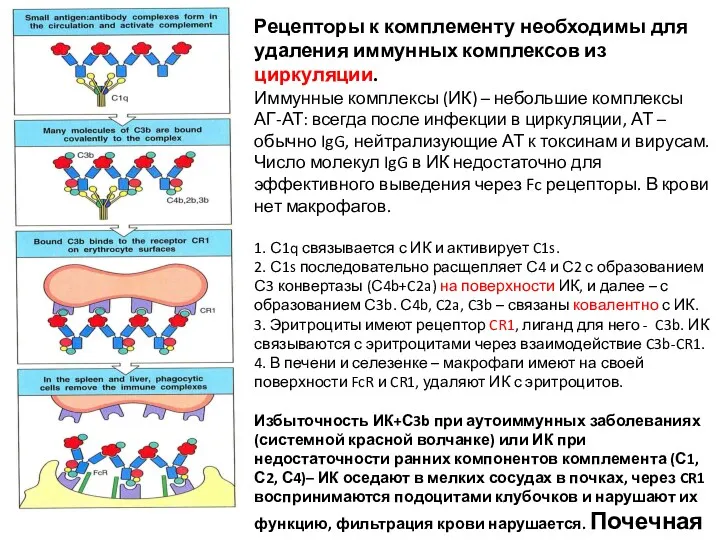
Рецепторы к комплементу необходимы для удаления иммунных комплексов из циркуляции.
Иммунные комплексы
Рецепторы к комплементу необходимы для удаления иммунных комплексов из циркуляции.
Иммунные комплексы
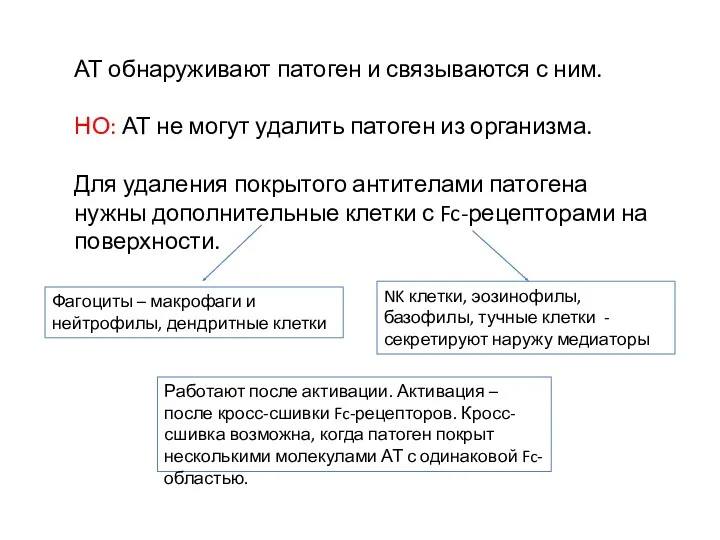
АТ обнаруживают патоген и связываются с ним.
НО: АТ не могут удалить
АТ обнаруживают патоген и связываются с ним.
НО: АТ не могут удалить
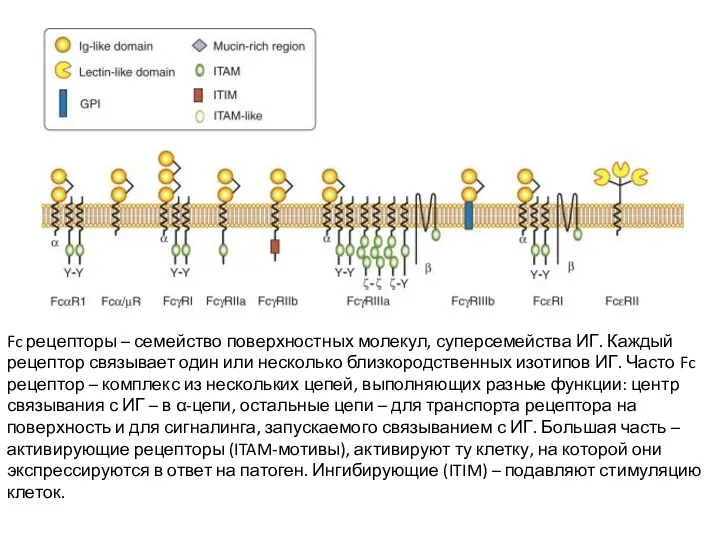
Fc рецепторы – семейство поверхностных молекул, суперсемейства ИГ. Каждый рецептор связывает
Fc рецепторы – семейство поверхностных молекул, суперсемейства ИГ. Каждый рецептор связывает
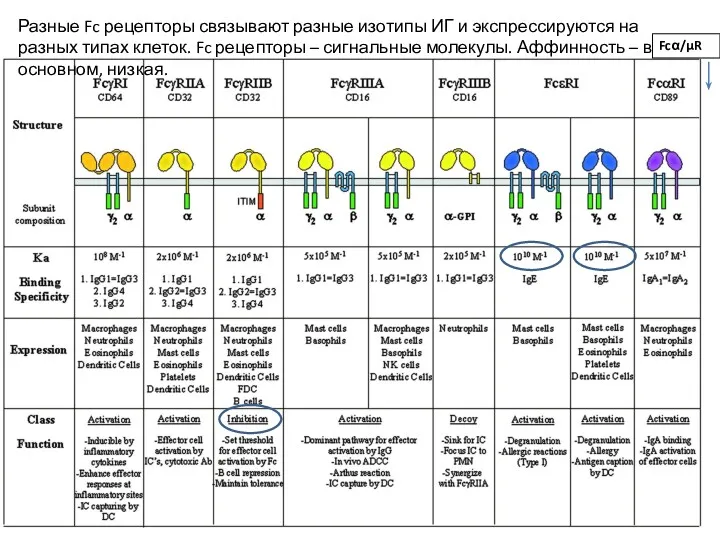
Разные Fc рецепторы связывают разные изотипы ИГ и экспрессируются на разных
Разные Fc рецепторы связывают разные изотипы ИГ и экспрессируются на разных
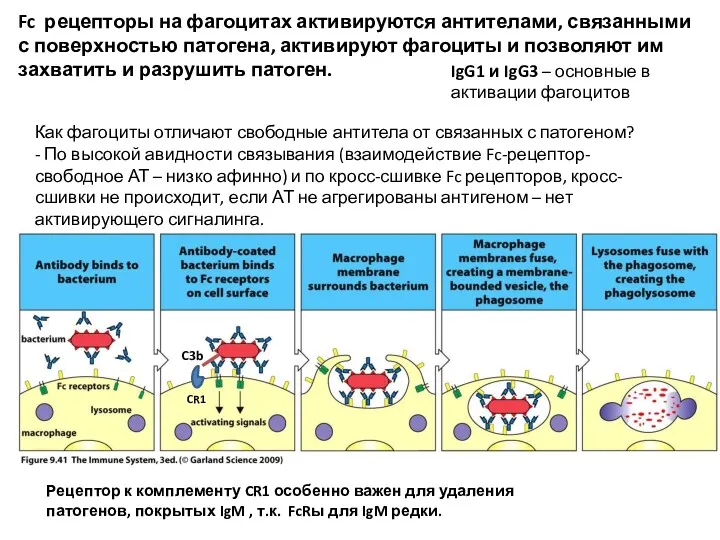
Fc рецепторы на фагоцитах активируются антителами, связанными с поверхностью патогена, активируют
Fc рецепторы на фагоцитах активируются антителами, связанными с поверхностью патогена, активируют
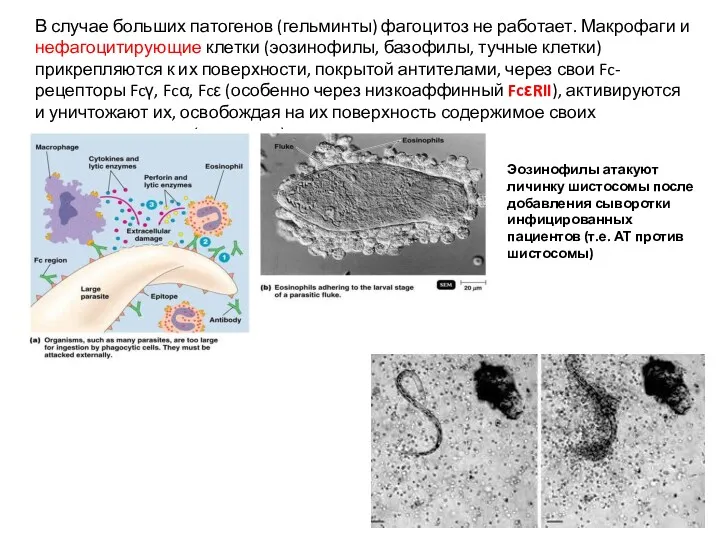
В случае больших патогенов (гельминты) фагоцитоз не работает. Макрофаги и нефагоцитирующие
В случае больших патогенов (гельминты) фагоцитоз не работает. Макрофаги и нефагоцитирующие
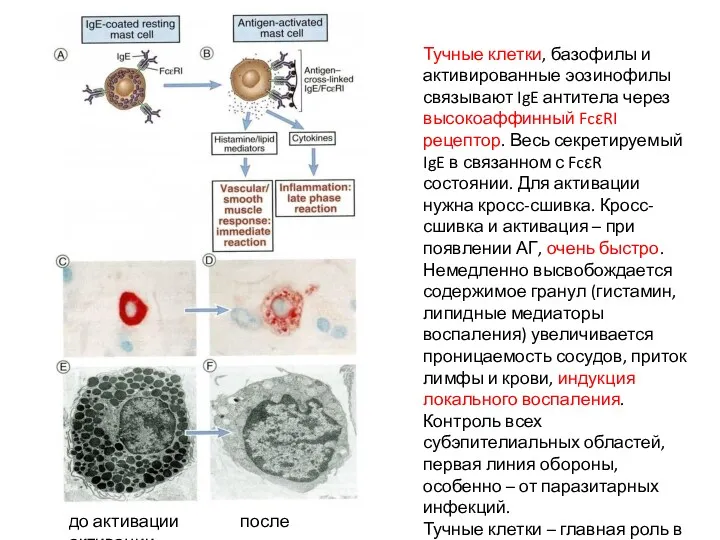
Тучные клетки, базофилы и активированные эозинофилы связывают IgE антитела через высокоаффинный
Тучные клетки, базофилы и активированные эозинофилы связывают IgE антитела через высокоаффинный
 Физиология и патология периода новорожденности
Физиология и патология периода новорожденности Технология изготовления полных съёмных пластиночных протезов при полном отсутствии зубов с использованием современных материалов
Технология изготовления полных съёмных пластиночных протезов при полном отсутствии зубов с использованием современных материалов Экстракорпоральное оплодотворение
Экстракорпоральное оплодотворение ХСН: причины, классификация, консервативное лечение
ХСН: причины, классификация, консервативное лечение Қан плазмасының азотты органикалық құрам бөліктері және оларды анықтаудың клиникалық маңызы
Қан плазмасының азотты органикалық құрам бөліктері және оларды анықтаудың клиникалық маңызы Заболевания плевры. Плевриты
Заболевания плевры. Плевриты Особенности препарирования кариозных полостей по Блэку
Особенности препарирования кариозных полостей по Блэку Гражданско-правовая ответственность медицинских работников
Гражданско-правовая ответственность медицинских работников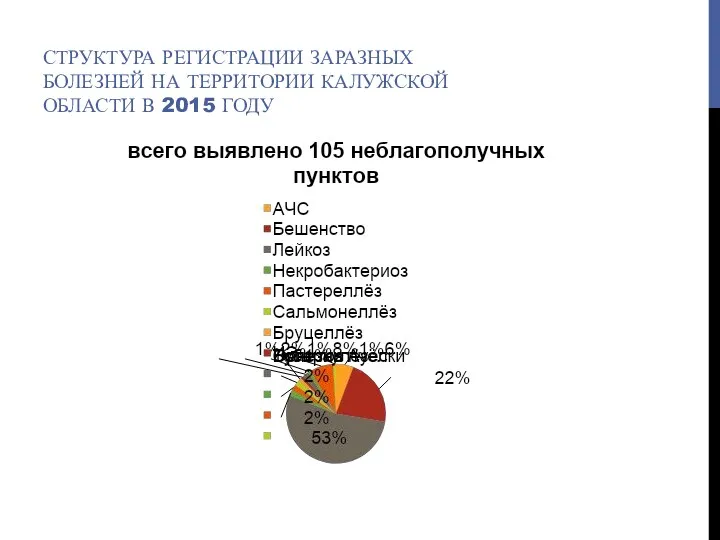 Структура регистрации заразных болезней на территории Калужской области в 2015 году
Структура регистрации заразных болезней на территории Калужской области в 2015 году Мүгедектік деген не
Мүгедектік деген не Инфекционный контроль
Инфекционный контроль Маточные кровотечения как ургентное состояние. Дифференциальная диагностика
Маточные кровотечения как ургентное состояние. Дифференциальная диагностика Грипп и ОРВИ
Грипп и ОРВИ Аллергия или гиперчувствительность
Аллергия или гиперчувствительность Рассеянный склероз
Рассеянный склероз Дети с нарушениями функций опорно-двигательного аппарата
Дети с нарушениями функций опорно-двигательного аппарата План исследования больного при заболеваниях системы дыхания
План исследования больного при заболеваниях системы дыхания Запальні захворювання жіночих статевих органів
Запальні захворювання жіночих статевих органів Тұрақты электротоктардың медицинада қолданылуы
Тұрақты электротоктардың медицинада қолданылуы Дезагрегантная терапия острого коронарного синдрома на реанимационном этапе
Дезагрегантная терапия острого коронарного синдрома на реанимационном этапе Острые отравления хлорпирофосом
Острые отравления хлорпирофосом Онтогенез. Определение понятия
Онтогенез. Определение понятия Психогения - кратковременная реакция или длительное состояние (болезнь), возникающие вследствие психотравмы
Психогения - кратковременная реакция или длительное состояние (болезнь), возникающие вследствие психотравмы Стоматология. Диагностика
Стоматология. Диагностика Диагностика заболеваний эндокринной системы. Диагностика сахарного диабета. Диагностика диффузно-токсического зоба
Диагностика заболеваний эндокринной системы. Диагностика сахарного диабета. Диагностика диффузно-токсического зоба Различные методики нейровизуализации в неврологии
Различные методики нейровизуализации в неврологии Психотропные средства. Нейролептики. Транквилизаторы. Седативные средства
Психотропные средства. Нейролептики. Транквилизаторы. Седативные средства Клинический разбор
Клинический разбор